
Estimating the Population Size of Masked Palm Civets Using Hair-Snaring in Southwest China

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Hair Sample Collection
2.3. Species Identification
2.4. Individual Identification
2.5. Population Density Estimation
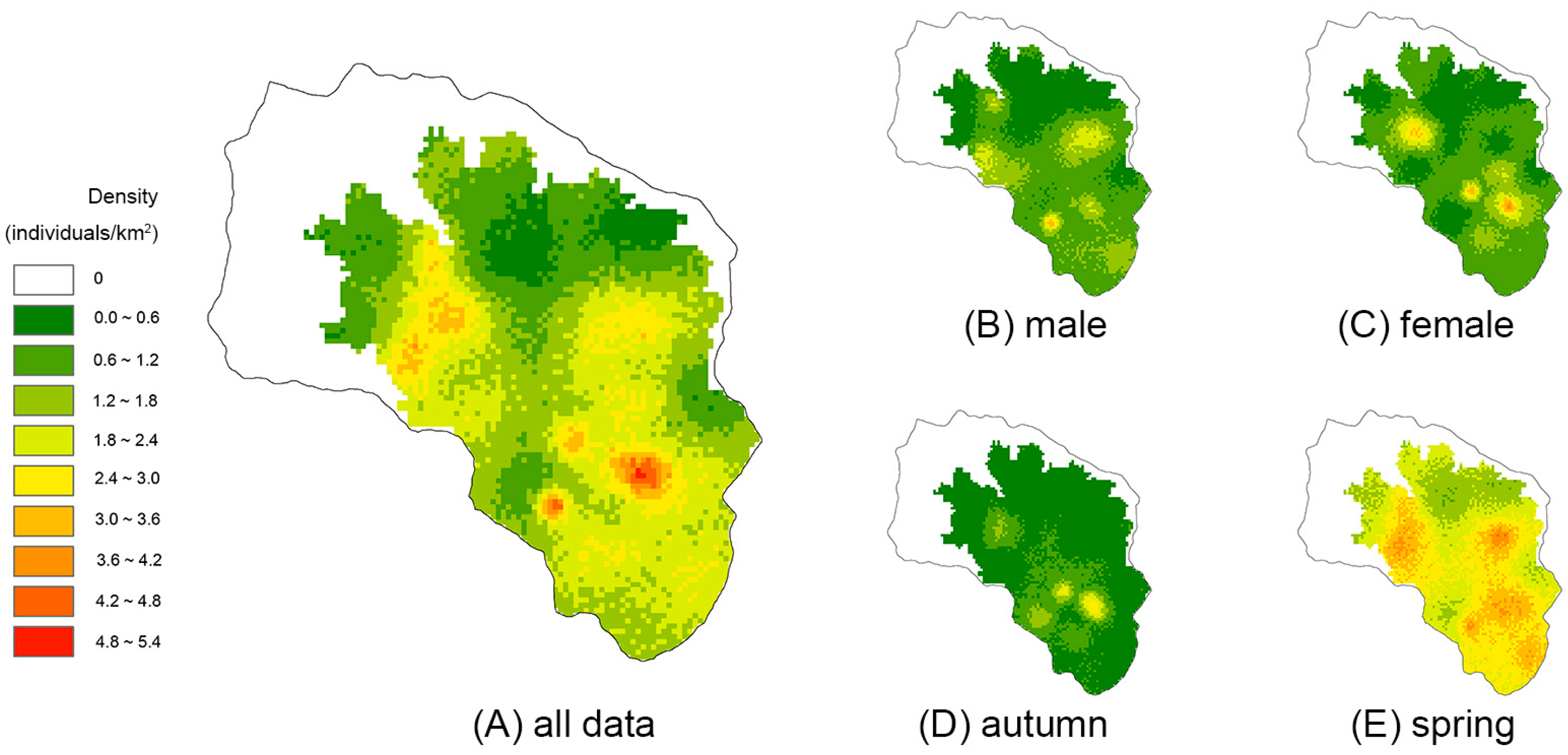
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ritchie, E.G.; Johnson, C.N. Predator interactions, mesopredator release and biodiversity conservation. Ecol. Lett. 2009, 12, 982–998. [Google Scholar] [CrossRef] [PubMed]
- Roemer, G.W.; Gompper, M.E.; van Valkenburgh, B. The ecological role of the mammalian mesocarnivore. BioScience 2009, 59, 165–173. [Google Scholar] [CrossRef]
- Crooks, K.R.; Soule, M.E. Mesopredator release and avifaunal extinctions in a fragmented system. Nature 1999, 400, 563–566. [Google Scholar] [CrossRef]
- Prugh, L.R.; Stoner, C.J.; Epps, C.W.; Bean, W.T.; Ripple, W.J.; Laliberte, A.S.; Brashares, J.S. The rise of the mesopredator. BioSience 2009, 59, 779–791. [Google Scholar] [CrossRef]
- Lesmeister, D.B.; Nielsen, C.K.; Schauber, E.M.; Hellgren, E.C. Spatial and temporal structure of a mesocarnivore guild in midwestern North America. Wildl. Monogr. 2015, 191, 1–61. [Google Scholar] [CrossRef]
- de Satgé, J.; Teichman, K.; Cristescu, B. Competition and coexistence in a small carnivore guild. Oecologia 2017, 184, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, D.Q.; Hacker, C.; Dong, W.; Wu, B.; Xue, Y.D. Spatial co-occurrence and temporal activity patterns of sympatric mesocarnivores guild in Qinling Mountains. Glob. Ecol. Conserv. 2022, 36, 02129. [Google Scholar] [CrossRef]
- Avrin, A.C.; Pekins, C.E.; Wilmers, C.C.; Sperry, J.H.; Allen, M.L. Can a mesocarnivore fill the functional role of an apex predator? Ecosphere 2023, 14, e4383. [Google Scholar] [CrossRef]
- Jung, T.S.; Boonstra, R.; Krebs, C.J. Mark my words: Experts’ choice of marking methods used in capture-mark-recapture studies of small mammals. J. Mammal. 2020, 101, 307–317. [Google Scholar] [CrossRef]
- Schwartz, M.K.; Luikart, G.; Waples, R.S. Genetic monitoring as a promising tool for conservation and management. Trends Ecol. Evol. 2007, 22, 25–33. [Google Scholar] [CrossRef]
- Rowcliffe, J.M.; Kays, R.; Kranstauber, B.; Carbone, C.; Jansen, P.A. Quantifying levels of animal activity using camera trap data. Methods Ecol. Evol. 2014, 5, 1170–1179. [Google Scholar] [CrossRef]
- Liu, S.; Wu, Y.; Li, S. Handbook of the Mammals of China, 3rd ed.; The Straits Publishing and Distributing Group: Fuzhou, China, 2022. [Google Scholar]
- Zhang, L.; Chang, J.; Feng, C.; Wan, D. New records of Paguma larvata in Liaoning Province, China. Acta Theriol. Sin. 2023, 43, 760–764. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, J.; Slade, E.; Zhang, L.; Palomares, F.; Chen, J.; Wang, X.; Zhang, S. Dietary shifts in relation to fruit availability among masked palm civets (Paguma larvata) in Central China. J. Mammal. 2008, 89, 435–447. [Google Scholar] [CrossRef]
- Corlett, R.T. Seed dispersal in Hong Kong, China: Past, present and possible futures. Integr. Zool. 2011, 6, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Bu, H.; Wang, F.; McShea, W.J.; Lu, Z.; Wang, D.; Li, S. Spatial co-occurrence and activity patterns of mesocarnivores in the temperate forests of Southwest China. PLoS ONE 2016, 11, e0164271. [Google Scholar] [CrossRef] [PubMed]
- Bu, H.; Hopkins, I.I.I.J.B.; Li, S.; Wang, D. Seasonal distribution and activity patterns of mesopredators and their prey in Southwest China. J. Mammal. 2023, 104, 941–950. [Google Scholar] [CrossRef]
- McShea, W.J.; Li, S.; Shen, X.; Wang, F.; Wang, D. Guide to the Wildlife of Southwest China; Smithsonian Institution Scholarly Press: Washington, DC, USA, 2018. [Google Scholar]
- Bu, H.; Hopkins, I.I.I.J.B.; Zhang, D.; Li, S.; Wang, R.; Yao, M.; Wang, D. An evaluation of hair-snaring devices for small-bodied carnivores in Southwest China. J. Mammal. 2016, 97, 589–598. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Tilson, R.; Hu, D.; Muntifering, J.; Nyhus, P.J. Dramatic decline of wild South China tigers Panthera tigris amoyensis: Field survey of priority tiger reserves. Oryx 2004, 38, 40–47. [Google Scholar] [CrossRef]
- Laguardia, A.; Kamler, J.F.; Li, S.; Zhang, C.; Zhou, Z.; Shi, K. The current distribution and status of leopards Panthera pardus in China. Oryx 2017, 51, 153–159. [Google Scholar] [CrossRef]
- Li, S.; McShea, W.J.; Wang, D.; Gu, X.; Zhang, X.; Zhang, L.; Shen, X. Retreat of large carnivores across the giant panda distribution range. Nat. Ecol. Evol. 2020, 4, 1327–1331. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; McShea, W.J.; Wang, D.; Shen, X.; Bu, H.; Guan, T.; Wang, F.; Gu, X.; Zhang, X.; Liao, H. Construction progress of the camera-trapping network for the mountains of Southwest China. Biodivers. Sci. 2020, 28, 1049–1058. [Google Scholar] [CrossRef]
- Li, S.; McShea, W.J.; Wang, D.; Lu, Z.; Gu, X. Gauging the impact of management expertise on the distribution of large mammals across protected areas. Divers. Distrib. 2012, 18, 1166–1176. [Google Scholar] [CrossRef]
- Li, S.; Wang, D.; Bu, H.; Liu, X.; Jin, T. Camera-trapping survey on the mammal diversity of the Laohegou Nature Reserve, Sichuan Province. Acta Theriol. Sin. 2016, 36, 282–291. [Google Scholar] [CrossRef]
- Li, S.; Wang, D.; Chen, X.; Bu, H.; Liu, X.; Jin, T. The wildlife camera-trapping dataset of Laohegou Protected Area, Sichuan Province (2011–2015). Biodivers. Sci. 2021, 29, 1170–1174. [Google Scholar] [CrossRef]
- Rabinowitz, A.R. Behaviour and movements of sympatric civet species in Huai Kha Khaeng Wildlife Sanctuary, Thailand. J. Zool. 1991, 223, 281–298. [Google Scholar] [CrossRef]
- Wang, H.; Fuller, T.K. Notes on the ecology of sympatric small carnivores in southeastern China. Mamm. Biol. 2001, 66, 251–255. [Google Scholar]
- Grassman, L.I., Jr.; Tewes, M.E.; Silvy, N.J.; Kreetiyutanont, K. Ecology of three sympatric felids in a mixed evergreen forest in north-central Thailand. J. Mammal. 2005, 86, 29–38. [Google Scholar] [CrossRef]
- Zhou, Y.; Newman, C.; Palomares, F.; Zhang, S.Y.; Xie, Z.Q.; Macdonald, D.W. Spatial organization and activity patterns of the masked palm civet (Paguma larvata) in central-south China. J. Mammal. 2014, 95, 534–542. [Google Scholar] [CrossRef]
- Xiong, M.; Shao, X.; Long, Y.; Bu, H.; Zhang, D.; Wang, D.; Li, S.; Wang, R.; Yao, M. Molecular analysis of vertebrates and plants in scats of leopard cats (Prionailurus bengalensis) in Southwest China. J. Mammal. 2016, 97, 1054–1064. [Google Scholar] [CrossRef]
- Zhang, D.; Xu, L.; Bu, H.; Wang, D.; Xu, C.; Wang, R. The complete mitochondrial genome of the masked palm civet (Paguma larvata, Mammalia, Carnivora). Mitochondrial DNA Part A 2016, 27, 3764–3765. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.P.; Andersen, D.H.; Veron, G.; Randi, E.; Zhang, S.Y. Isolation and characterization of polymorphic microsatellite markers for the masked palm civet (Paguma larvata). Biochem. Genet. 2008, 46, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Kaneko, Y.; Yamazaki, K.; Anezaki, T.; Yachimori, S.; Ochiai, K.; Lin, L.K.; Pei, K.J.C.; Chen, Y.J.; Chang, S.W.; et al. Genetic population structure of the masked palm civet Paguma larvata (Carnivora: Viverridae) in Japan, revealed from analysis of newly identified compound microsatellites. Conserv. Genet. 2012, 13, 1095–1107. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, D.; Xiong, M.; Bu, H.; Wang, D.; Yao, M.; Li, S.; Wang, R. Development and characterization of polymorphic microsatellite markers in the masked palm civet (Paguma larvata). Acta Sci. Nat. Univ. Pekin. 2021, 57, 395–400. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Xiong, M.; Bu, H.; Wang, D.; Li, S.; Yao, M.; Wang, R. Sex identification of the masked palm civet (Paguma larvata) using noninvasive hair samples. Conserv. Genet. Resour. 2016, 8, 207–209. [Google Scholar] [CrossRef]
- Gopalaswamy, A.M.; Royle, J.A.; Hines, J.E.; Singh, P.; Jathanna, D.; Kumar, N.S.; Karanth, K.U. Program SPACECAP: Software for estimating animal density using spatially explicit capture-recapture models. Methods Ecol. Evol. 2012, 3, 1067–1072. [Google Scholar] [CrossRef]
- Zhang, B.L.; Su, X.L.; Gao, G.C.; Zhang, W.H. Study on distribution and population composition of the masked palm civet (Paguma larvata). Chin. J. Zool. 1991, 26, 42–45. [Google Scholar]
- Jia, Z.; Jiang, Z.; Wang, Z. Copulatory behavior in captive masked palm civets, Paguma larvata. Folia Zool. 2001, 50, 271–279. [Google Scholar]
- Kang, M.; Li, W.; Qu, X.; Zhang, S. Study of the hibernation habit and characteristics of Paguma larvata. J. Hunan Agric. Univ. 1997, 23, 366–369. [Google Scholar]


{kind=link}
{kind=link}
| Parameter | Posterior Mean | Posterior SD | 95% Lower HPD Level | 95% Upper HPD Level |
|---|---|---|---|---|
| σ | 574.4 | 58.0 | 464.7 | 687.5 |
| λ0 | 0.119 | 0.030 | 0.064 | 0.178 |
| ψ | 0.080 | 0.015 | 0.051 | 0.109 |
| Nsuper (individuals) | 82 | 13 | 57 | 107 |
| D (individuals/km2) | 1.650 | 0.266 | 1.144 | 2.147 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, D.; Zhang, D.; Bu, H.; Hopkins, J.B., III; Xiong, M.; Wang, D.; Yao, M.; Li, S.; Wang, R. Estimating the Population Size of Masked Palm Civets Using Hair-Snaring in Southwest China. Diversity 2024, 16, 421. https://doi.org/10.3390/d16070421
Wang D, Zhang D, Bu H, Hopkins JB III, Xiong M, Wang D, Yao M, Li S, Wang R. Estimating the Population Size of Masked Palm Civets Using Hair-Snaring in Southwest China. Diversity. 2024; 16(7):421. https://doi.org/10.3390/d16070421
Chicago/Turabian StyleWang, Di, Dan Zhang, Hongliang Bu, John B. Hopkins, III, Mengyin Xiong, Dajun Wang, Meng Yao, Sheng Li, and Rongjiang Wang. 2024. "Estimating the Population Size of Masked Palm Civets Using Hair-Snaring in Southwest China" Diversity 16, no. 7: 421. https://doi.org/10.3390/d16070421
APA StyleWang, D., Zhang, D., Bu, H., Hopkins, J. B., III, Xiong, M., Wang, D., Yao, M., Li, S., & Wang, R. (2024). Estimating the Population Size of Masked Palm Civets Using Hair-Snaring in Southwest China. Diversity, 16(7), 421. https://doi.org/10.3390/d16070421