
Genomic, Epigenomic, Transcriptomic, Proteomic and Metabolomic Approaches in Atopic Dermatitis

Abstract
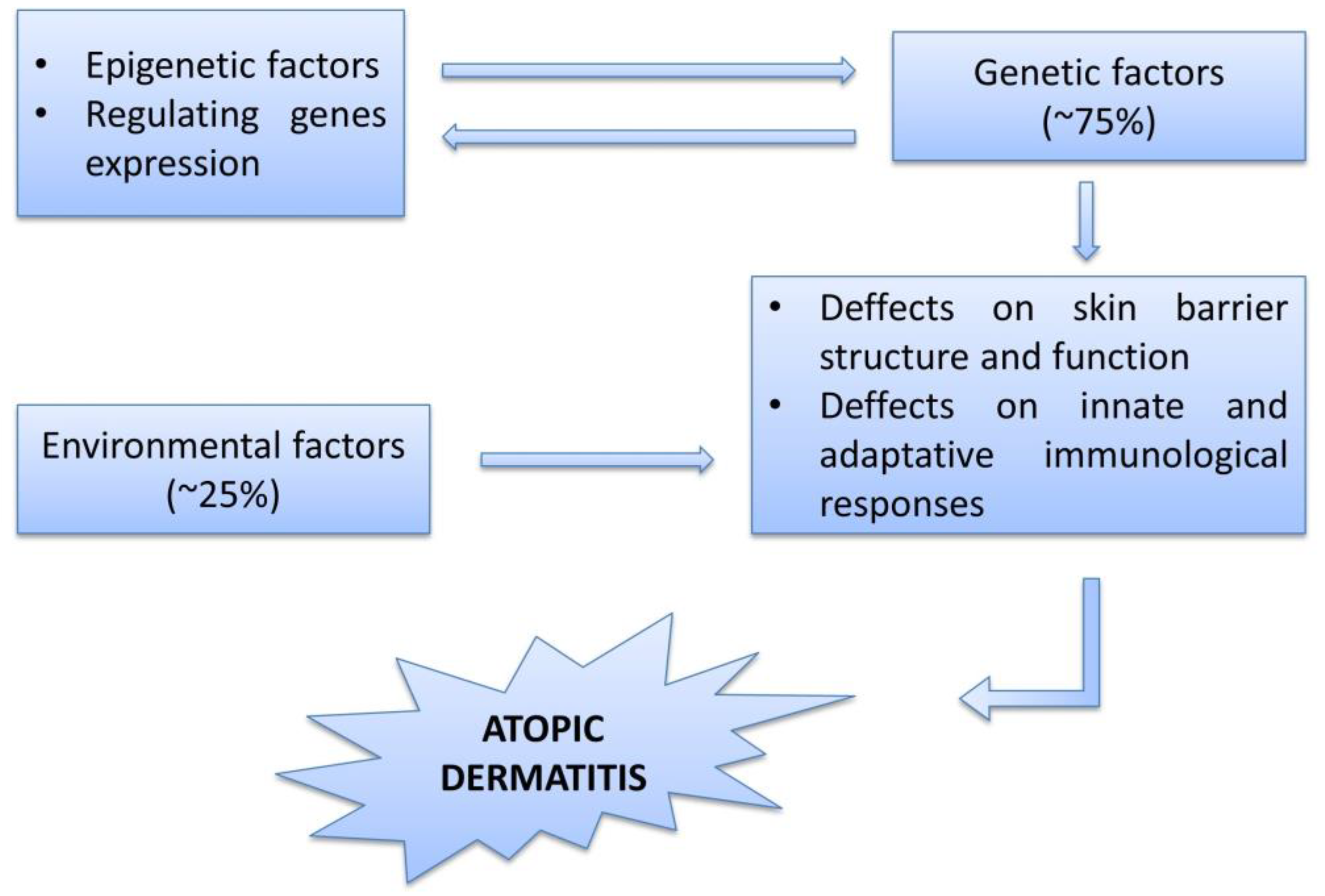
:1. Introduction
2. Etiology, Pathogenesis, Physiopathology
3. The Genome
4. The Epigenome
5. The Transcriptome, Proteome and Metabolome
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Han, H.; Roan, F.; Ziegler, S.F. The Atopic March: Current Insights into Skin Barrier Dysfunction and Epithelial Cell-Derived Cytokines. Immunol. Rev. 2017, 278, 116–130. [Google Scholar] [CrossRef] [PubMed]
- Aberg, N.; Hesselmar, B.; Aberg, B.; Eriksson, B. Increase of Asthma, Allergic Rhinitis and Eczema in Swedish Schoolchildren between 1979 and 1991. Clin. Exp. Allergy 1995, 25, 815–819. [Google Scholar] [CrossRef] [PubMed]
- Burr, M.L.; Butland, B.K.; King, S.; Vaughan-Williams, E. Changes in Asthma Prevalence: Two Surveys 15 Years Apart. Arch. Dis. Child. 1989, 64, 1452–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sears, M.R.; Rea, H.H.; Rothwell, R.P.; O’Donnell, T.V.; Holst, P.E.; Gillies, A.J.; Beaglehole, R. Asthma Mortality: Comparison between New Zealand and England. Br. Med. J. (Clin. Res. Ed.) 1987, 294, 646. [Google Scholar] [CrossRef] [Green Version]
- Eichenfield, L.F.; Hanifin, J.M.; Beck, L.A.; Lemanske, R.F., Jr.; Sampson, H.A.; Weiss, S.T.; Leung, D.Y. Atopic Dermatitis and Asthma: Parallels in the Evolution of Treatment. Pediatrics 2003, 111, 608–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez Carrera, Y.I.; Al Hammadi, A.; Huang, Y.-H.; Llamado, L.J.; Mahgoub, E.; Tallman, A.M. Epidemiology, Diagnosis, and Treatment of Atopic Dermatitis in the Developing Countries of Asia, Africa, Latin America, and the Middle East: A Review. Dermatol. Ther. 2019, 9, 685–705. [Google Scholar] [CrossRef] [Green Version]
- Kolb, L.; Ferrer-Bruker, S.J. StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Ständer, S. Atopic Dermatitis. N. Engl. J. Med. 2021, 384, 1136–1143. [Google Scholar] [CrossRef]
- McKenzie, C.; Silverberg, J.I. The Prevalence and Persistence of Atopic Dermatitis in Urban United States Children. Ann. Allergy Asthma Immunol. 2019, 123, 173–178.e1. [Google Scholar] [CrossRef]
- Ardern-Jones, M.R.; Flohr, C.; Reynolds, N.J.; Harwood, C.A. Rook’s Textbook of Dermatology, 9th ed.; Griffiths, C.E., Bleiker, T., Chalmers, R., Creamer, D., Eds.; John Wiley & Sons: Oxford, UK, 2016. [Google Scholar]
- Torres, T.; Ferreira, E.O.; Gonçalo, M.; Mendes-Bastos, P.; Selores, M.; Filipe, P. Update on Atopic Dermatitis. Acta Med. Port. 2019, 32, 606–613. [Google Scholar] [CrossRef] [Green Version]
- Nutten, S. Atopic Dermatitis: Global Epidemiology and Risk Factors. Ann. Nutr. Metab. 2015, 66 (Suppl. S1), 8–16. [Google Scholar] [CrossRef]
- Weidinger, S.; Beck, L.A.; Bieber, T.; Kabashima, K.I.A. Atopic Dermatitis. Nat. Rev. Dis. Prim. 2018, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Expert Perspectives on Management of Moderate-to-Severe Atopic Dermatitis: A Multidisciplinary Consensus Addressing Current and Emerging Therapies|Elsevier Enhanced Reader. Available online: https://reader.elsevier.com/reader/sd/pii/S2213219817306153?token=2F2F5707254043C4CF0CD6D299DE60FF94511623AC54E7B725F343D5916329C140A05D9E0700247768E5549B4BF4A5EC&originRegion=eu-west-1&originCreation=20220121141803 (accessed on 21 January 2022).
- Wollenberg, A.; Barbarot, S.; Bieber, T.; Christen-Zaech, S.; Deleuran, M.; Fink-Wagner, A.; Gieler, U.; Girolomoni, G.; Lau, S.; Muraro, A.; et al. Consensus-Based European Guidelines for Treatment of Atopic Eczema (Atopic Dermatitis) in Adults and Children: Part I. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 657–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deckers, I.A.G.; McLean, S.; Linssen, S.; Mommers, M.; van Schayck, C.P.; Sheikh, A. Investigating International Time Trends in the Incidence and Prevalence of Atopic Eczema 1990-2010: A Systematic Review of Epidemiological Studies. PLoS ONE 2012, 7, e39803. [Google Scholar] [CrossRef] [Green Version]
- Abuabara, K.; Yu, A.M.; Okhovat, J.P.; Allen, I.E.; Langan, S.M. The Prevalence of Atopic Dermatitis beyond Childhood: A Systematic Review and Meta-Analysis of Longitudinal Studies. Allergy Eur. J. Allergy Clin. Immunol. 2018, 73, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Silverberg, J.I.; Hanifin, J.M. Adult Eczema Prevalence and Associations with Asthma and Other Health and Demographic Factors: A US Population-Based Study. J. Allergy Clin. Immunol. 2013, 132, 1132–1138. [Google Scholar] [CrossRef]
- Barbarot, S.; Auziere, S.; Gadkari, A.; Girolomoni, G.; Puig, L.; Simpson, E.L.; Margolis, D.J.; de Bruin-Weller, M.; Eckert, L. Epidemiology of Atopic Dermatitis in Adults: Results from an International Survey. Allergy 2018, 73, 1284–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DaVeiga, S.P. Epidemiology of Atopic Dermatitis: A Review. Allergy Asthma Proc. 2012, 33, 227–234. [Google Scholar] [CrossRef]
- Kapur, S.; Watson, W.; Carr, S. Atopic Dermatitis. Allergy Asthma. Clin. Immunol. 2018, 14, 52. [Google Scholar] [CrossRef] [Green Version]
- Odhiambo, J.A.; Williams, H.C.; Clayton, T.O.; Robertson, C.F.; Asher, M.I.; ISAAC Phase Three Study Group. Global Variations in Prevalence of Eczema Symptoms in Children from ISAAC Phase Three. J. Allergy Clin. Immunol. 2009, 124, 1251–1258. [Google Scholar] [CrossRef]
- Davies, E.; Rogers, N.K.; Lloyd-Lavery, A.; Grindlay, D.J.C.; Thomas, K.S. What’s New in Atopic Eczema? An Analysis of Systematic Reviews Published in 2015. Part 1: Epidemiology and Methodology. Clin. Exp. Dermatol. 2018, 43, 375–379. [Google Scholar] [CrossRef]
- Waldman, A.R.; Ahluwalia, J.; Udkoff, J.; Borok, J.F.; Eichenfield, L.F. Atopic Dermatitis. Pediatr. Rev. 2018, 39, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Kantor, R.; Silverberg, J.I. Environmental Risk Factors and Their Role in the Management of Atopic Dermatitis. Expert Rev. Clin. Immunol. 2017, 13, 15–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomon, I.; Ilie, M.; Draghici, C.; Voiculescu, V.; Căruntu, C.; Boda, D.; Zurac, S. The Impact of Lifestyle Factors on Evolution of Atopic Dermatitis: An Alternative Approach (Review). Exp. Ther. Med. 2019, 17, 1078–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, S.J.; Elias, M.S.; Bradley, M. Genetics in Atopic Dermatitis: Historical Perspective and Future Prospects. Acta Derm. Venereol. 2020, 100, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.N.A.; Irvine, A.D.; Terron-Kwiatkowski, A.; Zhao, Y.; Liao, H.; Lee, S.P.; Goudie, D.R.; Sandilands, A.; Campbell, L.E.; Smith, F.J.D.; et al. Common Loss-of-Function Variants of the Epidermal Barrier Protein Filaggrin Are a Major Predisposing Factor for Atopic Dermatitis. Nat. Genet. 2006, 38, 441–446. [Google Scholar] [CrossRef]
- Fishbein, A.B.; Silverberg, J.I.; Wilson, E.J.; Ong, P.Y. Update on Atopic Dermatitis: Diagnosis, Severity Assessment, and Treatment Selection. J. Allergy Clin. Immunol. Pract. 2020, 8, 91–101. [Google Scholar] [CrossRef]
- Czarnowicki, T.; Krueger, J.G.; Guttman-Yassky, E. Novel Concepts of Prevention and Treatment of Atopic Dermatitis through Barrier and Immune Manipulations with Implications for the Atopic March. J. Allergy Clin. Immunol. 2017, 139, 1723–1734. [Google Scholar] [CrossRef] [Green Version]
- Tran, M.M.; Lefebvre, D.L.; Dharma, C.; Dai, D.; Lou, W.Y.W.; Subbarao, P.; Becker, A.B.; Mandhane, P.J.; Turvey, S.E.; Sears, M.R.; et al. Predicting the Atopic March: Results from the Canadian Healthy Infant Longitudinal Development Study. J. Allergy Clin. Immunol. 2018, 141, 601–607.e8. [Google Scholar] [CrossRef] [Green Version]
- Eller, E.; Kjaer, H.F.; Høst, A.; Andersen, K.E.; Bindslev-Jensen, C. Development of Atopic Dermatitis in the DARC Birth Cohort. Pediatr. Allergy Immunol. 2010, 21, 307–314. [Google Scholar] [CrossRef]
- Andersen, Y.M.F.; Egeberg, A.; Skov, L.; Thyssen, J.P. Comorbidities of Atopic Dermatitis: Beyond Rhinitis and Asthma. Curr. Dermatol. Rep. 2017, 6, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Whiteley, J.; Emir, B.; Seitzman, R.; Makinson, G. The Burden of Atopic Dermatitis in US Adults: Results from the 2013 National Health and Wellness Survey. Curr. Med. Res. Opin. 2016, 32, 1645–1651. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.H.; Silverberg, J.I. Association between Atopic Dermatitis and Depression in US Adults. J. Investig. Dermatol. 2015, 135, 3183–3186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaghmaie, P.; Koudelka, C.W.; Simpson, E.L. Mental Health Comorbidity in Patients with Atopic Dermatitis. J. Allergy Clin. Immunol. 2013, 131, 428–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimata, H. Prevalence of Suicidal Ideation in Patients with Atopic Dermatitis. Suicide Life Threat. Behav. 2006, 36, 120–124. [Google Scholar] [CrossRef]
- Zachariae, R.; Zachariae, C.; Ibsen, H.H.W.; Mortensen, J.T.; Wulf, H.C. Psychological Symptoms and Quality of Life of Dermatology Outpatients and Hospitalized Dermatology Patients. Acta Derm. Venereol. 2004, 84, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Camfferman, D.; Kennedy, J.D.; Gold, M.; Martin, A.J.; Lushington, K. Eczema and Sleep and Its Relationship to Daytime Functioning in Children. Sleep Med. Rev. 2010, 14, 359–369. [Google Scholar] [CrossRef]
- Wittkowski, A.; Richards, H.L.; Griffiths, C.E.M.; Main, C.J. The Impact of Psychological and Clinical Factors on Quality of Life in Individuals with Atopic Dermatitis. J. Psychosom. Res. 2004, 57, 195–200. [Google Scholar] [CrossRef]
- Yu, S.H.; Attarian, H.; Zee, P.; Silverberg, J.I. Burden of Sleep and Fatigue in US Adults With Atopic Dermatitis. Dermat. Contact Atopic Occup. Drug 2016, 27, 50–58. [Google Scholar] [CrossRef]
- Drucker, A.M.; Wang, A.R.; Li, W.Q.; Sevetson, E.; Block, J.K.; Qureshi, A.A. The Burden of Atopic Dermatitis: Summary of a Report for the National Eczema Association. J. Investig. Dermatol. 2017, 137, 26–30. [Google Scholar] [CrossRef] [Green Version]
- Sandhu, J.K.; Wu, K.K.; Bui, T.L.; Armstrong, A.W. Association between Atopic Dermatitis and Suicidality: A Systematic Review and Meta-Analysis. JAMA Dermatol. 2019, 155, 178–187. [Google Scholar] [CrossRef] [Green Version]
- Koo, J.; Marangell, L.B.; Nakamura, M.; Armstrong, A.; Jeon, C.; Bhutani, T.; Wu, J.J. Depression and Suicidality in Psoriasis: Review of the Literature Including the Cytokine Theory of Depression. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 1999–2009. [Google Scholar] [CrossRef] [PubMed]
- Davami, M.H.; Baharlou, R.; Vasmehjani, A.A.; Ghanizadeh, A.; Keshtkar, M.; Dezhkam, I.; Atashzar, M.R. Elevated IL-17 and TGF-β Serum Levels: A Positive Correlation between T-Helper 17 Cell-Related Pro-Inflammatory Responses with Major Depressive Disorder. Basic Clin. Neurosci. 2016, 7, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Lindqvist, D.; Janelidze, S.; Hagell, P.; Erhardt, S.; Samuelsson, M.; Minthon, L.; Hansson, O.; Björkqvist, M.; Träskman-Bendz, L.; Brundin, L. Interleukin-6 Is Elevated in the Cerebrospinal Fluid of Suicide Attempters and Related to Symptom Severity. Biol. Psychiatry 2009, 66, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Bieber, T. Atopic Dermatitis. N. Engl. J. Med. 2008, 358, 1483–1494. [Google Scholar] [CrossRef] [PubMed]
- Siegfried, E.C.; Hebert, A.A. Diagnosis of Atopic Dermatitis: Mimics, Overlaps, and Complications. J. Clin. Med. 2015, 4, 884–917. [Google Scholar] [CrossRef] [Green Version]
- Brunner, P.M.; Guttman-Yassky, E. Racial Differences in Atopic Dermatitis. Ann. Allergy. Asthma Immunol. 2019, 122, 449–455. [Google Scholar] [CrossRef] [Green Version]
- Yew, Y.W.; Thyssen, J.P.; Silverberg, J.I. A Systematic Review and Meta-Analysis of the Regional and Age-Related Differences in Atopic Dermatitis Clinical Characteristics. J. Am. Acad. Dermatol. 2019, 80, 390–401. [Google Scholar] [CrossRef]
- Bratu, D.; Boda, D.; Caruntu, C. Reflectance Confocal Microscopy in Monitoring Atopic Dermatitis Treated with Topical Calcineurin Inhibitors. Healthcare 2023, 11, 152. [Google Scholar] [CrossRef]
- Akan, A.; Dibek-Mısırlıoğlu, E.; Civelek, E.; Vezir, E.; Kocabaş, C.N. Diagnosis of Atopic Dermatitis in Children: Comparison of the Hanifin-Rajka and the United Kingdom Working Party Criteria. Allergol. Immunopathol. 2020, 48, 175–181. [Google Scholar] [CrossRef]
- Bieber, T. Atopic Dermatitis. Ann. Dermatol. 2010, 22, 125. [Google Scholar] [CrossRef] [Green Version]
- Adam, D.N.; Gooderham, M.J.; Beecker, J.R.; Hong, C.H.; Jack, C.S.; Jain, V.; Lansang, P.; Lynde, C.W.; Papp, K.A.; Prajapati, V.H.; et al. Expert Consensus on the Systemic Treatment of Atopic Dermatitis in Special Populations. J. Eur. Acad. Dermatol. Venereol. 2023, 37, 1135–1148. [Google Scholar] [CrossRef] [PubMed]
- Drucker, A.M.; Eyerich, K.; de Bruin-Weller, M.S.; Thyssen, J.P.; Spuls, P.I.; Irvine, A.D.; Girolomoni, G.; Dhar, S.; Flohr, C.; Murrell, D.F.; et al. Use of Systemic Corticosteroids for Atopic Dermatitis: International Eczema Council Consensus Statement. Br. J. Dermatol. 2018, 178, 768–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gooderham, M.J.; Hong, C.; Albrecht, L.; Bissonnette, R.; Dhadwal, G.; Gniadecki, R.; Grewal, P.; Kirchhof, M.G.; Landells, I.; Lansang, P.; et al. Approach to the Assessment and Management of Adult Patients With Atopic Dermatitis: A Consensus Document. J. Cutan. Med. Surg. 2018, 22, 3S–5S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiricozzi, A.; Maurelli, M.; Calabrese, L.; Peris, K.; Girolomoni, G. Overview of Atopic Dermatitis in Different Ethnic Groups. J. Clin. Med. 2023, 12, 2701. [Google Scholar] [CrossRef]
- Nedoszytko, B.; Reszka, E.; Gutowska-Owsiak, D.; Trzeciak, M.; Lange, M.; Jarczak, J.; Niedoszytko, M.; Jablonska, E.; Romantowski, J.; Strapagiel, D.; et al. Genetic and Epigenetic Aspects of Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 6484. [Google Scholar] [CrossRef]
- Kim, J.; Kim, B.E.; Leung, D.Y.M. Pathophysiology of Atopic Dermatitis: Clinical Implications. Allergy Asthma Proc. 2019, 40, 84–92. [Google Scholar] [CrossRef]
- Bjerre, R.D.; Bandier, J.; Skov, L.; Engstrand, L.; Johansen, J.D. The Role of the Skin Microbiome in Atopic Dermatitis: A Systematic Review. Br. J. Dermatol. 2017, 177, 1272–1278. [Google Scholar] [CrossRef] [Green Version]
- Kong, H.H.; Oh, J.; Deming, C.; Conlan, S.; Grice, E.A.; Beatson, M.A.; Nomicos, E.; Polley, E.C.; Komarow, H.D.; Mullikin, J.; et al. Temporal Shifts in the Skin Microbiome Associatedwith Disease Flares and Treatment in Childrenwith Atopic Dermatitis. Genome Res. 2012, 22, 850–859. [Google Scholar] [CrossRef] [Green Version]
- Edslev, S.M.; Agner, T.; Andersen, P.S. Skin Microbiome in Atopic Dermatitis. Acta Derm. Venereol. 2020, 100, 358–366. [Google Scholar] [CrossRef]
- Cabanillas, B.; Novak, N. Atopic Dermatitis and Filaggrin. Curr. Opin. Immunol. 2016, 42, 1–8. [Google Scholar] [CrossRef]
- Jakasa, I.; Koster, E.S.; Calkoen, F.; McLean, W.H.I.; Campbell, L.E.; Bos, J.D.; Verberk, M.M.; Kezic, S. Skin Barrier Function in Healthy Subjects and Patients with Atopic Dermatitis in Relation to Filaggrin Loss-of-Function Mutations. J. Investig. Dermatol. 2011, 131, 540–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidenari, S.; Giusti, G. Objective Assessment of the Skin of Children Affected by Atopic Dermatitis: A Study of PH, Capacitance and TEWL in Eczematous and Clinically Uninvolved Skin. Acta Derm. Venereol. 1995, 75, 429–433. [Google Scholar] [CrossRef] [PubMed]
- Kezic, S.; Kemperman, P.M.J.H.; Koster, E.S.; De Jongh, C.M.; Thio, H.B.; Campbell, L.E.; Irvine, A.D.; McLean, I.W.H.; Puppels, G.J.; Caspers, P.J. Loss-of-Function Mutations in the Filaggrin Gene Lead to Reduced Level of Natural Moisturizing Factor in the Stratum Corneum. J. Investig. Dermatol. 2008, 128, 2117–2119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Smeden, J.; Bouwstra, J.A. Stratum Corneum Lipids: Their Role for the Skin Barrier Function in Healthy Subjects and Atopic Dermatitis Patients. Curr. Probl. Dermatol. 2016, 49, 8–26. [Google Scholar] [CrossRef]
- Lee, S.Y.; Lee, E.; Park, Y.M.; Hong, S.J. Microbiome in the Gut-Skin Axis in Atopic Dermatitis. Allergy Asthma Immunol. Res. 2018, 10, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C.J. Host Interactions of Probiotic Bacterial Surface Molecules: Comparison with Commensals and Pathogens. Nat. Rev. Microbiol. 2010, 8, 171–184. [Google Scholar] [CrossRef]
- Ueno, N.; Fujiya, M.; Segawa, S.; Nata, T.; Moriichi, K.; Tanabe, H.; Mizukami, Y.; Kobayashi, N.; Ito, K.; Kohgo, Y. Heat-Killed Body of Lactobacillus Brevis SBC8803 Ameliorates Intestinal Injury in a Murine Model of Colitis by Enhancing the Intestinal Barrier Function. Inflamm. Bowel Dis. 2011, 17, 2235–2250. [Google Scholar] [CrossRef]
- Smits, H.H.; Engering, A.; Van Der Kleij, D.; De Jong, E.C.; Schipper, K.; Van Capel, T.M.M.; Zaat, B.A.J.; Yazdanbakhsh, M.; Wierenga, E.A.; Van Kooyk, Y.; et al. Selective Probiotic Bacteria Induce IL-10-Producing Regulatory T Cells in Vitro by Modulating Dendritic Cell Function through Dendritic Cell-Specific Intercellular Adhesion Molecule 3-Grabbing Nonintegrin. J. Allergy Clin. Immunol. 2005, 115, 1260–1267. [Google Scholar] [CrossRef]
- Iwase, T.; Uehara, Y.; Shinji, H.; Tajima, A.; Seo, H.; Takada, K.; Agata, T.; Mizunoe, Y. Staphylococcus Epidermidis Esp Inhibits Staphylococcus Aureus Biofilm Formation and Nasal Colonization. Nature 2010, 465, 346–349. [Google Scholar] [CrossRef]
- Tao, R.; Li, R.; Wang, R. Dysbiosis of Skin Mycobiome in Atopic Dermatitis. Mycoses 2022, 65, 285–293. [Google Scholar] [CrossRef]
- Løset, M.; Brown, S.J.; Saunes, M.; Hveem, K. Genetics of Atopic Dermatitis: From DNA Sequence to Clinical Relevance. Dermatology 2019, 235, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Esparza-Gordillo, J.; Weidinger, S.; Fölster-Holst, R.; Bauerfeind, A.; Ruschendorf, F.; Patone, G.; Rohde, K.; Marenholz, I.; Schulz, F.; Kerscher, T.; et al. A Common Variant on Chromosome 11q13 Is Associated with Atopic Dermatitis. Nat. Genet. 2009, 41, 596–601. [Google Scholar] [CrossRef]
- Paternoster, L.; Standl, M.; Waage, J.; Baurecht, H.; Hotze, M.; Strachan, D.P.; Curtin, J.A.; Bønnelykke, K.; Tian, C.; Takahashi, A.; et al. Multi-Ancestry Genome-Wide Association Study of 21,000 Cases and 95,000 Controls Identifies New Risk Loci for Atopic Dermatitis. Nat. Genet. 2015, 47, 1449–1456. [Google Scholar] [CrossRef] [PubMed]
- Rastegar-Mojarad, M.; Ye, Z.; Kolesar, J.M.; Hebbring, S.J.; Lin, S.M. Opportunities for Drug Repositioning from Phenome-Wide Association Studies. Nat. Biotechnol. 2015, 33, 342–345. [Google Scholar] [CrossRef]
- Taylan, F.; Nilsson, D.; Asad, S.; Lieden, A.; Wahlgren, C.-F.; Winge, M.C.G.; Bilcha, K.D.; Nordenskjöld, M.; Bradley, M. Whole-Exome Sequencing of Ethiopian Patients with Ichthyosis Vulgaris and Atopic Dermatitis. J. Allergy Clin. Immunol. 2015, 136, 507–509.e19. [Google Scholar] [CrossRef] [PubMed]
- Lawlor, D.A.; Harbord, R.M.; Sterne, J.A.C.; Timpson, N.; Davey Smith, G. Mendelian Randomization: Using Genes as Instruments for Making Causal Inferences in Epidemiology. Stat. Med. 2008, 27, 1133–1163. [Google Scholar] [CrossRef]
- Davey Smith, G.; Ebrahim, S. ‘Mendelian Randomization’: Can Genetic Epidemiology Contribute to Understanding Environmental Determinants of Disease? Int. J. Epidemiol. 2003, 32, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, M.D.; Holzinger, E.R.; Li, R.; Pendergrass, S.A.; Kim, D. Methods of Integrating Data to Uncover Genotype–Phenotype Interactions. Nat. Rev. Genet. 2015, 16, 85–97. [Google Scholar] [CrossRef]
- Zhang, A.; Silverberg, J.I. Association of Atopic Dermatitis with Being Overweight and Obese: A Systematic Review and Metaanalysis. J. Am. Acad. Dermatol. 2015, 72, 606–616.e4. [Google Scholar] [CrossRef]
- Silverberg, J.I.; Greenland, P. Eczema and Cardiovascular Risk Factors in 2 US Adult Population Studies. J. Allergy Clin. Immunol. 2015, 135, 721–728.e6. [Google Scholar] [CrossRef]
- Silverberg, J.I. Atopic Disease and Cardiovascular Risk Factors in US Children. J. Allergy Clin. Immunol. 2016, 137, 938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silverberg, J.I. Association between Adult Atopic Dermatitis, Cardiovascular Disease, and Increased Heart Attacks in Three Population-Based Studies. Allergy 2015, 70, 1300–1308. [Google Scholar] [CrossRef] [PubMed]
- Donath, M.Y.; Shoelson, S.E. Type 2 Diabetes as an Inflammatory Disease. Nat. Rev. Immunol. 2011, 11, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Poetker, D.M.; Reh, D.D. A Comprehensive Review of the Adverse Effects of Systemic Corticosteroids. Otolaryngol. Clin. N. Am. 2010, 43, 753–768. [Google Scholar] [CrossRef]
- Manousaki, D.; Paternoster, L.; Standl, M.; Moffatt, M.F.; Farrall, M.; Bouzigon, E.; Strachan, D.P.; Demenais, F.; Lathrop, M.; Cookson, W.O.C.M.; et al. Vitamin D Levels and Susceptibility to Asthma, Elevated Immunoglobulin E Levels, and Atopic Dermatitis: A Mendelian Randomization Study. PLoS Med. 2017, 14, e1002294. [Google Scholar] [CrossRef] [Green Version]
- Egawa, G.; Kabashima, K. Barrier Dysfunction in the Skin Allergy. Allergol. Int. 2018, 67, 3–11. [Google Scholar] [CrossRef]
- Weidinger, S.; Illig, T.; Baurecht, H.; Irvine, A.D.; Rodriguez, E.; Diaz-Lacava, A.; Klopp, N.; Wagenpfeil, S.; Zhao, Y.; Liao, H.; et al. Loss-of-Function Variations within the Filaggrin Gene Predispose for Atopic Dermatitis with Allergic Sensitizations. J. Allergy Clin. Immunol. 2006, 118, 214–219. [Google Scholar] [CrossRef]
- McLean, W.H.I. Filaggrin Failure—from Ichthyosis Vulgaris to Atopic Eczema and Beyond. Br. J. Dermatol. 2016, 175, 4–7. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.J.; Kroboth, K.; Sandilands, A.; Campbell, L.E.; Pohler, E.; Kezic, S.; Cordell, H.J.; McLean, W.H.I.; Irvine, A.D. Intragenic Copy Number Variation within Filaggrin Contributes to the Risk of Atopic Dermatitis with a Dose-Dependent Effect. J. Investig. Dermatol. 2012, 132, 98–104. [Google Scholar] [CrossRef] [Green Version]
- Irvine, A.D.; McLean, W.H.I.; Leung, D.Y.M. Filaggrin Mutations Associated with Skin and Allergic Diseases. N. Engl. J. Med. 2011, 365, 1315–1327. [Google Scholar] [CrossRef] [Green Version]
- Irvine, A.D.; McLean, W.H.I. Breaking the (Un)Sound Barrier: Filaggrin Is a Major Gene for Atopic Dermatitis. J. Investig. Dermatol. 2006, 126, 1200–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, G.; Hashimoto-Hachiya, A.; Kiyomatsu-Oda, M.; Takemura, M.; Ohno, F.; Ito, T.; Morino-Koga, S.; Mitoma, C.; Nakahara, T.; Uchi, H.; et al. Aryl Hydrocarbon Receptor Activation Restores Filaggrin Expression via OVOL1 in Atopic Dermatitis. Cell Death Dis. 2017, 8, e2931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Q.; Yang, H.; Liu, E.; Wang, H. P38/ERK MAPK Signaling Pathways Are Involved in the Regulation of Filaggrin and Involucrin by IL-17. Mol. Med. Rep. 2017, 16, 8863–8867. [Google Scholar] [CrossRef] [Green Version]
- Leonardi, S.; Cuppari, C.; Manti, S.; Filippelli, M.; Parisi, G.F.; Borgia, F.; Briuglia, S.; Cannavò, S.P.; Salpietro, A.; Arrigo, T.; et al. Serum Interleukin 17, Interleukin 23, and Interleukin 10 Values in Children with Atopic Eczema/Dermatitis Syndrome (AEDS): Association with Clinical Severity and Phenotype. Allergy Asthma Proc. 2015, 36, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Howell, M.D.; Kim, B.E.; Gao, P.; Grant, A.V.; Boguniewicz, M.; DeBenedetto, A.; Schneider, L.; Beck, L.A.; Barnes, K.C.; Leung, D.Y.M. Cytokine Modulation of Atopic Dermatitis Filaggrin Skin Expression. J. Allergy Clin. Immunol. 2009, 124, R7–R12. [Google Scholar] [CrossRef]
- Meinke, M.C.; Richter, H.; Kleemann, A.; Lademann, J.; Tscherch, K.; Rohn, S.; Schempp, C.M. Characterization of Atopic Skin and the Effect of a Hyperforin-Rich Cream by Laser Scanning Microscopy. J. Biomed. Opt. 2014, 20, 051013. [Google Scholar] [CrossRef]
- Nygaard, U.; van den Bogaard, E.H.; Niehues, H.; Hvid, M.; Deleuran, M.; Johansen, C.; Vestergaard, C. The “Alarmins” HMBG1 and IL-33 Downregulate Structural Skin Barrier Proteins and Impair Epidermal Growth. Acta Derm. Venereol. 2017, 97, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Cuppari, C.; Manti, S.; Salpietro, A.; Valenti, S.; Capizzi, A.; Arrigo, T.; Salpietro, C.; Leonardi, S. HMGB1 Levels in Children with Atopic Eczema/Dermatitis Syndrome (AEDS). Pediatr. Allergy Immunol. 2016, 27, 99–102. [Google Scholar] [CrossRef]
- Dharmage, S.C.; Lowe, A.J.; Matheson, M.C.; Burgess, J.A.; Allen, K.J.; Abramson, M.J. Atopic Dermatitis and the Atopic March Revisited. Allergy 2014, 69, 17–27. [Google Scholar] [CrossRef]
- Lowe, A.J.; Leung, D.Y.M.; Tang, M.L.K.; Su, J.C.; Allen, K.J. The Skin as a Target for Prevention of the Atopic March. Ann. Allergy, Asthma Immunol. 2018, 120, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Boguniewicz, M. Biologic Therapy for Atopic Dermatitis: Moving Beyond the Practice Parameter and Guidelines. J. Allergy Clin. Immunol. Pract. 2017, 5, 1477–1487. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, B.P.; Guttman-Yassky, E.; Alexis, A.F. Atopic Dermatitis in Diverse Racial and Ethnic Groups-Variations in Epidemiology, Genetics, Clinical Presentation and Treatment. Exp. Dermatol. 2018, 27, 340–357. [Google Scholar] [CrossRef] [Green Version]
- Bin, L.; Leung, D.Y.M. Genetic and Epigenetic Studies of Atopic Dermatitis. Allergy Asthma. Clin. Immunol. 2016, 12, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esaki, H.; Ewald, D.A.; Ungar, B.; Rozenblit, M.; Zheng, X.; Xu, H.; Estrada, Y.D.; Peng, X.; Mitsui, H.; Litman, T.; et al. Identification of Novel Immune and Barrier Genes in Atopic Dermatitis by Laser Capture Micro-Dissection. J. Allergy Clin. Immunol. 2015, 135, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.L.; Yen, J.J.Y.; Hsu, L.C.; Kuo, N.W.; Su, M.W.; Yang, M.F.; Hsiao, Y.P.; Wang, I.J.; Liu, F.T. Association of STAT6 Genetic Variants with Childhood Atopic Dermatitis in Taiwanese Population. J. Dermatol. Sci. 2015, 79, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.S.; Leung, D.Y.M.; Rafaels, N.M.; Boguniewicz, M.; Hand, T.; Gao, L.; Hata, T.R.; Schneider, L.C.; Hanifin, J.M.; Beaty, T.H.; et al. Genetic Variants InInterferon Regulatory Factor 2(IRF2) Are Associated with Atopic Dermatitis AndEczema Herpeticum. J. Investig. Dermatol. 2012, 132, 650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salpietro, C.; Rigoli, L.; Del Giudice, M.M.; Cuppari, C.; Di Bella, C.; Salpietro, A.; Maiello, N.; La Rosa, M.; Marseglia, G.L.; Leonardi, S.; et al. TLR2 and TLR4 Gene Polymorphisms and Atopic Dermatitis in Italian Children: A Multicenter Study. Int. J. Immunopathol. Pharmacol. 2011, 24, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, H.; Makino, Y.; Nagata, M.; Furuta, J.; Enomoto, H.; Hirota, T.; Tamari, M.; Noguchi, E. A Rare Variant in CYP27A1 and Its Association with Atopic Dermatitis with High Serum Total IgE. Allergy 2016, 71, 1486–1489. [Google Scholar] [CrossRef]
- Heine, G.; Hoefer, N.; Franke, A.; Nöthling, U.; Schumann, R.R.; Hamann, L.; Worm, M. Association of Vitamin D Receptor Gene Polymorphisms with Severe Atopic Dermatitis in Adults. Br. J. Dermatol. 2013, 168, 855–858. [Google Scholar] [CrossRef]
- Yang, G.; Seok, J.K.; Kang, H.C.; Cho, Y.-Y.; Lee, H.S.; Lee, J.Y. Skin Barrier Abnormalities and Immune Dysfunction in Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 2867. [Google Scholar] [CrossRef] [Green Version]
- Horimukai, K.; Morita, K.; Narita, M.; Kondo, M.; Kitazawa, H.; Nozaki, M.; Shigematsu, Y.; Yoshida, K.; Niizeki, H.; Motomura, K.; et al. Application of Moisturizer to Neonates Prevents Development of Atopic Dermatitis. J. Allergy Clin. Immunol. 2014, 134, 824–830.e6. [Google Scholar] [CrossRef] [Green Version]
- Tsakok, T.; Woolf, R.; Smith, C.H.; Weidinger, S.; Flohr, C. Atopic Dermatitis: The Skin Barrier and Beyond. Br. J. Dermatol. 2019, 180, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Simpson, E.L.; Chalmers, J.R.; Hanifin, J.M.; Thomas, K.S.; Cork, M.J.; McLean, W.H.I.; Brown, S.J.; Chen, Z.; Chen, Y.; Williams, H.C. Emollient Enhancement of the Skin Barrier from Birth Offers Effective Atopic Dermatitis Prevention. J. Allergy Clin. Immunol. 2014, 134, 818–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, M.J.; Estravís, M.; García-Sánchez, A.; Dávila, I.; Isidoro-García, M.; Sanz, C. Genetics and Epigenetics of Atopic Dermatitis: An Updated Systematic Review. Genes 2020, 11, 442. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Chang, C.; Lu, Q. The Genetics and Epigenetics of Atopic Dermatitis-Filaggrin and Other Polymorphisms. Clin. Rev. Allergy Immunol. 2016, 51, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Stemmler, S.; Hoffjan, S. Trying to Understand the Genetics of Atopic Dermatitis. Mol. Cell. Probes 2016, 30, 374–385. [Google Scholar] [CrossRef]
- Bonamonte, D.; Filoni, A.; Vestita, M.; Romita, P.; Foti, C.; Angelini, G. The Role of the Environmental Risk Factors in the Pathogenesis and Clinical Outcome of Atopic Dermatitis. Biomed Res. Int. 2019, 2019, 2450605. [Google Scholar] [CrossRef]
- Mu, Z.; Zhang, J. The Role of Genetics, the Environment, and Epigenetics in Atopic Dermatitis. Adv. Exp. Med. Biol. 2020, 1253, 107–140. [Google Scholar] [CrossRef]
- Leitch, C.S.; Natafji, E.; Yu, C.; Abdul-Ghaffar, S.; Madarasingha, N.; Venables, Z.C.; Chu, R.; Fitch, P.M.; Muinonen-Martin, A.J.; Campbell, L.E.; et al. Filaggrin-Null Mutations Are Associated with Increased Maturation Markers on Langerhans Cells. J. Allergy Clin. Immunol. 2016, 138, 482–490.e7. [Google Scholar] [CrossRef] [Green Version]
- Ziyab, A.H.; Karmaus, W.; Holloway, J.W.; Zhang, H.; Ewart, S.; Arshad, S.H. DNA Methylation of the Filaggrin Gene Adds to the Risk of Eczema Associated with Loss-of-Function Variants. J. Eur. Acad. Dermatol. Venereol. 2013, 27, e420–e423. [Google Scholar] [CrossRef] [Green Version]
- Harb, H.; Renz, H. Update on Epigenetics in Allergic Disease. J. Allergy Clin. Immunol. 2015, 135, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, B.E.; Meissner, A.; Lander, E.S. The Mammalian Epigenome. Cell 2007, 128, 669–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botchkarev, V.A.; Gdula, M.R.; Mardaryev, A.N.; Sharov, A.A.; Fessing, M.Y. Epigenetic Regulation of Gene Expression in Keratinocytes. J. Investig. Dermatol. 2012, 132, 2505–2521. [Google Scholar] [CrossRef] [Green Version]
- Makeyev, E.V.; Maniatis, T. Multilevel Regulation of Gene Expression by MicroRNAs. Science 2008, 319, 1789. [Google Scholar] [CrossRef] [Green Version]
- Krol, J.; Loedige, I.; Filipowicz, W. The Widespread Regulation of MicroRNA Biogenesis, Function and Decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef]
- Rebane, A.; Akdis, C.A. MicroRNAs: Essential Players in the Regulation of Inflammation. J. Allergy Clin. Immunol. 2013, 132, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Simpson, M.R.; Brede, G.; Johansen, J.; Johnsen, R.; Storrø, O.; Sætrom, P.; Øien, T. Human Breast Milk MiRNA, Maternal Probiotic Supplementation and Atopic Dermatitis in Offspring. PLoS ONE 2015, 10, e0143496. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Shin, C. Emerging Roles of DROSHA beyond Primary MicroRNA Processing. RNA Biol. 2018, 15, 186–193. [Google Scholar] [CrossRef] [Green Version]
- Sonkoly, E.; Ståhle, M.; Pivarcsi, A. MicroRNAs and Immunity: Novel Players in the Regulation of Normal Immune Function and Inflammation. Semin. Cancer Biol. 2008, 18, 131–140. [Google Scholar] [CrossRef]
- Yu, X.; Wang, M.; Li, L.; Zhang, L.; Chan, M.T.V.; Wu, W.K.K. MicroRNAs in Atopic Dermatitis: A Systematic Review. J. Cell. Mol. Med. 2020, 24, 5966–5972. [Google Scholar] [CrossRef]
- Li, H.M.; Xiao, Y.J.; Min, Z.S.; Tan, C. Identification and Interaction Analysis of Key Genes and MicroRNAs in Atopic Dermatitis by Bioinformatics Analysis. Clin. Exp. Dermatol. 2019, 44, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Meng, W.; Ye, S.; Zhang, X.; Mo, X.; Liu, J.; Chen, D.; Lin, Y. MicroRNA-146a as a Potential Regulator Involved in the Pathogenesis of Atopic Dermatitis. Mol. Med. Rep. 2019, 20, 4645–4653. [Google Scholar] [CrossRef] [Green Version]
- Sonkoly, E.; Wei, T.; Janson, P.C.J.; Sääf, A.; Lundeberg, L.; Tengvall-Linder, M.; Norstedt, G.; Alenius, H.; Homey, B.; Scheynius, A.; et al. MicroRNAs: Novel Regulators Involved in the Pathogenesis of Psoriasis? PLoS ONE 2007, 2, e610. [Google Scholar] [CrossRef] [Green Version]
- Sonkoly, E.; Ståhle, M.; Pivarcsi, A. MicroRNAs: Novel Regulators in Skin Inflammation. Clin. Exp. Dermatol. 2008, 33, 312–315. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.X.; Rothenberg, M.E. Diagnostic, Functional, and Therapeutic Roles of MicroRNA in Allergic Diseases. J. Allergy Clin. Immunol. 2013, 132, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonkoly, E.; Janson, P.; Majuri, M.-L.; Savinko, T.; Fyhrquist, N.; Eidsmo, L.; Xu, N.; Meisgen, F.; Wei, T.; Bradley, M.; et al. MiR-155 Is Overexpressed in Patients with Atopic Dermatitis and Modulates T-Cell Proliferative Responses by Targeting Cytotoxic T Lymphocyte–Associated Antigen 4. J. Allergy Clin. Immunol. 2010, 126, 581–589.e20. [Google Scholar] [CrossRef]
- Stevens, M.L.; Zhang, Z.; Johansson, E.; Ray, S.; Jagpal, A.; Ruff, B.P.; Kothari, A.; He, H.; Martin, L.J.; Ji, H.; et al. Disease-Associated KIF3A Variants Alter Gene Methylation and Expression Impacting Skin Barrier and Atopic Dermatitis Risk. Nat. Commun. 2020, 11, 4092. [Google Scholar] [CrossRef]
- Johansson, E.; Biagini Myers, J.M.; Martin, L.J.; He, H.; Pilipenko, V.; Mersha, T.; Weirauch, M.; Salomonis, N.; Ryan, P.; LeMasters, G.K.; et al. KIF3A Genetic Variation Is Associated with Pediatric Asthma in the Presence of Eczema Independent of Allergic Rhinitis. J. Allergy Clin. Immunol. 2017, 140, 595–598.e5. [Google Scholar] [CrossRef] [Green Version]
- Melnik, B.C.; John, S.; Schmitz, G. Milk: An Exosomal MicroRNA Transmitter Promoting Thymic Regulatory T Cell Maturation Preventing the Development of Atopy? J. Transl. Med. 2014, 12, 43. [Google Scholar] [CrossRef] [Green Version]
- Yao, R.; Ma, Y.-L.; Liang, W.; Li, H.-H.; Ma, Z.-J.; Yu, X.; Liao, Y.-H. MicroRNA-155 Modulates Treg and Th17 Cells Differentiation and Th17 Cell Function by Targeting SOCS1. PLoS ONE 2012, 7, e46082. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Xue, H.-B.; Wang, F.; Shu, C.-M.; Zhang, J.-H. MicroRNA-155 May Be Involved in the Pathogenesis of Atopic Dermatitis by Modulating the Differentiation and Function of T Helper Type 17 (Th17) Cells. Clin. Exp. Immunol. 2015, 181, 142–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liew, W.C.; Sundaram, G.M.; Quah, S.; Lum, G.G.; Tan, J.S.L.; Ramalingam, R.; Common, J.E.A.; Tang, M.B.Y.; Lane, E.B.; Thng, S.T.G.; et al. Belinostat Resolves Skin Barrier Defects in Atopic Dermatitis by Targeting the Dysregulated MiR-335:SOX6 Axis. J. Allergy Clin. Immunol. 2020, 146, 606–620.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagundes, B.O.; de Sousa, T.R.; Nascimento, A.; Fernandes, L.A.; Sgnotto, F.d.R.; Orfali, R.L.; Aoki, V.; Duarte, A.J. da S.; Sanabani, S.S.; Victor, J.R. IgG from Adult Atopic Dermatitis (AD) Patients Induces Nonatopic Neonatal Thymic Gamma–Delta T Cells (ΓδT) to Acquire IL-22/IL-17 Secretion Profile with Skin-Homing Properties and Epigenetic Implications Mediated by MiRNA. Int. J. Mol. Sci. 2022, 23, 6872. [Google Scholar] [CrossRef] [PubMed]
- White, G.P.; Watt, P.M.; Holt, B.J.; Holt, P.G. Differential Patterns of Methylation of the IFN-γ Promoter at CpG and Non-CpG Sites Underlie Differences in IFN-γ Gene Expression Between Human Neonatal and Adult CD45RO− T Cells. J. Immunol. 2002, 168, 2820–2827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaminuma, O.; Kitamura, F.; Miyatake, S.; Yamaoka, K.; Miyoshi, H.; Inokuma, S.; Tatsumi, H.; Nemoto, S.; Kitamura, N.; Mori, A.; et al. T-Box 21 Transcription Factor Is Responsible for Distorted TH2 Differentiation in Human Peripheral CD4+ T Cells. J. Allergy Clin. Immunol. 2009, 123, 813–823.e3. [Google Scholar] [CrossRef] [PubMed]
- Fujimaki, W.; Takahashi, N.; Ohnuma, K.; Nagatsu, M.; Kurosawa, H.; Yoshida, S.; Dang, N.H.; Uchiyama, T.; Morimoto, C. Comparative study of regulatory T cell function of human CD25+ CD4+ T cells from thymocytes, cord blood, and adult peripheral blood. Clin. Dev. Immunol. 2008, 2008, 305859. [Google Scholar] [CrossRef] [Green Version]
- Schaub, B.; Liu, J.; Schleich, I.; Höppler, S.; Sattler, C.; von Mutius, E. Impairment of T Helper and T Regulatory Cell Responses at Birth. Allergy 2008, 63, 1438–1447. [Google Scholar] [CrossRef]
- Breton, C.V.; Byun, H.-M.; Wenten, M.; Pan, F.; Yang, A.; Gilliland, F.D. Prenatal Tobacco Smoke Exposure Affects Global and Gene-Specific DNA Methylation. Am. J. Respir. Crit. Care Med. 2009, 180, 462–467. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Levin, L.; Talaska, G.; Cheung, Y.Y.; Herbstman, J.; Tang, D.; Miller, R.L.; Perera, F.; Ho, S.-M. Maternal Exposure to Polycyclic Aromatic Hydrocarbons and 5′-CpG Methylation of Interferon-γ in Cord White Blood Cells. Environ. Health Perspect. 2012, 120, 1195–1200. [Google Scholar] [CrossRef] [Green Version]
- Hinz, D.; Bauer, M.; Röder, S.; Olek, S.; Huehn, J.; Sack, U.; Borte, M.; Simon, J.C.; Lehmann, I.; Herberth, G. Cord Blood Tregs with Stable FOXP3 Expression Are Influenced by Prenatal Environment and Associated with Atopic Dermatitis at the Age of One Year. Allergy 2012, 67, 380–389. [Google Scholar] [CrossRef]
- Herberth, G.; Bauer, M.; Gasch, M.; Hinz, D.; Röder, S.; Olek, S.; Kohajda, T.; Rolle-Kampczyk, U.; von Bergen, M.; Sack, U.; et al. Maternal and Cord Blood MiR-223 Expression Associates with Prenatal Tobacco Smoke Exposure and Low Regulatory T-Cell Numbers. J. Allergy Clin. Immunol. 2014, 133, 543–550.e4. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Feehley, T.J.; Nagler, C.R. The Role of Commensal Bacteria in the Regulation of Sensitization to Food Allergens. FEBS Lett. 2014, 588, 4258–4266. [Google Scholar] [CrossRef] [Green Version]
- Amberbir, A.; Medhin, G.; Erku, W.; Alem, A.; Simms, R.; Robinson, K.; Fogarty, A.; Britton, J.; Venn, A.; Davey, G. Effects of Helicobacter Pylori, Geohelminth Infection and Selected Commensal Bacteria on the Risk of Allergic Disease and Sensitization in 3-Year-Old Ethiopian Children. Clin. Exp. Allergy 2011, 41, 1422–1430. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Li, Y.; Stoll, M.L.; Tollefsbol, T.O. The Epigenetic Connection Between the Gut Microbiome in Obesity and Diabetes. Front. Genet. 2020, 10, 1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaub, B.; Liu, J.; Höppler, S.; Schleich, I.; Huehn, J.; Olek, S.; Wieczorek, G.; Illi, S.; von Mutius, E. Maternal Farm Exposure Modulates Neonatal Immune Mechanisms through Regulatory T Cells. J. Allergy Clin. Immunol. 2009, 123, 774–782.e5. [Google Scholar] [CrossRef]
- Lluis, A.; Depner, M.; Gaugler, B.; Saas, P.; Casaca, V.I.; Raedler, D.; Michel, S.; Tost, J.; Liu, J.; Genuneit, J.; et al. Increased Regulatory T-Cell Numbers Are Associated with Farm Milk Exposure and Lower Atopic Sensitization and Asthma in Childhood. J. Allergy Clin. Immunol. 2014, 133, 551–559.e10. [Google Scholar] [CrossRef]
- Michel, S.; Busato, F.; Genuneit, J.; Pekkanen, J.; Dalphin, J.-C.; Riedler, J.; Mazaleyrat, N.; Weber, J.; Karvonen, A.M.; Hirvonen, M.-R.; et al. Farm Exposure and Time Trends in Early Childhood May Influence DNA Methylation in Genes Related to Asthma and Allergy. Allergy 2013, 68, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Schieck, M.; Sharma, V.; Michel, S.; Toncheva, A.A.; Worth, L.; Potaczek, D.P.; Genuneit, J.; Kretschmer, A.; Depner, M.; Dalphin, J.-C.; et al. A Polymorphism in the TH2 Locus Control Region Is Associated with Changes in DNA Methylation and Gene Expression. Allergy 2014, 69, 1171–1180. [Google Scholar] [CrossRef]
- Brand, S.; Teich, R.; Dicke, T.; Harb, H.; Yildirim, A.Ö.; Tost, J.; Schneider-Stock, R.; Waterland, R.A.; Bauer, U.-M.; von Mutius, E.; et al. Epigenetic Regulation in Murine Offspring as a Novel Mechanism for Transmaternal Asthma Protection Induced by Microbes. J. Allergy Clin. Immunol. 2011, 128, 618–625.e7. [Google Scholar] [CrossRef]
- Cole, C.; Kroboth, K.; Schurch, N.J.; Sandilands, A.; Sherstnev, A.; O’Regan, G.M.; Watson, R.M.; Irwin McLean, W.H.; Barton, G.J.; Irvine, A.D.; et al. Filaggrin-Stratified Transcriptomic Analysis of Pediatric Skin Identifies Mechanistic Pathways in Patients with Atopic Dermatitis. J. Allergy Clin. Immunol. 2014, 134, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Goleva, E.; Calatroni, A.; Lebeau, P.; Berdyshev, E.; Taylor, P.; Kreimer, S.; Cole, R.N.; Leung, D.Y.M. Skin tape proteomics identifies pathways associated with transepidermal water loss and allergen polysensitization in atopic dermatitis. J. Allergy Clin. Immunol. 2021, 146, 1367–1378. [Google Scholar] [CrossRef]
- Simpson, E.L.; Villarreal, M.; Jepson, B.; Rafaels, N.; David, G.; Hanifin, J.; Taylor, P.; Boguniewicz, M.; Yoshida, T.; De Benedetto, A.; et al. Patients with Atopic Dermatitis Colonized with Staphylococcus Aureus Have a Distinct Phenotype and Endotype. J. Investig. Dermatol. 2018, 138, 2224–2233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamlin, S.L.; Kao, J.; Frieden, I.J.; Sheu, M.Y.; Fowler, A.J.; Fluhr, J.W.; Williams, M.L.; Elias, P.M. Ceramide-Dominant Barrier Repair Lipids Alleviate Childhood Atopic Dermatitis: Changes in Barrier Function Provide a Sensitive Indicator of Disease Activity. J. Am. Acad. Dermatol. 2002, 47, 198–208. [Google Scholar] [CrossRef]
- Flohr, C.; Perkin, M.; Logan, K.; Marrs, T.; Radulovic, S.; Campbell, L.E.; MacCallum, S.F.; Irwin McLean, W.H.; Lack, G. Atopic Dermatitis and Disease Severity Are the Main Risk Factors for Food Sensitization in Exclusively Breastfed Infants. J. Investig. Dermatol. 2014, 134, 345–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, H.; Brown, S.; Danby, S.; Flohr, C. Research Techniques Made Simple: Transepidermal Water Loss Measurement as a Research Tool. J. Investig. Dermatol. 2018, 138, 2295–2300.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zouboulis, C.C.; Elewa, R.; Ottaviani, M.; Fluhr, J.; Picardo, M.; Bernois, A.; Heusèle, C.; Camera, E. Age Influences the Skin Reaction Pattern to Mechanical Stress and Its Repair Level through Skin Care Products. Mech. Ageing Dev. 2018, 170, 98–105. [Google Scholar] [CrossRef]
- Kottner, J.; Lichterfeld, A.; Blume-Peytavi, U. Transepidermal Water Loss in Young and Aged Healthy Humans: A Systematic Review and Meta-Analysis. Arch. Dermatol. Res. 2013, 305, 315–323. [Google Scholar] [CrossRef]
- Yu, W.; Freeland, D.M.H.; Nadeau, K.C. Food Allergy: Immune Mechanisms, Diagnosis and Immunotherapy. Nat. Rev. Immunol. 2016, 16, 751–765. [Google Scholar] [CrossRef]
- Schjødt, M.S.; Gürdeniz, G.; Chawes, B. The Metabolomics of Childhood Atopic Diseases: A Comprehensive Pathway-Specific Review. Metabolites 2020, 10, 511. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, G.; Liu, X.; Shao, Y.; Gao, P.; Xin, C.; Cui, Z.; Zhao, X.; Xu, G. Serum Metabolomics Study and Eicosanoid Analysis of Childhood Atopic Dermatitis Based on Liquid Chromatography–Mass Spectrometry. J. Proteome Res. 2014, 13, 5715–5723. [Google Scholar] [CrossRef]
- Crestani, E.; Harb, H.; Charbonnier, L.-M.; Leirer, J.; Motsinger-Reif, A.; Rachid, R.; Phipatanakul, W.; Kaddurah-Daouk, R.; Chatila, T.A. Untargeted Metabolomic Profiling Identifies Disease-Specific Signatures in Food Allergy and Asthma. J. Allergy Clin. Immunol. 2020, 145, 897–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turi, K.N.; Romick-Rosendale, L.; Gebretsadik, T.; Watanabe, M.; Brunwasser, S.; Anderson, L.J.; Moore, M.L.; Larkin, E.K.; Peebles, R.S.; Hartert, T.V. Using Urine Metabolomics to Understand the Pathogenesis of Infant Respiratory Syncytial Virus (RSV) Infection and Its Role in Childhood Wheezing. Metabolomics 2018, 14, 135. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| MiRNA Species | AD | References |
| MiR-155 | - overexpressed - essential for the differentiation of Th17 and Treg - associated with sustained inflammation, severity of the disease, and the percentage of Th17 | [58,106,117,123,129,136,137,138,139,142,143,144] |
| Let-7-a-d-family | - downregulated - overproduction of IL-13 and CCR17 - predominance of Th2 responses | [58,129,138] |
| MiR-375 | - similar with Let-7-a-d-family - induces the synthesis of TSLP - blocks the expression of KLF5 | [58,129,138] |
| MiR-151a | - regulates the beta2 receptor of IL-12 - increased - biomarker AD | [58,143] |
| hsa-mir-144-3p | - biomarker AD - umbilical cord fluid | [58,143] |
| MiR-29b | - positive correlation with SCORAD | [133] |
| MiR-143 | - inhibits IL-13 (when IL-13 is expressed, it downregulates filaggrin, involucrin and loricrin) | [133] |
| MiR-335 and SOX work codependently | - miR-335 is suppressed, while SOX is upregulated | [145] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bratu, D.; Boda, D.; Caruntu, C. Genomic, Epigenomic, Transcriptomic, Proteomic and Metabolomic Approaches in Atopic Dermatitis. Curr. Issues Mol. Biol. 2023, 45, 5215-5231. https://doi.org/10.3390/cimb45060331
Bratu D, Boda D, Caruntu C. Genomic, Epigenomic, Transcriptomic, Proteomic and Metabolomic Approaches in Atopic Dermatitis. Current Issues in Molecular Biology. 2023; 45(6):5215-5231. https://doi.org/10.3390/cimb45060331
Chicago/Turabian StyleBratu, Dalia, Daniel Boda, and Constantin Caruntu. 2023. "Genomic, Epigenomic, Transcriptomic, Proteomic and Metabolomic Approaches in Atopic Dermatitis" Current Issues in Molecular Biology 45, no. 6: 5215-5231. https://doi.org/10.3390/cimb45060331