
How Does Sharing Data from Research Institutions on Global Biodiversity Information Facility Enhance Its Scientific Value?



Abstract

1. Introduction
- How frequently are the NHC data from FBAMU accessed by users of the AMUNATCOLL and GBIF platforms?
- How much data do users download from both sources?
- How often are these data cited in scientific publications?
- In which journals do publications citing FBAMU appear?
- What is the thematic scope of publications utilizing this database?
- What contributions do publications based on this type of data make to science and nature conservation?
2. Material and Methods
2.1. Natural Collections of the Faculty of Biology at Adam Mickiewicz University in the AMUNATCOLL IT System and on the GBIF Platform
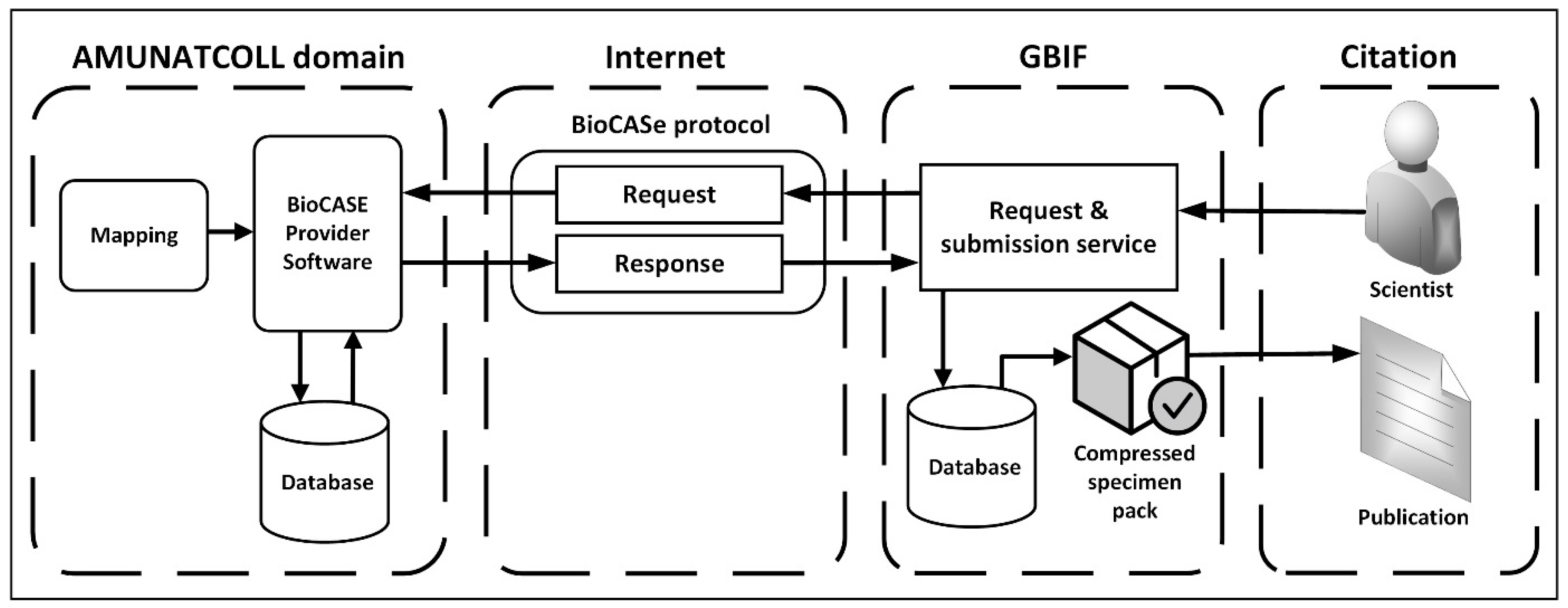
2.2. Data Transfer from AMUNATCOLL IT to GBIF
2.3. Data Sources, Scope, and Analysis Methods
3. Results
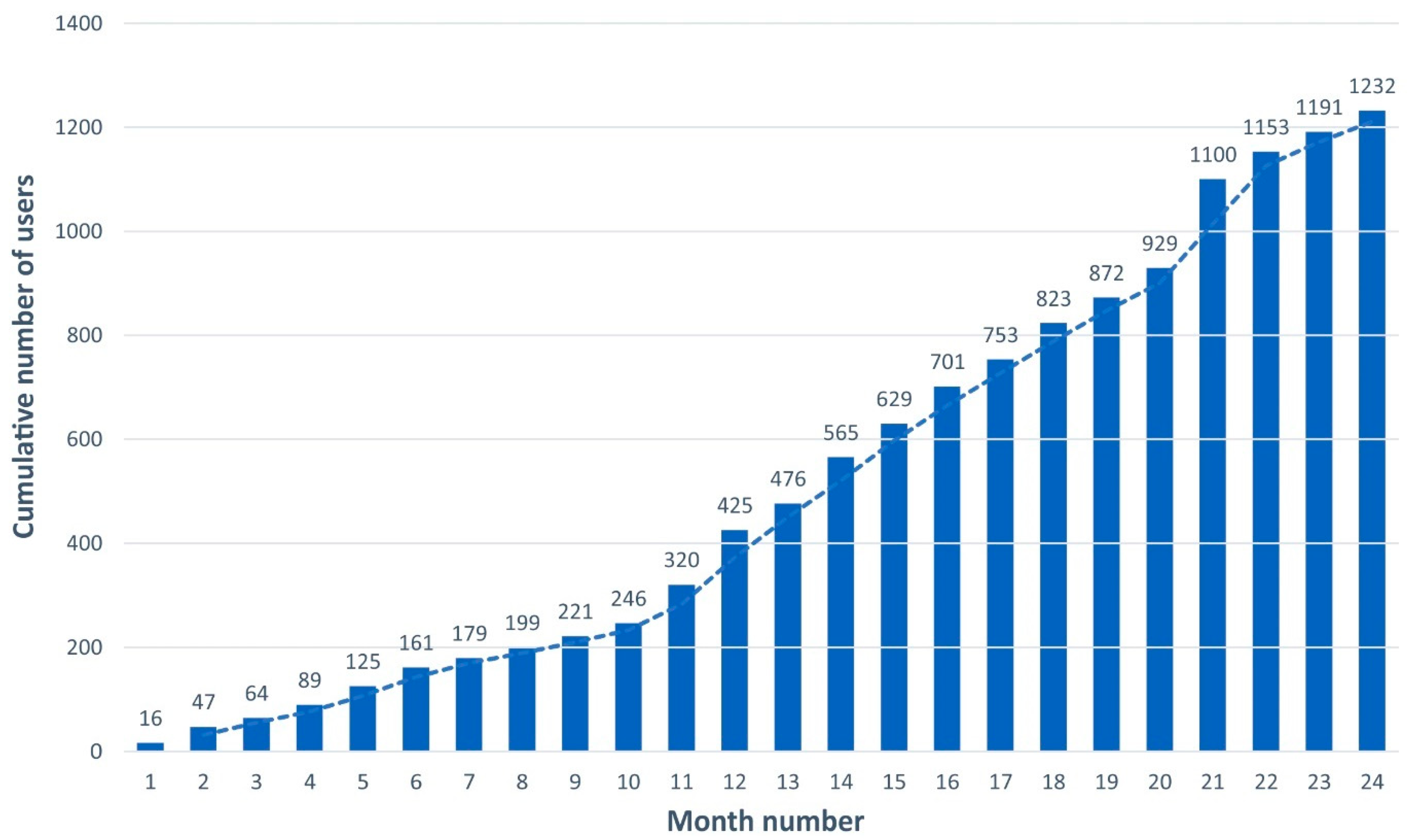
3.1. Use of FBAMU Data Made Available on the AMUNATCOLL Platform
3.2. Use of FBAMU Data Made Available on the GBIF Platform
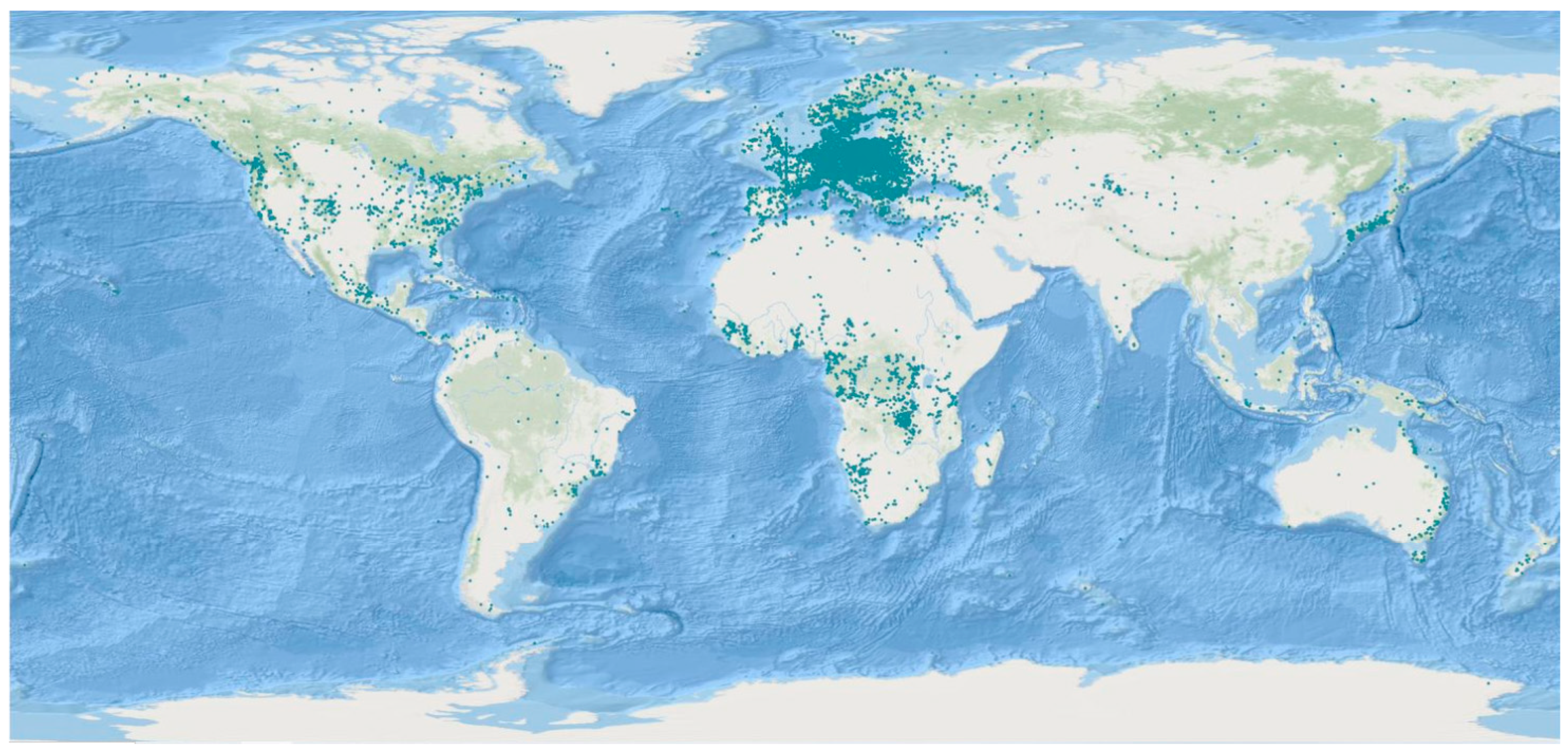
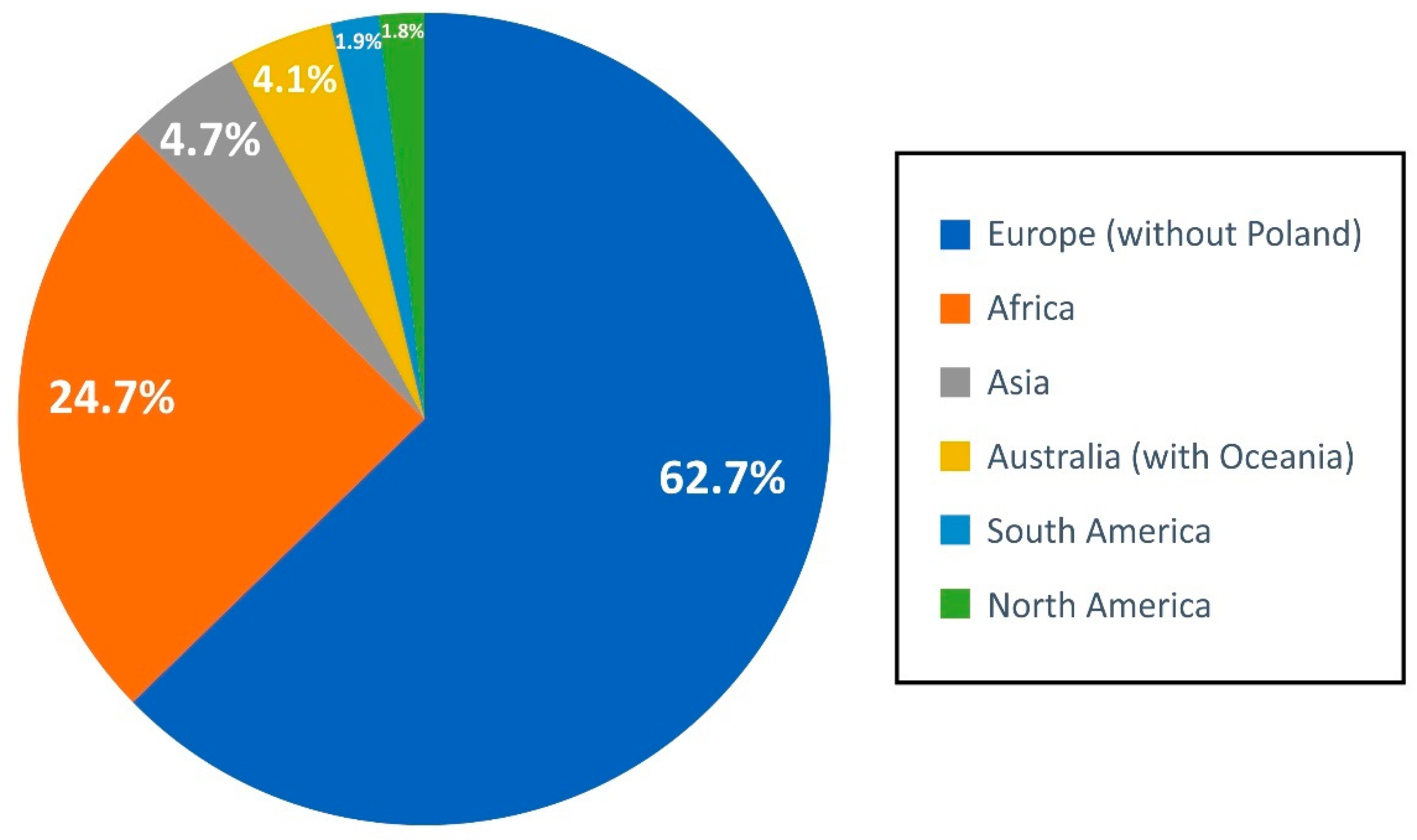
3.2.1. FBAMU Biodiversity Data in GBIF
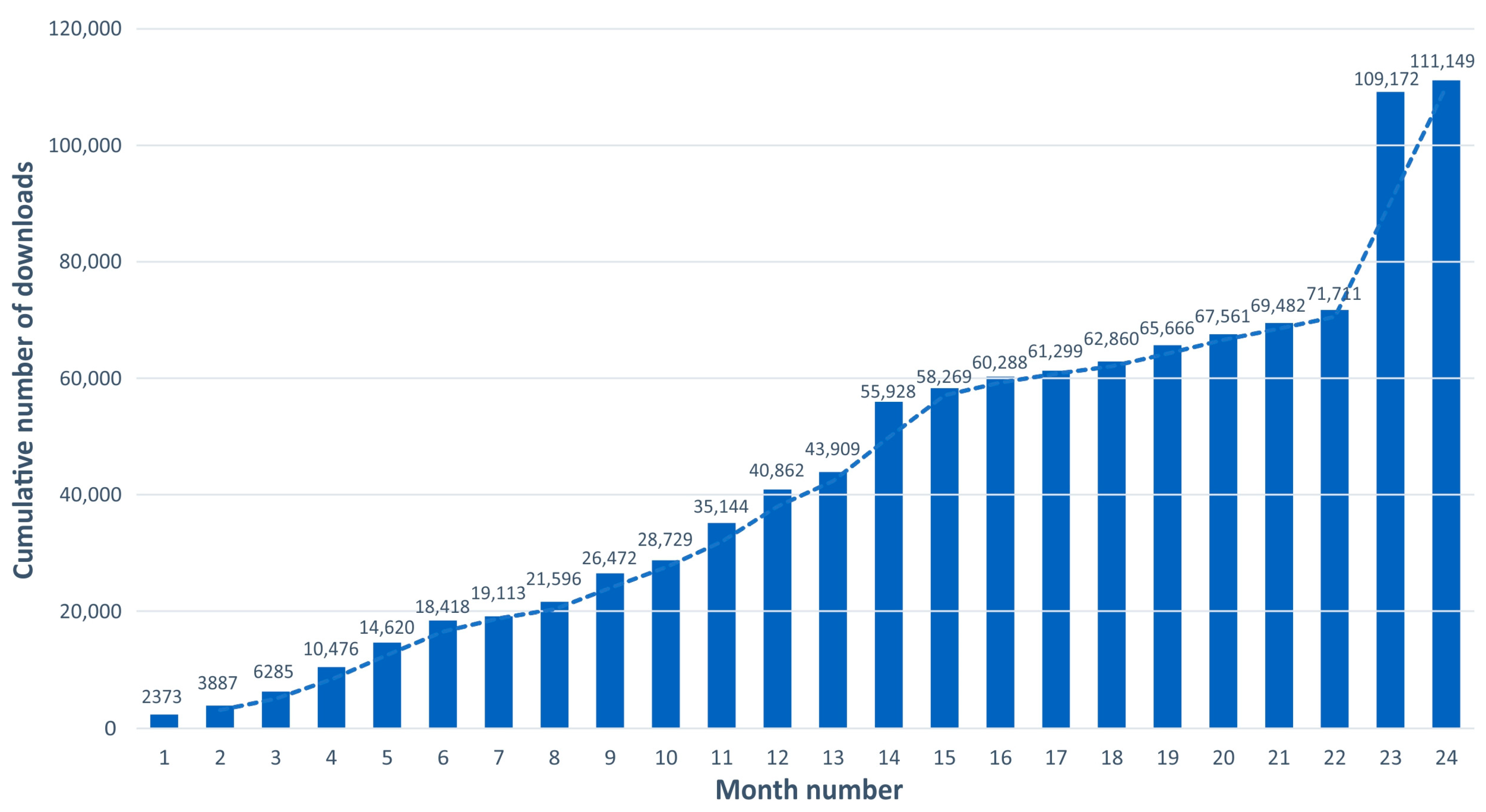
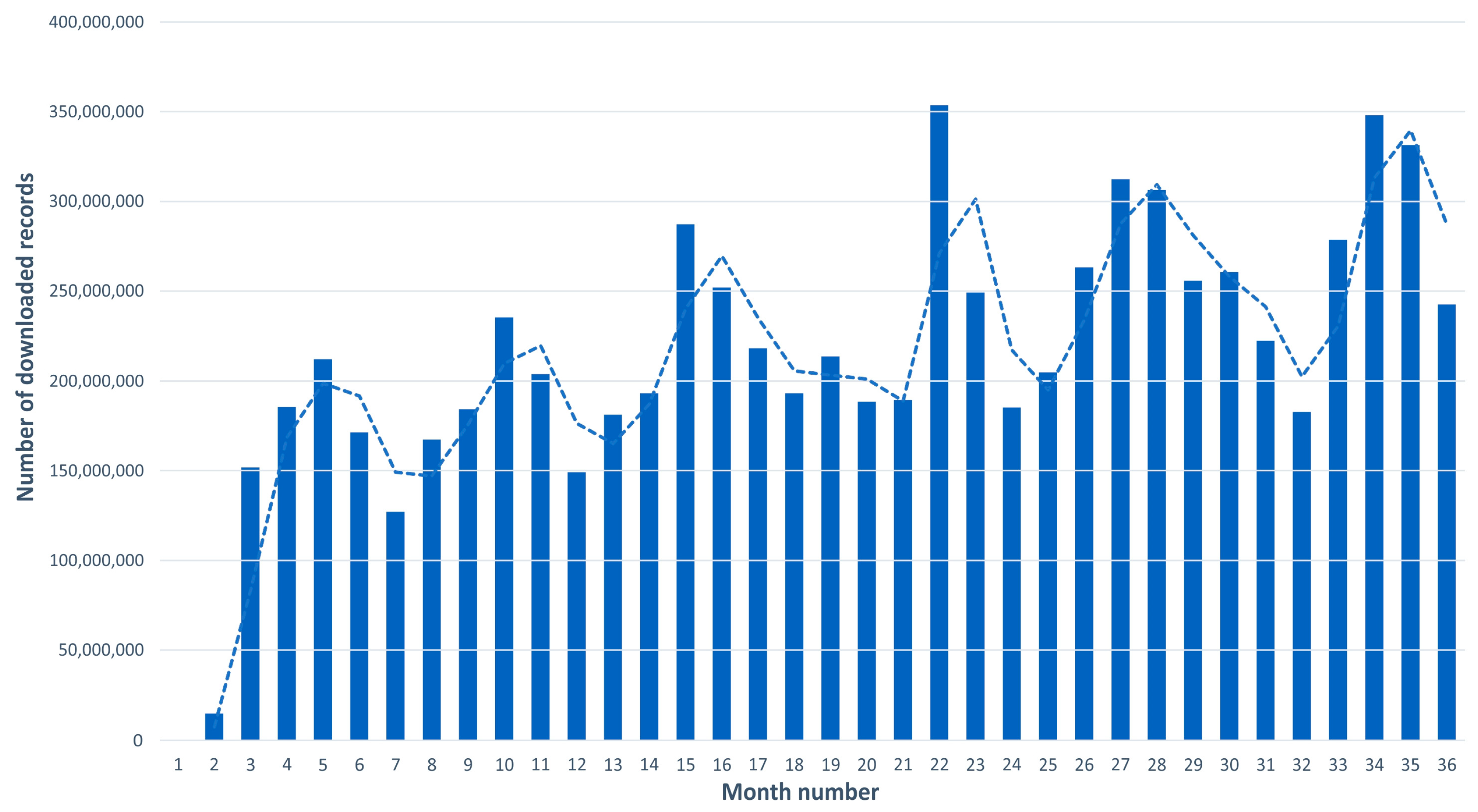
3.2.2. GBIF/FBAMU Data Download Statistics
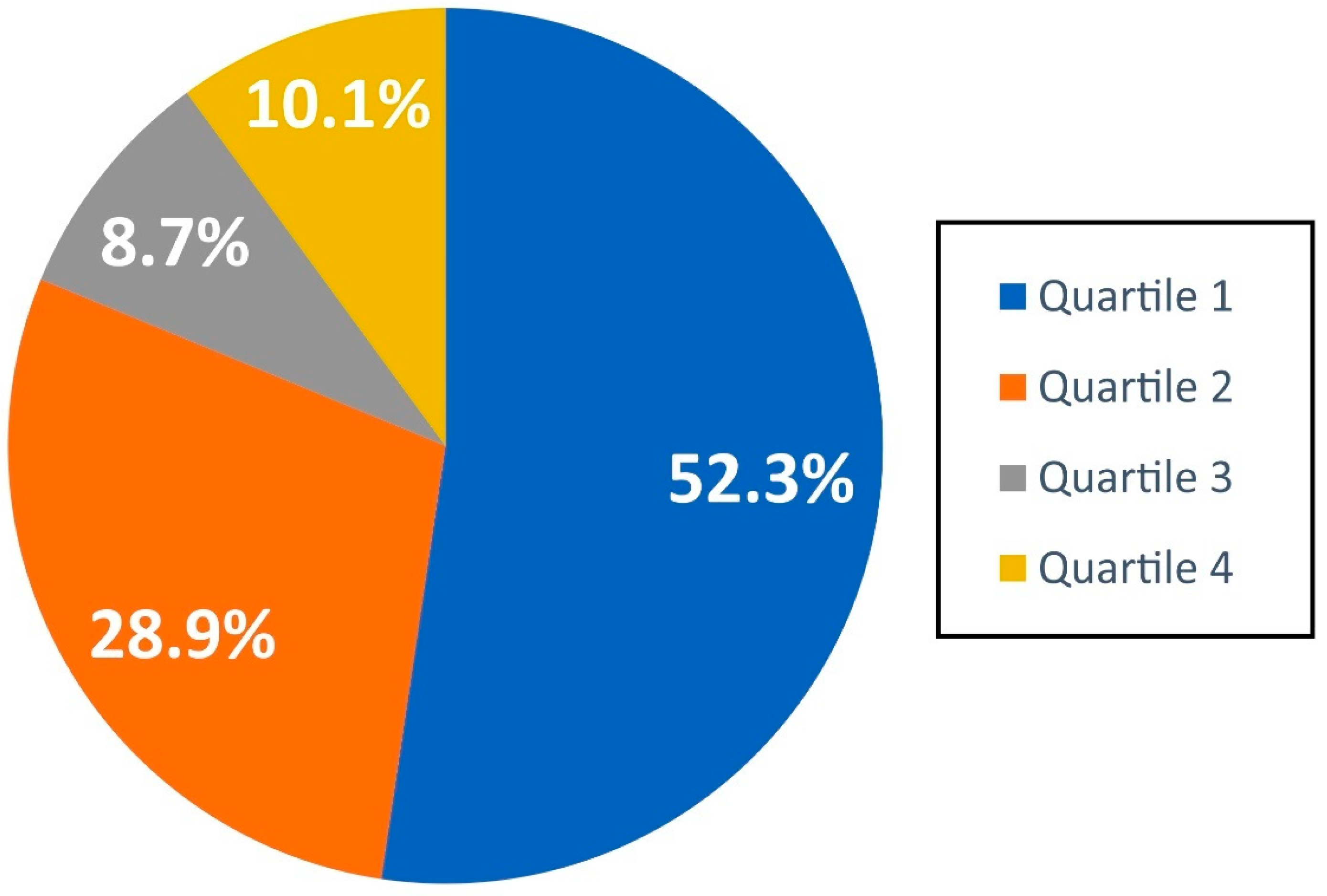
3.2.3. GBIF/FBAMU Data in Scientific Publications
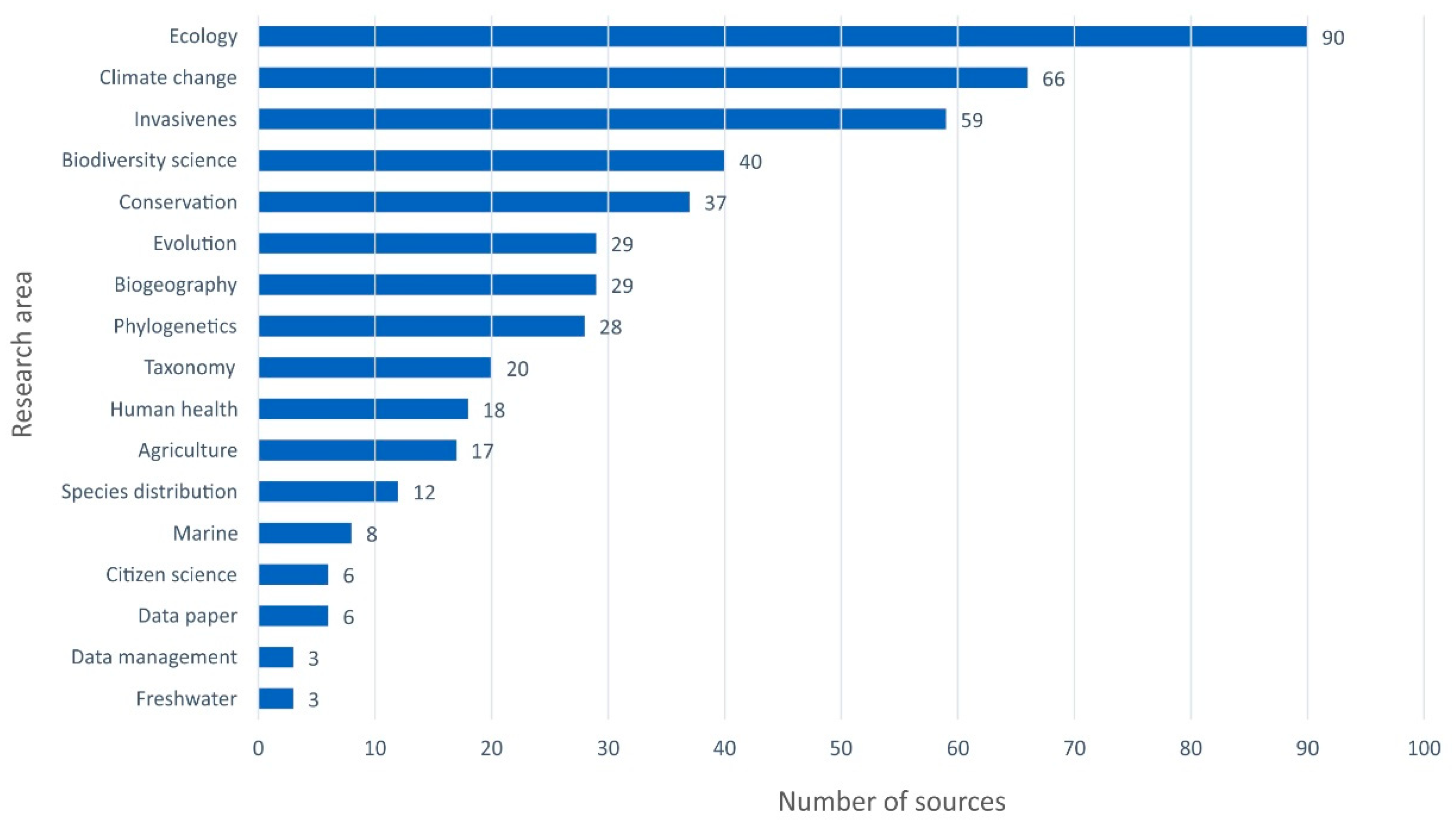
3.2.4. Phenomena and Processes Investigated Using GBIF/FBAMU Data
Morphology, Taxonomy, and Systematics
Biogeography
Ecology
Biodiversity Conservation
Paleobiology
Phylogeny and Evolution
Biodiversity Informatics
Applied Sciences and Other Research
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lane, M.A. Roles of Natural History Collections. Ann. Mo. Bot. Gard. 1996, 83, 536–545. [Google Scholar] [CrossRef]
- Krishtalka, L.; Humphrey, P.S. Can natural history museums capture the future? BioScience 2000, 50, 611–617. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. B 2003, 270, 313–321. [Google Scholar] [CrossRef]
- Suarez, A.V.; Tsutsui, N.D. The value of museum collections for research collections. BioScience 2004, 54, 66–74. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Gregory, T.R. The promise of DNA barcoding for taxonomy. Syst. Biol. 2005, 54, 852–859. [Google Scholar] [CrossRef] [PubMed]
- Savolainen, V.; Cowan, R.S.; Vogler, A.P.; Roderick, G.K.; Lane, R. Towards writing the encyclopaedia of life: An introduction to DNA barcoding. Phil. Trans. R. Soc. B 2005, 360, 1805–1811. [Google Scholar] [CrossRef] [PubMed]
- Wandeler, P.; Hoeckl, P.E.A.; Keller, L.F. Back to the future: Museum specimens in population genetics. Trends Ecol. Evol. 2007, 22, 634–642. [Google Scholar] [CrossRef]
- Borisenko, A.V.; Sones, J.E.; Hebert, P.D.N. The frontend logistics of DNA barcoding: Challenges and prospects. Mol. Ecol. Resour. 2009, 9, 27–34. [Google Scholar] [CrossRef]
- Pyke, G.H.; Ehrlich, P.R. Biological collections and ecological/environmental research: A review, some observations and a look to the future. Biol. Rev. 2010, 85, 247–266. [Google Scholar] [CrossRef]
- Smith, V.S.; Blagoderov, V. Bringing collections out of the dark. In No Specimen Left Behind: Mass Digitization of Natural History Collections; Blagoderov, V., Smith, V.S., Eds.; ZooKeys; Pensoft Publishers: Sofia, Bulgaria, 2012; Volume 209, pp. 1–6. [Google Scholar] [CrossRef]
- Nelson, G.; Paul, D.; Riccardi, G.; Mast, A. Five task clusters that enable efficient and effective digitization of biological collections. In No Specimen Left Behind: Mass Digitization of Natural History Collections; Blagoderov, V., Smith, V.S., Eds.; ZooKeys; Pensoft Publishers: Sofia, Bulgaria, 2012; Volume 209, pp. 19–45. [Google Scholar] [CrossRef]
- Page, L.M.; MacFadden, B.J.; Fortes, J.A.; Soltis, P.S.; Riccardi, G. Digitization of biodiversity collections reveals biggest data on biodiversity. BioScience 2015, 65, 841–842. [Google Scholar] [CrossRef]
- Cantrill, D. The Australasian Virtual Herbaria: Tracking data usage and benefits for biological collections. Appl. Plant Sci. 2018, 6, e1026. [Google Scholar] [CrossRef]
- Nowak, M.M.; Słupecka, K.; Jackowiak, B. Geotagging of natural history collections for reuse in environmental research. Ecol. Indic. 2021, 131, 108131. [Google Scholar]
- Soltis, P.S.; Nelson, G.; James, S.A. Green digitization: Online botanical collections data answering real-world questions. Appl. Plant Sci. 2018, 6, e1028. [Google Scholar] [CrossRef]
- Huang, T.; Chen, J.; Hummer, K.E.; Alice, L.A.; Wang, W.; He, Y.; Yu, S.; Yang, M.; Chai, T.; Zhu, X.; et al. Phylogeny of Rubus (Rosaceae): Integrating molecular and morphological evidence into an infrageneric revision. Taxon 2023, 72, 278–306. [Google Scholar] [CrossRef]
- Pérez-Escobar, O.A.; Bogarín, D.; Przelomska, N.A.; Ackerman, J.D.; Balbuena, J.A.; Bellot, S.; Bühlmann, R.P.; Cabrera, B.; Cano, J.A.; Charitonidou, M.; et al. The origin and speciation of orchids. New Phytol. 2024, 242, 700–716. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, H.; Ye, C.; Wang, Z.; Ma, C.; Lin, D.; Jin, X. Progress in systematics and biogeography of Orchidaceae. Plant Divers. 2024, 46, 425–434. [Google Scholar] [CrossRef]
- Liu, J.; Milne, R.I.; Zhu, G.-F.; Spicer, R.A.; Wambulwa, M.C.; Wu, Z.-Y.; Boufford, D.E.; Luo, Y.-H.; Provan, J.; Yi, T.-S.; et al. Name and scale matter: Clarifying the geography of Tibetan Plateau and adjacent mountain regions. Glob. Planet. Change 2022, 215, 103893. [Google Scholar] [CrossRef]
- Petrosyan, V.; Osipov, F.; Feniova, I.; Dergunova, N.; Warshavsky, A.; Khlyap, L.; Dzialowski, A. The TOP-100 most dangerous invasive alien species in Northern Eurasia: Invasion trends and species distribution modelling. NeoBiota 2023, 82, 23–56. [Google Scholar] [CrossRef]
- Gallardo, B.; Bacher, S.; Barbosa, A.M.; Gallien, L.; González-Moreno, P.; Martínez-Bolea, V.; Sorte, C.; Vimercati, G.; Vilà, M. Risks posed by invasive species to the provision of ecosystem services in Europe. Nat. Commun. 2024, 15, 2631. [Google Scholar] [CrossRef]
- Henriksen, M.V.; Arlé, E.; Pili, A.; Clarke, D.A.; García-Berthou, E.; Groom, Q.; Lenzner, B.; Meyer, C.; Seebens, H.; Tingley, R.; et al. Global indicators of the environmental impacts of invasive alien species and their information adequacy. Philos. Trans. R. Soc. B 2024, 379, 20230323. [Google Scholar] [CrossRef]
- Willis, C.G.; Ellwood, E.R.; Primack, R.B.; Davis, C.C.; Pearson, K.D.; Gallinat, A.S.; Yost, J.M.; Nelson, G.; Mazer, S.J.; Rossington, N.L.; et al. Old plants, new tricks: Phenological research using herbarium specimens. Trends Ecol. Evol. 2017, 32, 531–546. [Google Scholar] [CrossRef] [PubMed]
- Yost, J.M.; Sweeney, P.W.; Gilbert, E.; Nelson, G.; Guralnick, R.; Gallinat, A.S.; Ellwood, E.R.; Rossington, N.; Willis, C.G.; Blum, S.D.; et al. Digitization protocol for scoring reproductive phenology from herbarium specimens of seed plants. Appl. Plant Sci. 2018, 6, e1022. Available online: http://www.wileyonlinelibrary.com/journal/AppsPlantSci (accessed on 25 February 2025). [CrossRef]
- Contreras, D.L. A workflow and protocol describing the field to digitization process for new project-based fossil leaf collections. Appl. Plant Sci. 2018, 6, e1025. [Google Scholar] [CrossRef] [PubMed]
- Reichgelt, T.; Baumgartner, A.; Feng, R.; Willard, D.A. Poleward amplification, seasonal rainfall and forest heterogeneity in the Miocene of the eastern USA. Global Planet. Change 2023, 222, 104073. [Google Scholar] [CrossRef]
- Bonnet-Lebrun, A.-S.; Sweetlove, M.; Griffiths, H.J.; Sumner, M.; Provoost, P.; Raymond, B.; Ropert-Coudert, Y.; Van de Putte, A.P. Opportunities and limitations of large open biodiversity occurrence databases in the context of a Marine Ecosystem Assessment of the Southern Ocean. Front. Mar. Sci. 2023, 10, 1150603. [Google Scholar] [CrossRef]
- Li, J.; Wang, W.; Li, L.; Ding, Q.; Bai, M. Hexagon-based adaptive hierarchies for efficient point-in-spherical-polygon tests on GPUs. Int. J. Geogr. Inf. Sci. 2024, 38, 322–353. [Google Scholar] [CrossRef]
- Contreras-Díaz, R.G.; Nori, J.; Chiappa-Carrara, X.; Peterson, A.T.; Soberón, J.; Osorio-Olvera, L. Well-intentioned initiatives hinder understanding biodiversity conservation: Cloaked iNaturalist information for threatened species. Biol. Conserv. 2023, 282, 110042. [Google Scholar] [CrossRef]
- Botella, C.A.; Bonnet, J.P.; Monestiez, P.; Munoz, F. Species distribution modeling based on the automated identification of citizen observations. Appl. Plant Sci. 2018, 6, e1029. [Google Scholar] [CrossRef]
- Ballard, H.L.; Robinson, L.D.; Young, A.N.; Pauly, G.B.; Higgins, L.M.; Johnson, R.F.; Tweddle, J.C. Contributions to conservation outcomes by natural history museum-led citizen science: Examining evidence and next steps. Biol. Conserv. 2017, 208, 87–97. [Google Scholar] [CrossRef]
- Lendemer, J.; Thiers, B.; Monfils, A.K.; Zaspel, J.; Ellwood, E.R.; Bentley, A.; LeVan, K.; Bates, J.; Jennings, D.; Contreras, D.; et al. The Extended Specimen Network: A Strategy to Enhance US Biodiversity Collections, Promote Research and Education. BioScience 2020, 70, 23–30. [Google Scholar] [CrossRef]
- Jackowiak, B.; Lawenda, M.; Nowak, M.M.; Wolniewicz, P.; Błoszyk, J.; Urbaniak, M.; Szkudlarz, P.; Jędrasiak, D.; Wiland-Szymańska, J.; Bajaczyk, R.; et al. Open Access to the Digital Biodiversity Database: A Comprehensive Functional Model of the Natural History Collections. Diversity 2022, 14, 596. [Google Scholar] [CrossRef]
- What Is GBIF? Available online: https://www.gbif.org/what-is-gbif (accessed on 13 January 2025).
- Wheeler, Q.D. What is GBIF? BioScience 2004, 54, 717. [Google Scholar] [CrossRef]
- Flemons, P.; Guralnick, R.; Krieger, J.; Ranipeta, A.; Neufeld, D. A web-based GIS tool for exploring the world’s biodiversity: The Global Biodiversity Information Facility Mapping and Analysis Portal Application (GBIF-MAPA). Ecol. Inform. 2007, 2, 49–60. [Google Scholar] [CrossRef]
- Integrated Digitized Biocollections (iDigBio), the National Resource for Advancing Digitization of Biodiversity Collections (ADBC). Available online: https://www.idigbio.org/about-idigbio (accessed on 13 January 2025).
- What Is the ALA? Available online: https://www.ala.org.au/about-ala/ (accessed on 13 January 2025).
- Canobio Comisio’n Nacional para el Conocimiento y Uso de la Biodiversidad. Available online: https://www.gob.mx/conabio (accessed on 13 January 2025).
- FAPESP. Available online: http://splink.cria.org.br/ (accessed on 13 January 2025).
- The Distributed System of Scientific Collection. Available online: http://dissco.eu/ (accessed on 13 January 2025).
- South African National Biodiversity Institute. Available online: http://biodiversityadvisor.sanbi.org (accessed on 13 January 2025).
- Dauby, G.; Zaiss, R.; Blach-Overgaard, A.; Catarino, L.; Damen, T.; Deblauwe, V.; Dessein, S.; Dransfield, J.; Droissart, V.; Duarte, M.C.; et al. RAINBIO: A mega-database of tropical African vascular plants distributions. PhytoKeys 2016, 74, 1–18. [Google Scholar] [CrossRef]
- The China’s National Specimen Information Infrastructure. Available online: http://www.nsii.org.cn (accessed on 13 January 2025).
- Marsico, T.D.; Krimmel, E.R.; Carter, J.R.; Gillespie, E.L.; Lowe, P.D.; McCauley, R.; Morris, A.B.; Nelson, G.; Smith, M.; Soteropoulos, D.L.; et al. Small herbaria contribute unique biogeographic records to county, locality, and temporal scales. Am. J. Bot. 2020, 107, 1577–1587. [Google Scholar] [CrossRef]
- Jackowiak, B.; Błoszyk, J.; Celka, Z.; Konwerski, S.; Szkudlarz, P.; Wiland-Szymańska, J. The natural history collections of Adam Mickiewicz University in Poznań (Poland): An outline of their history and content. Biodivers. Res. Conserv. 2022, 65, 1–22. [Google Scholar] [CrossRef]
- Jackowiak, B.; Błoszyk, J.; Dylewska, M.; Nowak, M.M.; Szkudlarz, P.; Lawenda, M.; Meyer, N. Digitization of and online access to data from the natural history collections of Adam Mickiewicz University in Poznań: Assumptions and implementation of the AMUNATCOLL project. Biodivers. Res. Conserv. 2022, 65, 23–34. [Google Scholar] [CrossRef]
- Lawenda, M.; Wiland-Szymańska, J.; Nowak, M.M.; Jędrasiak, D.; Jackowiak, B. The Adam Mickiewicz University Nature Collections IT system (AMUNATCOLL): Metadata structure, database and operational procedures. Biodivers. Res. Conserv. 2022, 65, 35–48. [Google Scholar] [CrossRef]
- Nowak, M.M.; Lawenda, M.; Wolniewicz, P.; Urbaniak, M.; Jackowiak, B. The Adam Mickiewicz University Nature Collections IT system (AMUNATCOLL): Portal, mobile application and graphical interface. Biodivers. Res. Conserv. 2022, 65, 49–67. [Google Scholar] [CrossRef]
- AMUNATCOLL IT Website, Adam Mickiewicz University Natural Collections. Available online: https://www.amunatcoll.pl (accessed on 28 January 2025).
- GBIF. Global Biodiversity Information Facility. Available online: https://www.gbif.org (accessed on 28 January 2025).
- REpresentational State Transfer (REST). Available online: https://restfulapi.net/ (accessed on 5 February 2025).
- Biological Collection Access Service. Available online: https://www.biocase.org/ (accessed on 11 February 2025).
- Application Programming Interface. Available online: https://www.ibm.com/topics/api (accessed on 5 February 2025).
- AMUNATCOLL Mobile Application at Google Play. Available online: https://play.google.com/store/apps/details?id=pl.pcss.amunatcoll.mobile (accessed on 10 February 2025).
- AMUNATCOLL Mobile Application at App Store. Available online: https://apps.apple.com/pl/app/amunatcoll/id1523442673 (accessed on 10 February 2025).
- JWT—JSON Web Tokens. Available online: https://jwt.io/ (accessed on 20 January 2025).
- Integrated Digitized Biocollections. Available online: https://www.idigbio.org/ (accessed on 20 January 2025).
- BioCASe Provider Software. Available online: https://www.biocase.org/products/provider_software/ (accessed on 11 February 2025).
- Integrated Publishing Toolkit. Available online: https://ipt.gbif.org/manual/en/ipt/latest/ (accessed on 10 February 2025).
- Access to Biological Collections Data. Available online: https://abcd.tdwg.org/ (accessed on 10 February 2025).
- AMUNATCOLL IT PyWrapper Manual Query Form. Available online: https://rhus-76.man.poznan.pl/biocase/utilities/queryforms/qf_manual.cgi?dsa=amunatcoll (accessed on 10 February 2025).
- GBIF—Website of Adam Mickiewicz University in Poznań. Available online: https://www.gbif.org/publisher/5480dea7-2a71-409b-a832-cbc5f1b5a2e6 (accessed on 10 February 2025).
- GBIF—Literature Tracking. Available online: https://www.gbif.org/literature-tracking (accessed on 10 February 2025).
- Web of Science. Available online: https://clarivate.com/academia-government/scientific-and-academic-research/research-discovery-and-referencing/web-of-science/ (accessed on 20 January 2025).
- Farr, J.; Krisantini, S.; Fabillo, M. The leaf micromorphology and anatomy of gamba grass, Andropogon gayanus Kunth (Poaceae: Panicoideae). Aust. J. Bot. 2024, 72, BT23104. [Google Scholar] [CrossRef]
- Kalinina, L.B.; Ageev, D.V.; Bulyonkowa, T.M. Hohenbuehelia filicina sp. nov. (Agaricales, Basidiomycota), from Southwestern Siberia, Russia. Phytotaxa 2023, 600, 272–280. [Google Scholar] [CrossRef]
- Sokoloff, D.D.; Degtjareva, G.V.; Skaptsov, M.V.; Vislobokov, N.A.; Kirejtshuk, A.G.; Sennikov, A.N.; Severova, E.E.; Chepinoga, V.V.; Samigullin, T.H.; Valiejo-Roman, C.M.; et al. Diploids and tetraploids of Acorus (Acoraceae) in temperate Asia are pseudocryptic species with clear differences in micromorphology, DNA sequences and distribution patterns, but shared pollination biology. Taxon 2024, 73, 718–761. [Google Scholar] [CrossRef]
- Li, X.; Wiens, J.J. Estimating Global Biodiversity: The Role of Cryptic Insect Species. Syst. Biol. 2023, 72, 391–403. [Google Scholar] [CrossRef]
- Schiavinato, D.J.; Gutiérrez, D.G.; Bartoli, A. Tagetes dombeyi (Asteraceae, Tageteae), a new species from the Central Andes first collected in the 18th century. Rodriguésia 2023, 74, e00162023. [Google Scholar] [CrossRef]
- Stevanoski, I.; Kuzmanović, N.; Šatović, Z.; Liber, Z.; Radosavljević, I.; Lakušić, D. Campanula ramosissima subsp. megapetala, a new subspecies of Campanula sect. Decumbentes (Campanulaceae) revealed by morphological and genetic analyses. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2023, 158, 102–118. [Google Scholar] [CrossRef]
- Oliver, E.G.H.; Forshaw, N.; Oliver, I.M.; Volk, F.; Schumann, A.W.S.; Dorr, L.J.; Hoekstra, R.D.; Musker, S.D.; Nürk, N.M.; Pirie, M.D.; et al. Genus Erica: An Identification Aid Version 4.00. PhytoKeys 2024, 241, 143–154. [Google Scholar] [CrossRef]
- Krueger, T.; Robinson, A.; Bourke, G.; Fleischmann, A. Small Leaves, Big Diversity: Citizen Science and Taxonomic Revision Triples Species Number in the Carnivorous Drosera microphylla Complex (D. Section Ergaleium, Droseraceae). Biology 2023, 12, 141. [Google Scholar] [CrossRef]
- Tippery, N.P.; Gonzalez-Socoloske, D.; Leliaert, F.; Thompson, T.A.; Scatigna, A.V.; Souza, V.C. Systematics and biogeography of Bacopa (Plantaginaceae). Plant Syst. Evol. 2024, 310, 7. [Google Scholar] [CrossRef]
- Chvikov, V.S.; Khudych, A.S.; Akulov, O.Y.; Zinenko, O.I. Revision of some specimens of the genus Tulostoma (Agaricaceae) from Ukraine, deposited in the Herbarium of V.N. Karazin National University of Kharkiv (CWU). Ukr. Bot. J. 2022, 79, 84–90. [Google Scholar] [CrossRef]
- Geisler, L.; Hansen, A.; Kappel, S.; Josh, J.; Solodovnikov, A. Integrative taxonomy suggests species status for Philonthus sideropterus Kolenati, 1846 sp. propr., the former color form of Philonthus splendens (Fabricius, 1793) (Coleoptera: Staphylinidae). Integr. Syst. 2024, 7, 9–23. [Google Scholar] [CrossRef]
- Lima, L.V.; Almeida, T.E.; Kessler, M.; Rouhan, G.; Salino, A. (3006) Proposal to conserve the name Sticherus (Gleicheniaceae) with a conserved type. Taxon 2024, 73, 305–306. [Google Scholar] [CrossRef]
- Gabyshev, V.; Davydov, D.; Vilnet, A.; Sidelev, S.; Chernova, E.; Barinova, S.; Gabysheva, O.; Zhakovskaya, Z. Gloeotrichia cf. natans (Cyanobacteria) in the Continuous Permafrost Zone of Buotama River, Lena Pillars Nature Park, in Yakutia (Russia). Water 2023, 15, 2370. [Google Scholar] [CrossRef]
- Islomiddinov, Z.S.; Mustafaev, I.M.; Shirqulova, J.P.; Khabibullaev, B.S.; Lim, Y.W. The first record of Pisolithus arhizus (Sclerodermataceae, Basidiomycota) in Central Asia. Ukr. Bot. J. 2023, 80, 337–342. [Google Scholar] [CrossRef]
- Villalba-Alemán, J.; Mayorga, P.; Pinto, C.M.; Jaramillo Díaz, P. Confirmed presence of Clathrus columnatus Bosc (Phallales, Clathraceae) in the Galapagos Islands, Ecuador. Check List 2023, 19, 727–733. [Google Scholar] [CrossRef]
- Gületkin, N.; Ismailova, M.S.; Gületkin, L.; Yunakov, N.; Korytyaev, B.A. Notes on the distribution, food plants and morphology of Chlorophanus CR Sahlberg, 1823 species (Coleoptera: Curculionidae) in Turkey. J. Insect Biodivers. 2024, 49, 1–30. [Google Scholar] [CrossRef]
- Sutton, G.F.; van Steenderen, C.J.M.; Yell, L.D.; Canavan, K.; McConnachie, A.; Paterson, I.D. First record of an African grass-feeding wasp (Tetramesa; Eurytomidae) on the invasive grass Eragrostis curvula (African lovegrass; Poaceae) in Australia. BioInvasions Rec. 2023, 12, 673–680. [Google Scholar] [CrossRef]
- Martínez-Fonseca, J.G.; Trujillo, L.A.; Westeen, E.P.; Reid, F.A.; Hood, C.; Fernández-Mena, M.A.; GutiérrezLópez, L.E.; Molina-Loza, J.C.; Chambers, C.L. New departmental and noteworthy records of mammals (Mammalia, Theria) from Nicaragua. Check List 2024, 20, 706–720. [Google Scholar] [CrossRef]
- Gosline, G.; Bidault, E.; van der Burgt, X.; Cahen, D.; Challen, G.; Condé, N.; Couch, C.; Couvreur, T.L.P.; Dagallier, L.-P.M.J.; Darbyshire, I.; et al. A Taxonomically-verified and Vouchered Checklist of the Vascular Plants of the Republic of Guinea. Sci. Data 2023, 10, 327. [Google Scholar] [CrossRef]
- Örücü, Ö.K.; Azadi, H.; Arslan, E.S.; Aksoy, Ö.K.; Choobchian, S.; Nooghabi, S.N.; Stefanie, H.I. Predicting the distribution of European Hop Hornbeam: Application of MaxEnt algorithm and climatic suitability models. Eur. J. For. Res. 2023, 142, 579–591. [Google Scholar] [CrossRef]
- Walusiak, E.; Krztoń, W.; Cieślak, E.; Szczepaniak, M.; Wilk-Woźniak, E. Native recovery or expansive threat? Past and predicted distribution of Trapa natans L. s. l. on northern limit of species’ range—Handout for species management. Ecological Indic. 2024, 158, 111349. [Google Scholar] [CrossRef]
- Dutucu, A.A. Modeling for present and future (2100) possible distribution of Anadolu Chestnut (Castanea sativa) in Anatolia. J. Hum. Sci. 2023, 20, 446–460. [Google Scholar] [CrossRef]
- Kanmaz, O.; Şenel, T.; Dalfes, H.N. A Modeling Framework to Frame a Biological Invasion: Impatiens glandulifera in North America. Plants 2023, 12, 1433. [Google Scholar] [CrossRef]
- Sambou, M.; Koné, B.; Sane, S.; Vodounnon, M.E.J.; Diatta, A.A.; Diatta, L.; Sambou, B.; Diop, F.; Sambou, S.; Diatta, M.; et al. Impact of climate change on the habitat range and distribution of Cordyla pinnata, Faidherbia albida and Balanites aegyptiaca in Senegal. Model. Earth Syst. Environ. 2024, 10, 3137–3155. [Google Scholar] [CrossRef]
- Adamova, V.V.; Ukrainskiy, P.A. Modeling Current and Future Distribution of Cochlodina laminata in Eastern Europe under Climate Change. Diversity 2023, 15, 1155. [Google Scholar] [CrossRef]
- Hausdorf, B. Distribution patterns of established alien land snail species in the Western Palaearctic Region. NeoBiota 2023, 81, 1–32. [Google Scholar] [CrossRef]
- Atsawawaranunt, K.; Whibley, A.; Cain, K.E.; Major, R.E.; Santure, A.W. Projecting the current and potential future distribution of New Zealand’s invasive sturnids. Biol. Invasions 2024, 26, 1345–1366. [Google Scholar] [CrossRef]
- Qian, Q.; Xu, D.; Liao, W.; Zhuo, Z. Predicting the current and future suitable distribution range of Trilocha varians (Walker, 1855) (Lepidoptera: Bombycidae) in China. Bull. Entomol. Res. 2024, 114, 317–326. [Google Scholar] [CrossRef]
- Xian, X.; Zhao, H.; Humair, L.; Yang, N.; Li, J.; Weyl, P.; Liu, W. Niche shifts undermine the prediction performance of species distribution models: Estimating potentially suitable areas for Myriophyllum aquaticum at the global scale. Glob. Ecol. Conserv. 2023, 48, e02764. [Google Scholar] [CrossRef]
- Pérez-Suárez, M.; Ramírez-Albores, J.E.; Martínez-Campos, Á.R. Predicting the impacts of climate change on potential suitability habitats of three Juniperus trees in Mexico. Plant Ecol. 2024, 225, 37–51. [Google Scholar] [CrossRef]
- Liu, Y.; Heberling, K.M.; Wang, Z.; Soltis, D.E.; Soltis, P.S. Niche unfilling dominates the naturalization of species from intercontinentally disjunct genera. Glob. Ecol. Biogeogr. 2023, 32, 1977–1990. [Google Scholar] [CrossRef]
- Drummond, C.P.; Cochrane, T.S.; Sytsma, K.J. Western North American Plants Disjunct in the Great Lakes Region: 40 Years after Marquis and Voss. Int. J. Plant Sci. 2022, 183, 691–705. [Google Scholar] [CrossRef]
- Mitka, J.; Wróblewska, A.; Boroń, P.; Kucharzyk, S.; Stachurska-Swakoń, A. Perhaps there were northern refugia in LGM? The phylogeographic structure of the thermophilic tree Carpinus betulus in the Carpathian region. Sci. Total Environ. 2023, 905, 167214. [Google Scholar] [CrossRef]
- Howard, C.C.; Nanyeni, L.; Mollel, N.; Chuba, D.; Zuntini, A.R.; Malakasi, P.; Harvey, T.S.; Cellinese, N. From southern Africa and beyond: Historical biogeography of a monocotyledonous bulbous geophyte. J. Biogeogr. 2023, 50, 1623–1638. [Google Scholar] [CrossRef]
- García-Navarrete, P.G.; Escalante, T.; Espinosa, D.; Morrone, J.J. Evolutionary biogeography of the Revillagigedo Archipelago, Mexico. J. Nat. Hist. 2023, 57, 685–709. [Google Scholar] [CrossRef]
- Oliveira, U.; Azevedo, F.; Dias, A.; Almeida, A.C.S.; Senna, A.R.; Marques, A.C.; Rezende, D.; Hajdu, E.; Lopes-Filho, E.A.P.; Pitombo, F.B.; et al. Beta diversity and regionalization of the western Atlantic marine biota. J. Biogeogr. 2024, 51, 1469–1480. [Google Scholar] [CrossRef]
- Torres, A.F.; Wangensteen, O.S.; Renema, W.; Meyer, C.P.; Kendrich, I.; Fontanilla, C.; Todd, J.A. Global species hotspots and COI barcoding cold spots of marine Gastropoda. Biodivers. Conserv. 2024, 33, 2925–2947. [Google Scholar] [CrossRef]
- Krah, F.-S.; Büntgen, U.; Bässler, C. Temperature affects the timing and duration of fungal fruiting patterns across major terrestrial biomes. Ecol. Lett. 2023, 26, 1572–1583. [Google Scholar] [CrossRef]
- Kibria, M.G.; Tjoelker, M.G.; Marchin, R.M.; Arndt, S.K.; Rymer, P.D. Can species climate niche predict canopy growth, functional traits and phenotypic plasticity in urban trees? Urban For. Urban Green. 2024, 98, 128417. [Google Scholar] [CrossRef]
- Wan, Q.; Du, S.; Chen, Y.; Li, F.; Salah, R.; Njenga, M.N.; Li, J.; Wang, S. Ecological Niche Differentiation and Response to Climate Change of the African Endemic Family Myrothamnaceae. Plants 2024, 13, 1544. [Google Scholar] [CrossRef]
- Feng, L.; Tian, X.; El-Kassaby, Y.A.; Qiu, J.; Feng, Z.; Sun, J.; Wang, G.; Wang, T. Predicting suitable habitats of Melia azedarach L. in China using data mining. Sci. Rep. 2022, 12, 12617. [Google Scholar] [CrossRef]
- Ramírez-Ojeda, G.; Ramírez-Segura, E.; Barrera-Guzmán, L.Á.; Vázquez-González, A. Ecogeography and Climate Change in Forage Grasses from Arid and Semi-Arid Regions of Mexico. Grasses 2024, 3, 110–129. [Google Scholar] [CrossRef]
- Edwards-Calma, K.; Jiménez, L.; Zenil-Ferguson, R.; Heyduk, K.; Thomas, M.K.; Tribble, C.M. Conservation applications of niche modeling: Native and naturalized ferns may compete for limited Hawaiian dryland habitat. Appl. Plant Sci. 2024, 12, e11598. [Google Scholar] [CrossRef]
- Mathur, M.; Mathur, P. Ecological Niche Modelling of an Industrially Important Mushroom—Ganoderma lucidum (Leys.) Karsten: A Machine Learning Global Appraisal. J. Sci. Ind. Res. 2023, 82, 1231–1249. [Google Scholar] [CrossRef]
- Gao, H.; Wei, X.; Peng, Y.; Zhuo, Z. Predicting the Impact of Climate Change on the Future Distribution of Paederus fuscipes Curtis, 1826, in China Based on the MaxEnt Model. Insects 2024, 15, 437. [Google Scholar] [CrossRef]
- Wieners, M.; Thines, M.; Scholler, M. Hidden treasures—Historical specimens from the late blight pandemic discovered in the Herbarium of the State Museum of Natural History Karlsruhe. J. Plant Dis. Prot. 2024, 131, 1113–1119. [Google Scholar] [CrossRef]
- Strubbe, D.; Jiménez, L.; Barbosa, A.M.; Davis, A.J.; Lens, L.; Rahbek, C. Mechanistic models project bird invasions with accuracy. Nat. Commun. 2023, 14, 2520. [Google Scholar] [CrossRef]
- Louw, G.J.; Potgieter, L.J.; Richardson, D.M. Myoporum (Scrophulariaceae): Introduction, naturalization, and invasion of an enigmatic tree genus in South Africa. S. Afr. J. Bot. 2024, 168, 529–541. [Google Scholar] [CrossRef]
- Riera, M.; Pino, J.; Sáez, L.; Aymerich, P.; Melero, Y. Effect of introduction pathways on the invasion success of non-native plants along environmental gradients. Biological Invasions 2024, 26, 1561–1580. [Google Scholar] [CrossRef]
- Rodríguez-Merino, A. Identifying and Managing Areas under Threat in the Iberian Peninsula: An Invasion Risk Atlas for Non-Native Aquatic Plant Species as a Potential Tool. Plants 2023, 12, 3069. [Google Scholar] [CrossRef]
- Medzihorský, V.; Trombik, J.; Mally, R.; Turčáni, M.; Liebhold, A.M. Insect invasions track a tree invasion: Global distribution of black locust herbivores. J. Biogeogr. 2023, 50, 1285–1298. [Google Scholar] [CrossRef]
- Zhu, K.; Song, Y. Harnessing herbaria to advance plant phenology research under global change. New Phytol. 2023, 239, 2057–2059. [Google Scholar] [CrossRef] [PubMed]
- Kotilainen, A.; Mattila, A.L.; Møller, C.; Koivusaari, S.; Hyvärinen, M.T.; Hällfors, M.H. Higher thermal plasticity in flowering phenology increases flowering output. Ecol. Evol. 2024, 14, e11657. [Google Scholar] [CrossRef] [PubMed]
- Wilgan, R. High Species Diversity but Low Specificity to Ectomycorrhizal Tree Partners Exhibited by Native Truffle Species (Tuber spp., Pezizales) in Poland, Central Europe. Forests 2023, 14, 2407. [Google Scholar] [CrossRef]
- Barra, P.; Pereyra, P.J.; Gastaldi, M.; Saad, J.F.; Rodríguez, E.A.; Narvarte, M.A.; Calcagno, J.A. Intertidal populations of Ulva spp. and Undaria pinnatifida are good habitat providers for invertebrates but not for fish. Mar. Biol. 2023, 170, 91. [Google Scholar] [CrossRef]
- Dellinger, A.S.; Hamilton, A.M.; Wessinger, C.A.; Smith, S.D. Opposing Patterns of Altitude-Driven Pollinator Turnover in the Tropical and Temperate Americas. Am. Nat. 2023, 202, 152–165. [Google Scholar] [CrossRef]
- Minghetti, E.; Dellapé, P.M.; Maestro, M.; Montemayor, S.I. Evaluating the climatic suitability of Engytatus passionarius Minghetti et al. (Heteroptera, Miridae) as a biological control agent of the invasive stinking passion flower Passiflora foetida L. in Australia through ecological niche models. Biol. Control 2024, 191, 105461. [Google Scholar] [CrossRef]
- Gazol, A.; Valeriano, C.; Colangelo, M.; Ibáñez, R.; Valerio, M.; Rubio-Cuadrado, Á.; Camarero, J.J. Growth of tree (Pinus sylvestris) and shrub (Amelanchier ovalis) species is constrained by drought with higher shrub sensitivity in dry sites. Sci. Total Environ. 2024, 918, 170539. [Google Scholar] [CrossRef]
- Grossman, J.J.; Coe, H.B.; Fey, O.; Fraser, N.; Salaam, M.; Semper, C.; Williamson, C.G. Temperate woody species across the angiosperm phylogeny acquire tolerance to water deficit stress during the growing season. New Phytol. 2024, 24, 1981–1985. [Google Scholar] [CrossRef]
- Marks, R.A.; Delgado, P.; Makonya, G.M.; Cooper, K.; VanBuren, R.; Farrant, J.M. Higher order polyploids exhibit enhanced desiccation tolerance in the grass Microchloa caffra. J. Exp. Bot. 2024, 75, 3612–3623. [Google Scholar] [CrossRef]
- Riddell, E.A.; Mutanen, M.M.; Ghalambor, C.K. Hydric effects on thermal tolerance influence climate vulnerability of a high-latitude click beetle. Glob. Change Biol. 2023, 29, 5184–5198. [Google Scholar] [CrossRef]
- Hidalgo-Triana, N.; Solakis, A.; Casimiro-Soriguer, F.; Choe, H.; Navarro, T.; Pérez-Latorre, A.V.; Thorne, J.H. The high climate vulnerability of western Mediterranean forests. Sci. Total Environ. 2023, 895, 164983. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.; Zhu, Y.; Yang, X.; Eldridge, D.J. Predicted changes in distribution and grazing value of Stipa-based plant communities across the Eurasian steppe. J. Environ. Manag. 2024, 356, 120757. [Google Scholar] [CrossRef]
- Azevedo-Schmidt, L.A.; Currano, E.D.; Dunn, R.E.; Gjieli, E.; Pittermann, J.; Sessa, E.; Gill, J.L. Ferns as facilitators of community recovery following biotic upheaval. BioScience 2024, 74, 322–332. [Google Scholar] [CrossRef]
- Lajeunesse, A.; Fourcade, Y. Temporal analysis of GBIF data reveals the restructuring of communities following climate change. J. Anim. Ecol. 2023, 92, 391–402. [Google Scholar] [CrossRef]
- Arrogante-Funes, F.; Aguado, I.; Chuvieco, E. Global impacts of fire regimes on wildland bird diversity. Fire Ecol. 2024, 20, 25. [Google Scholar] [CrossRef]
- Federico, N.A.; Guralnik, R.P.; Belitz, M.W. Large uncertainty in trait responses across insects among overall declines in a subtropical city. Insect Conserv. Divers. 2024, 17, 312–323. [Google Scholar] [CrossRef]
- Yang, J.; Wu, Q.; Dakhil, M.A.; Halmy, M.W.A.; Bedair, H.; Fouad, M.S. Towards Forest Conservation Planning: How Temperature Fluctuations Determine the Potential Distribution and Extinction Risk of Cupressus funebris Conifer Trees in China. Forests 2023, 14, 2234. [Google Scholar] [CrossRef]
- Borgelt, J.; Dorber, M.; Høiberg, M.A.; Verones, F. More than half of data deficient species predicted to be threatened by extinction. Commun. Biol. 2022, 5, 679. [Google Scholar] [CrossRef]
- Carmona-Higuita, M.J.; Mendieta-Leiva, G.; Gómez-Díaz, J.A.; Villalobos, F.; Ramos, F.N.; Elias, J.P.C.; Jiménez-López, D.A.; Zuluaga, A.; Holst, B.; Kessler, M.; et al. Conservation status of vascular epiphytes in the neotropics. Biodivers. Conserv. 2024, 33, 51–71. [Google Scholar] [CrossRef]
- Raggi, L.; Zucchini, C.; Sayde, E.; Gigante, D.; Negri, V. Priority areas for the establishment of genetic reserves to actively protect key crop wild relative species in Italy. Glob. Ecol. Conserv. 2024, 50, e02836. [Google Scholar] [CrossRef]
- Mirea, M.D.; Miu, I.V.; Popescu, V.D.; Brodie, B.S.; Chiriac, S.; Rozylowicz, L. Priority conservation areas for protected saproxylic beetles in Romania under current and future climate scenarios. Biodivers. Conserv. 2024, 33, 2949–2973. [Google Scholar] [CrossRef]
- Kor, L.; Diazgranados, M. Identifying important plant areas for useful plant species in Colombia. Biol. Conserv. 2023, 284, 110187. [Google Scholar] [CrossRef]
- Trájer, A.J. Reconstruction of palaeoenvironmental conditions that led to the formation of Eocene sub-bituminous coal seams in the Hungarian Paleogene Basin. Rev. Palaeobot. Palynol. 2024, 323, 105080. [Google Scholar] [CrossRef]
- Poore, C.; Jud, N.A.; Gandolfo, M.A. Fossil fruits from the early Paleocene of Patagonia, Argentina reveal west Gondwanan history of Icacinaceae. Rev. Palaeobot. Palynol. 2023, 317, 104940. [Google Scholar] [CrossRef]
- Geier, C.; Bouchal, J.M.; Ulrich, S.; Uhl, D.; Wappler, T.; Wedmann, S.; Zetterg, R.; Schőnenberger, J.; Grímsson, F. Potential pollinators and paleoecological aspects of Eocene Ludwigia (Onagraceae) from Eckfeld, Germany. Palaeoworld 2024, 33, 1079–1104. [Google Scholar] [CrossRef]
- Molnár, Á.P.; Demeter, L.; Biró, M.; Chytrý, M.; Bartha, S.; Gantuya, B.; Molnár, Z. Is there a massive glacial–Holocene flora continuity in Central Europe? Biol. Rev. 2023, 98, 2307–2319. [Google Scholar] [CrossRef] [PubMed]
- Wuttke, M.; Poschmann, M.J.; Wappler, T.; Bouchal, J.M.; Geier, C.; Ulrich, S.; Grímsson, F. Pollen-feeding in a giant pelobatid tadpole from the late Oligocene of Enspel, Germany. Palaeobiodiversity Palaeoenvironments 2024, 104, 999–1026. [Google Scholar] [CrossRef]
- Boyko, J.D.; Hagen, E.R.; Beaulieu, J.M.; Vasconcelos, T. The evolutionary responses of life-history strategies to climatic variability in flowering plants. New Phytol. 2023, 240, 1587–1600. Available online: https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.18971 (accessed on 10 February 2025).
- Yang, X.; Zhang, X.; Xue, T.; Zhang, X.; Yang, F.; Yu, J.; Janssens, S.B.; Bussman, R.W.; Yu, S. Phylogenomics and historical biogeography of Hydrangeeae (Hydrangeaceae) elucidate the effects of geologic and climatic dynamics on diversification. Proc. R. Soc. B 2023, 290, 20230659. [Google Scholar] [CrossRef]
- Karunarathne, P.; Zhou, Q.; Lascoux, M.; Milesi, P. Hybridization mediated range expansion and climate change resilience in two keystone tree species of boreal forests. Glob. Change Biol. 2024, 30, e17262. [Google Scholar] [CrossRef]
- Ziegler, C.; Martínez, E.J.; Honfi, A.I.; Reutemann, A.V. Discovery of natural Paspalum L. (Poaceae) triploid hybrids near sympatric populations of Paspalum urvillei Steud. and species of Paniculata group in northeastern Argentina. Euphytica 2024, 220, 78. [Google Scholar] [CrossRef]
- Bharti, D.K.; Pawar, P.Y.; Edgecombe, G.D.; Joshi, J. Genetic diversity varies with species traits and latitude in predatory soil arthropods (Myriapoda: Chilopoda). Glob. Ecol. Biogeogr. 2023, 32, 1508–1521. [Google Scholar] [CrossRef] [PubMed]
- Jahanshiri, E.; Azam-Ali, S.; Gregory, P.J.; Wimalasiri, E.M. A Shortlisting Framework for Crop Diversification in the United Kingdom. Agriculture 2023, 13, 787. [Google Scholar] [CrossRef]
- Shumskaya, M.; Filippova, N.; Lorentzen, L.; Blue, S.; Andrew, C.; Lorusso, N.S. Citizen science helps in the study of fungal diversity in New Jersey. Sci. Data 2023, 10, 10. [Google Scholar] [CrossRef]
- Daru, B.H.; Rodriguez, J. Mass production of unvouchered records fails to represent global biodiversity patterns. Nat. Ecol. Evol. 2023, 7, 816–831. [Google Scholar] [CrossRef]
- Díaz-Calafat, J.; Jaume-Ramis, S.; Soacha, K.; Álvarez, A.; Piera, J. Revealing biases in insect observations: A comparative analysis between academic and citizen science data. PLoS ONE 2024, 19, e0305757. [Google Scholar] [CrossRef]
- García-Roselló, E.; González-Dacosta, J.; Lobo, J.M. The biased distribution of existing information on biodiversity hinders its use in conservation, and we need an integrative approach to act urgently. Biol. Conserv. 2023, 283, 110118. [Google Scholar] [CrossRef]
- Serra-Diaz, J.M.; Borderieux, J.; Maitner, B.; Boonman, C.C.; Park, D.; Guo, W.Y.; Callebaut, A.; Enquist, B.E.; Svenning, J.-C.; Merow, C. occTest: An integrated approach for quality control of species occurrence data. Glob. Ecol. Biogeogr. 2024, 33, e13847. [Google Scholar] [CrossRef]
- de Melo, P.H.A.; Bystriakova, N.; Lucas, E.; Monro, A.K. A new R package to parse plant species occurrence records into unique collection events efficiently reduces data redundancy. Sci. Rep. 2024, 14, 5450. [Google Scholar] [CrossRef]
- Zhang, H.T.; Guo, W.Y.; Wang, W.T. The dimensionality reductions of environmental variables have a significant effect on the performance of species distribution models. Ecol. Evol. 2023, 13, e10747. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Yu, S.E.; Wang, W.L.; Yang, L.E.; Lu, Q.Q.; Xie, C.T.; Liu, T.; Dong, Y.W. Temperature sensitivity of marine macroalgae for aquaculture in China. Aquaculture 2023, 567, 739262. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); de la Peña, E.; Petri, C.; Diaz-Pendón, J.A.; Hormaza, J.I.; Romero, H.; Hoedekie, A.; Maertens, F.; Debode, J.; Graziosi, I.; et al. Characterisation of fruit trees in the EU: A tool for crop-based survey of Union quarantine pests. EFSA Support. Publ. 2024, 21, 8823E. [Google Scholar] [CrossRef]
- Chemura, A.; Gleixner, S.; Gornott, C. Dataset of the suitability of major food crops in Africa under climate change. Sci. Data 2024, 11, 294. [Google Scholar] [CrossRef]
- Barve, N.; Ashraf, U.; Barve, V.; Cobos, M.E.; Nuñez-Penichet, C.; Peterson, A.T. Revisiting plant hardiness zones to include multiple climatic stress dimensions. Iscience 2024, 27, 10. [Google Scholar] [CrossRef]
- Lahlou, R.A.; Samba, N.; Soeiro, P.; Alves, G.; Gonçalves, A.C.; Silva, L.R.; Silvestre, S.; Rodilla, J.; Ismael, M.I. Thymus hirtus Willd. ssp. algeriensis Boiss. and Reut: A Comprehensive Review on Phytochemistry, Bioactivities, and Health-Enhancing Effects. Foods 2022, 11, 3195. [Google Scholar] [CrossRef]
- Nersezashvili, M.; Skalicka-Woźniak, K.; Berashvili, D. Phytochemical and Pharmaceutical Characterization of Seseli L. Species From Georgia. Acta Pol. Pharm. 2024, 81, 35. [Google Scholar] [CrossRef]
- Gan, Z.; Fang, X.; Yin, C.; Tian, Y.; Zhang, L.; Zhong, X.; Jiang, G.; Tao, A. Extraction, purification, structural characterization, and bioactivities of the genus Rhodiola L. polysaccharides: A review. Int. J. Biol. Macromol. 2024, 276, 133614. [Google Scholar] [CrossRef]
- Klages, J.P.; Hillenbrand, C.D.; Bohaty, S.M.; Salzmann, U.; Bickert, T.; Lohmann, G.; Knahl, H.S.; Gierz, P.; Niu, L.; Titschack, J.; et al. Ice sheet–free West Antarctica during peak early Oligocene glaciation. Science 2024, 385, 322–327. [Google Scholar] [CrossRef]
- Gavashelishvili, A.; Chukhua, M.; Sakhltkhutsishvili, K.; Koptekin, D.; Somel, M. The time and place of origin of South Caucasian languages: Insights into past human societies, ecosystems and human population genetics. Sci. Rep. 2023, 13, 21133. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Journal Name | JCR Category | JIF 5-Year | No. of Papers |
|---|---|---|---|
| New Phytologist | Plant sciences | 10.5 | 9 |
| Science of The Total Environment | Environmental sciences | 9.6 | 8 |
| Global Change Biology | Biodiversity conservation | 12.3 | 7 |
| Nature Communications | Multidisciplinary sciences | 17.0 | 7 |
| Scientific Data | Multidisciplinary sciences | 10.8 | 5 |
| Ecology Letters | Ecology | 9.8 | 3 |
| Nature Plants | Plant sciences | 18.6 | 3 |
| Nature Ecology & Evolution | Ecology | 16.9 | 2 |
| PNAS | Multidisciplinary sciences | 12.0 | 2 |
| Biological Reviews | Biology | 12.8 | 1 |
| BioScience | Biology | 9.7 | 1 |
| Ecological Monographs | Ecology | 9.0 | 1 |
| Journal of Integrative Plant Biology | Biochemistry & molecular biology | 9.3 | 1 |
| International Journal of Extreme Manufacturing | Engineering, manufacturing | 13.3 | 1 |
| Nature Climate Change | Environmental sciences | 31.4 | 1 |
| Nature Sustainability | Environmental sciences | 29.2 | 1 |
| Systematic Biology | Evolutionary biology | 11.0 | 1 |
| Water Research | Engineering, environmental | 12.2 | 1 |
| Subject Areas | No. of Papers | Phenomena and Processes * |
|---|---|---|
| Agriculture | 11 | Agriculture’s environmental footprint · Changes in species ranges (2) · Climate change adaptation · Climate change · Climatic niche dynamics · Epidemic spread · Open-access database · Plant hardiness zones · Regulated pests · Temperature sensitivity |
| Biodiversity conservation | 22 | Biodiversity loss (13) · Biological invasions · Conservation prioritization · Data density · Ecoregions · In situ conservation · Protected areas · Restoration · Spatial conservation (2) · |
| Biodiversity informatics | 37 | Big data · Biodiversity hotspot · Biodiversity loss (2) · Changes in species ranges · Climate change · Completeness and geographical biases · Data analysis (9) · Data cleaning · Database (4)· Fungal diversity · Geospatial suitability model · Global collection · Imperfect data · Knowledge gaps (4) · Plant diversity · Potential of the data (2) · Species distribution modeling (5) |
| Biogeography | 81 | Animal diversity · Biodiversity data (2) · Biodiversity hotspot (3) · Biodiversity loss (2) · Biogeographical patterns (13) · Biological invasions (8) · Biome evolution (2) · Changes in species ranges (17) · Changes in flora and vegetation · Checklists · Climate change adaptation · Climatic niche dynamics (4) · Cultural evolution · Database (2) · Disjunct distribution (2) · Geographical names · Interactions between organisms · Mechanisms of evolution · New record (6) · Plant diversity · Regionalization (2) · Relicts · Species distribution modeling (7) · Vicariance |
| Climate science | 1 | Greenhouse–icehouse transition |
| Ecology | 100 | Biodiversity loss (3) · Bioindication (3) · Biological invasions (15) · Carbon accumulation (2) · Changes in species ranges (3) · Changes in flora and Vegetation (6] · Climate change adaptation (8) · Climatic niche dynamics (22) · Community changes (6) · Data analysis · Diet · Dietary or habitat needs · Environmental changes (2) · Flora diversity · Forest management · Interactions between organisms (12) · Mechanisms of evolution · Organisms and habitat (5) · Remediation · Restoration of degraded sites · Spatial patterns · Toxicity · Transplantation · Tree mortality · Weed biocontrol |
| Human health | 16 | Bioactivities (7) · Biological invasions (2) · Bioprospecting · Database · Epidemic modeling · Metabolic syndrome · Schistosomatoza · Specialized metabolism · Toxicity |
| Linguistic phylogeny | 1 | Changes in species ranges |
| Morphology | 2 | Leaf characteristics · Shape modeling |
| Paleobiology | 8 | Environmental changes (2) · Fossil fruits · Fossil plants · Interactions between organisms · New record · Refugia · Vegetation reconstruction |
| Phylogeny and evolution | 34 | Biogeographical patterns · Climatic niche dynamics · Conservation prioritization · Convergent evolution · Evolutionary radiations · Genetic variability (3) · Hybridization (2) · Interactions between organisms (3) · Mechanisms of evolution (8) · Monophyly · Mutation pressure · Origin and speciation (6) · Peripheral populations · Polyploidy (2) · Reticulate evolution · Species distribution model |
| Taxonomy and systematics | 11 | Cryptic species (2) · New taxon · Nomenclatural issue · Taxon identification (4) · Taxonomical status · Typification · Variability |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jackowiak, B.; Lawenda, M. How Does Sharing Data from Research Institutions on Global Biodiversity Information Facility Enhance Its Scientific Value? Diversity 2025, 17, 221. https://doi.org/10.3390/d17040221
Jackowiak B, Lawenda M. How Does Sharing Data from Research Institutions on Global Biodiversity Information Facility Enhance Its Scientific Value? Diversity. 2025; 17(4):221. https://doi.org/10.3390/d17040221
Chicago/Turabian StyleJackowiak, Bogdan, and Marcin Lawenda. 2025. "How Does Sharing Data from Research Institutions on Global Biodiversity Information Facility Enhance Its Scientific Value?" Diversity 17, no. 4: 221. https://doi.org/10.3390/d17040221
APA StyleJackowiak, B., & Lawenda, M. (2025). How Does Sharing Data from Research Institutions on Global Biodiversity Information Facility Enhance Its Scientific Value? Diversity, 17(4), 221. https://doi.org/10.3390/d17040221