
The Anti-Endometriotic Effect of Cyperi Rhizoma Extract, Inhibiting Cell Adhesion and the Expression of Pain-Related Factors through Akt and NF-kB Pathways

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Preparation of the Extract
2.2. Materials
2.3. Cell Culture
2.4. Transfection
2.5. Cell Viability Assay
2.6. Adhesion Assay
2.7. Nuclear Extraction
2.8. Western Blot Analysis
2.9. RNA Isolation and Real-Time RT–PCR Analysis
2.10. Statistical Analysis
3. Results
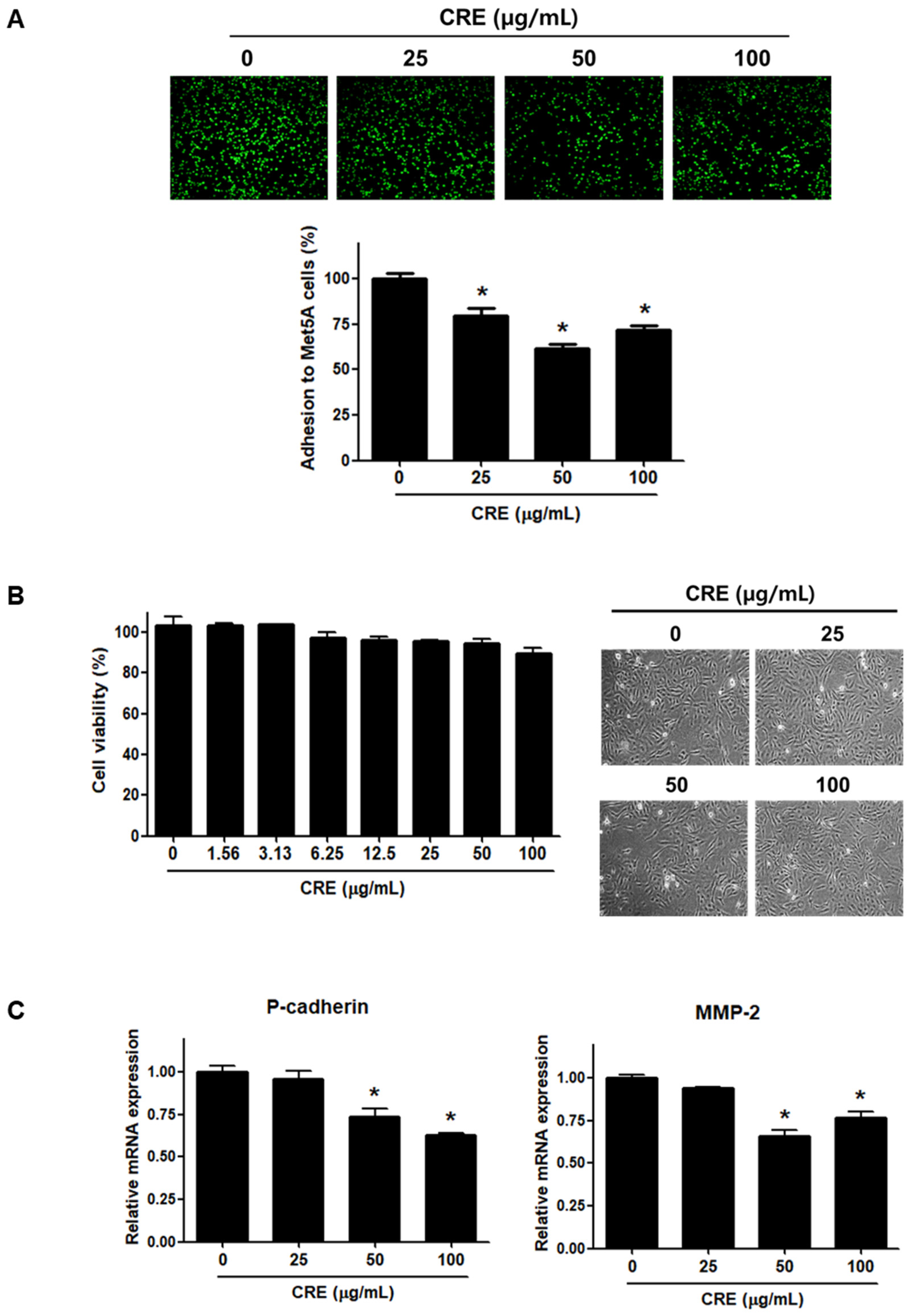
3.1. Cyperi Rhizoma Extract Inhibits Endometriotic Cell Adhesion to Mesothelial Cells
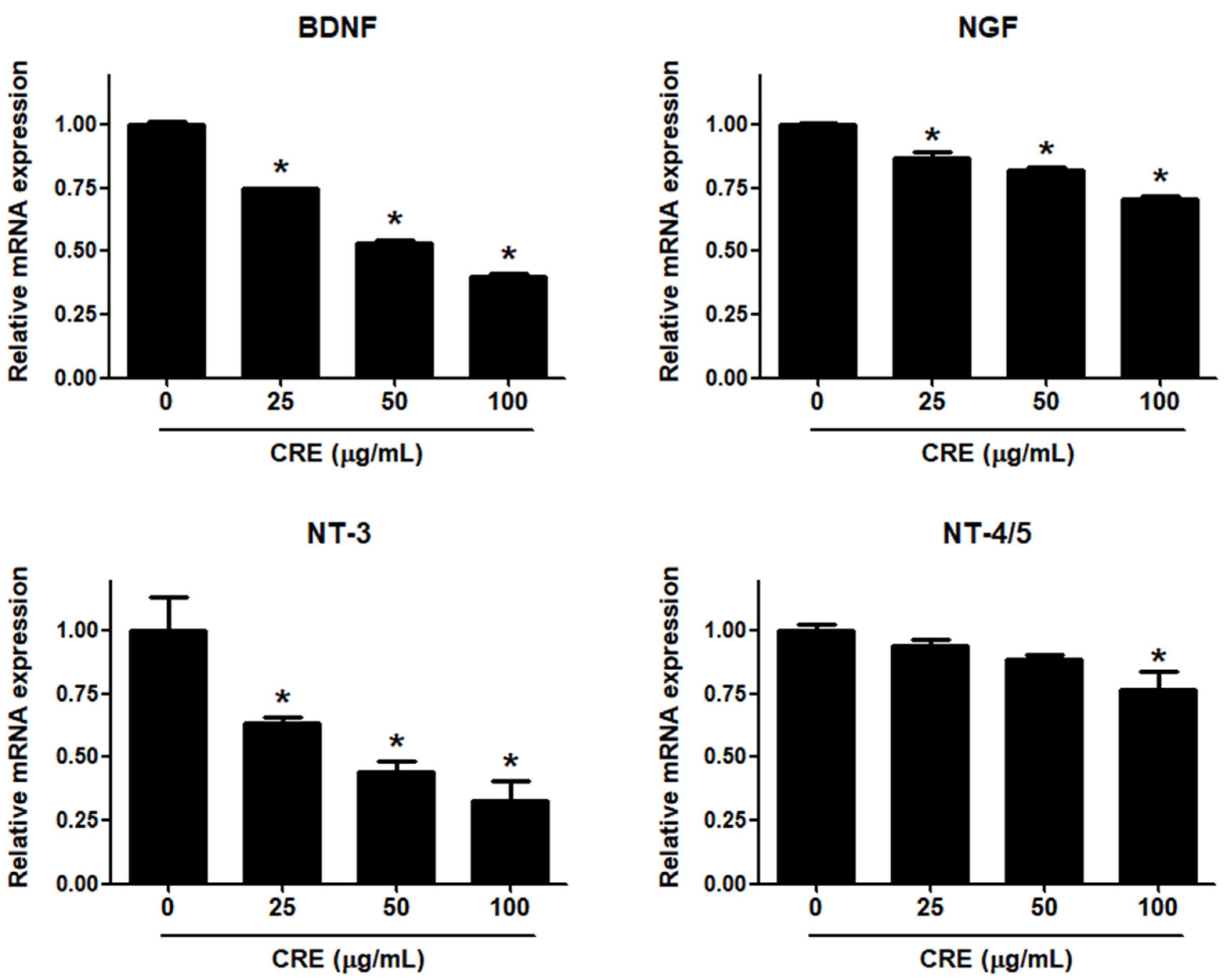
3.2. Cyperi Rhizoma Extract Decreases the Expression of Neurotrophins in Human Endometriotic Cells
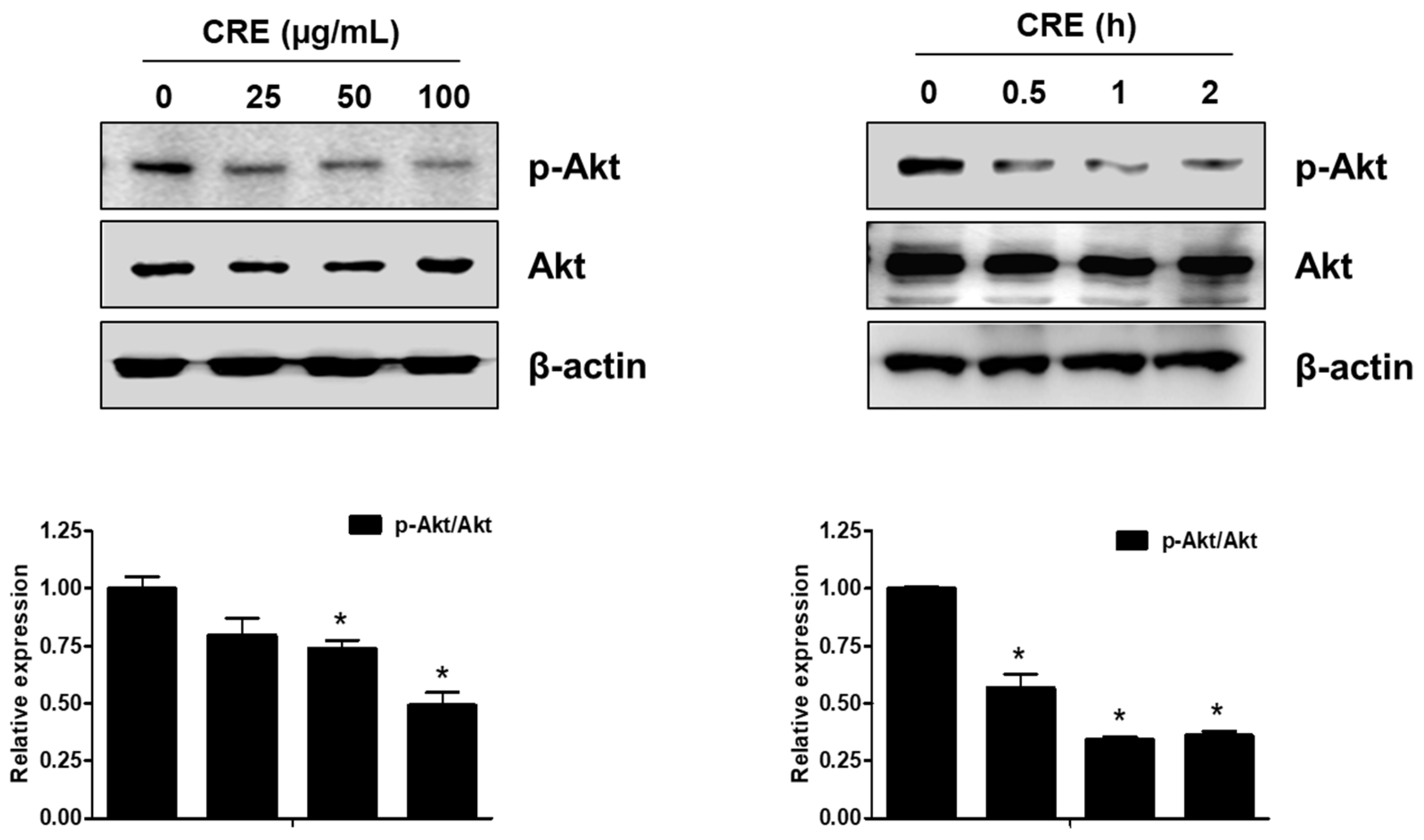
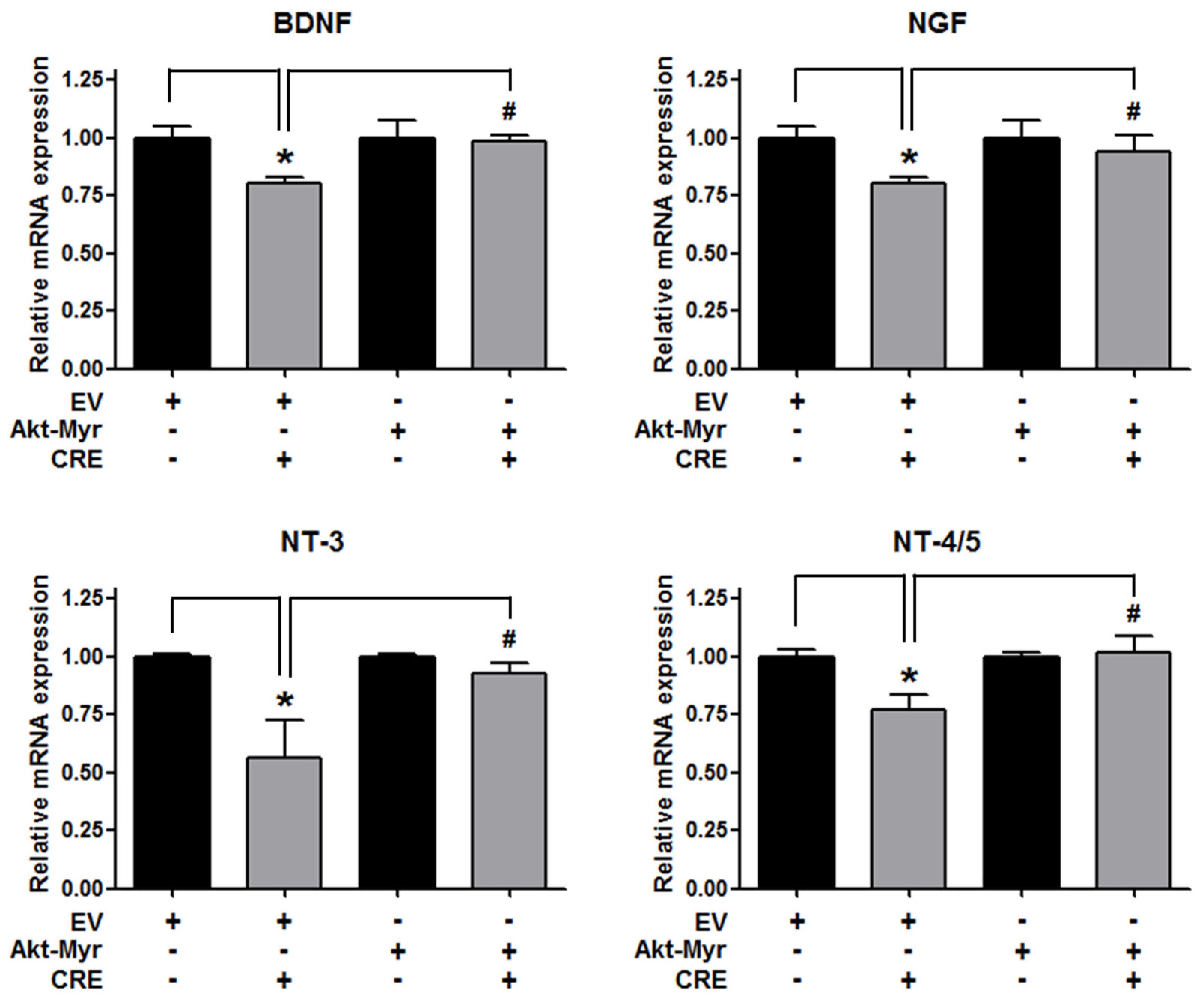
3.3. The Akt Pathway Is Involved in Cyperi Rhizoma Extract-Induced Anti-Endometriotic Effects in Human Endometriotic Cells
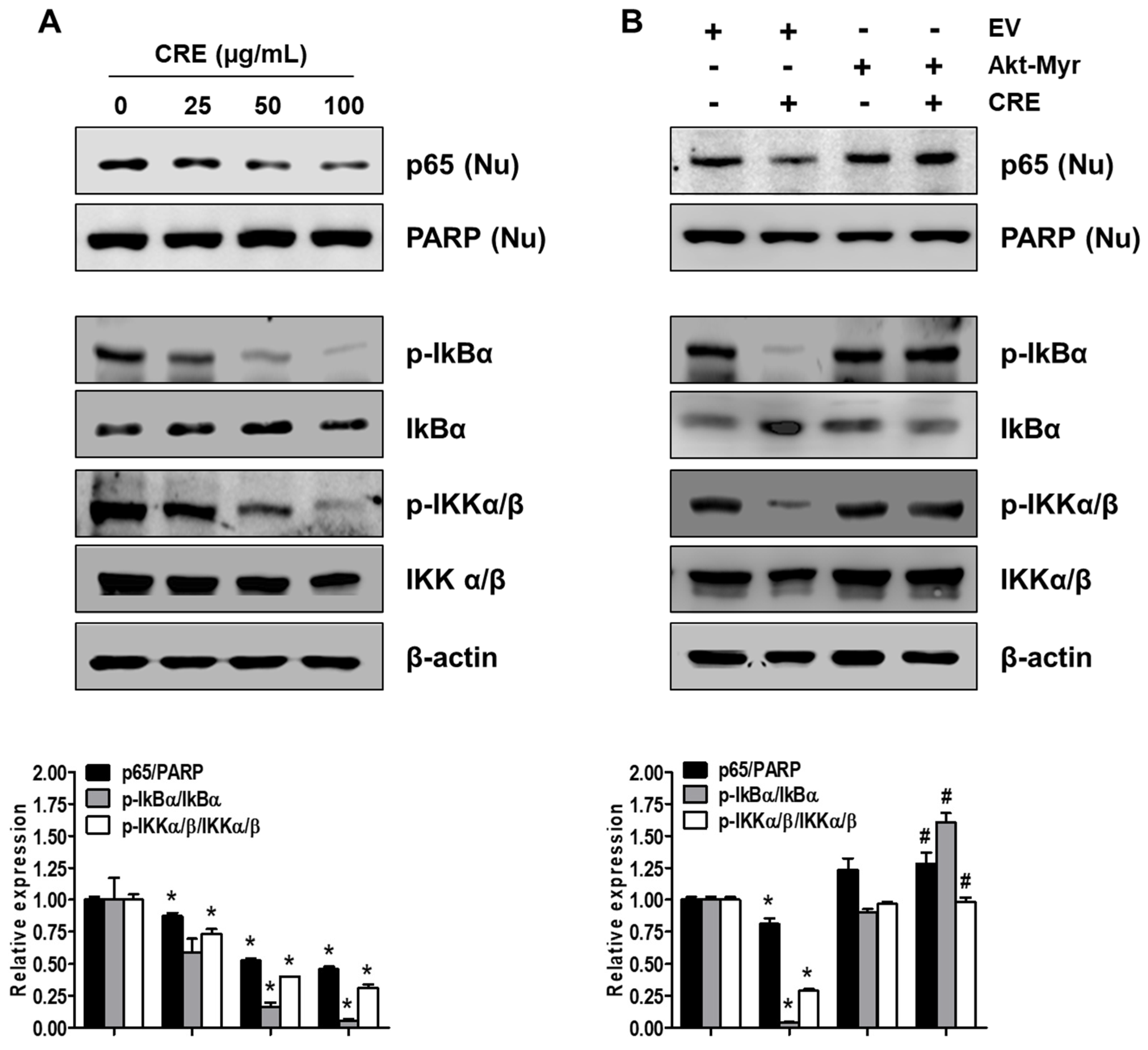
3.4. Cyperi Rhizoma Extract Inhibits NF-kB Activation through the Akt Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Burney, R.O.; Giudice, L.C. Pathogenesis and pathophysiology of endometriosis. Fertil. Steril. 2012, 98, 511–519. [Google Scholar] [CrossRef] [Green Version]
- Giudice, L.C.; Kao, L.C. Endometriosis. Lancet 2004, 364, 1789–1799. [Google Scholar] [CrossRef]
- Czyzyk, A.; Podfigurna, A.; Szeliga, A.; Meczekalski, B. Update on endometriosis pathogenesis. Minerva Ginecol. 2017, 69, 447–461. [Google Scholar] [CrossRef] [PubMed]
- Andres, M.P.; Arcoverde, F.V.L.; Souza, C.C.C.; Fernandes, L.F.C.; Abrao, M.S.; Kho, R.M. Extrapelvic Endometriosis: A Systematic Review. J. Minim. Invasive Gynecol. 2020, 27, 373–389. [Google Scholar] [CrossRef] [Green Version]
- Young, V.J.; Brown, J.K.; Saunders, P.T.; Horne, A.W. The role of the peritoneum in the pathogenesis of endometriosis. Hum. Reprod. Update 2013, 19, 558–569. [Google Scholar] [CrossRef]
- Chen, G.T.; Tai, C.T.; Yeh, L.S.; Yang, T.C.; Tsai, H.D. Identification of the cadherin subtypes present in the human peritoneum and endometriotic lesions: Potential role for P-cadherin in the development of endometriosis. Mol. Reprod. Dev. 2002, 62, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Yang, Y.I.; Ahn, J.H.; Lee, J.G.; Lee, K.T.; Choi, J.H. Deer (Cervus elaphus) antler extract suppresses adhesion and migration of endometriotic cells and regulates MMP-2 and MMP-9 expression. J. Ethnopharmacol. 2012, 140, 391–397. [Google Scholar] [CrossRef]
- Morotti, M.; Vincent, K.; Becker, C.M. Mechanisms of pain in endometriosis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2017, 209, 8–13. [Google Scholar] [CrossRef]
- Pezet, S.; McMahon, S.B. Neurotrophins: Mediators and modulators of pain. Annu. Rev. Neurosci. 2006, 29, 507–538. [Google Scholar] [CrossRef]
- Wessels, J.M.; Kay, V.R.; Leyland, N.A.; Agarwal, S.K.; Foster, W.G. Assessing brain-derived neurotrophic factor as a novel clinical marker of endometriosis. Fertil. Steril. 2016, 105, 119–128.e5. [Google Scholar] [CrossRef] [Green Version]
- Streiter, S.; Fisch, B.; Sabbah, B.; Ao, A.; Abir, R. The importance of neuronal growth factors in the ovary. Mol. Hum. Reprod. 2016, 22, 3–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uddin, S.J.; Mondal, K.; Shilpi, J.A.; Rahman, M.T. Antidiarrhoeal activity of Cyperus rotundus. Fitoterapia 2006, 77, 134–136. [Google Scholar] [CrossRef] [PubMed]
- Dang, G.K.; Parekar, R.R.; Kamat, S.K.; Scindia, A.M.; Rege, N.N. Antiinflammatory activity of Phyllanthus emblica, Plumbago zeylanica and Cyperus rotundus in acute models of inflammation. Phytother. Res. 2011, 25, 904–908. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Qu, F. Treating gynaecological disorders with traditional Chinese medicine: A review. Afr. J. Tradit. Complement. Altern. Med. 2009, 6, 494–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, M.B.; Palit, T.K.; Singh, N.; Bhargava, K.P. Pharmacological studies to isolate the active constituents from Cyperus rotundus possessing anti-inflammatory, anti-pyretic and analgesic activities. Indian J. Med. Res. 1971, 59, 76–82. [Google Scholar]
- Singh, P.; Khosa, R.L.; Mishra, G.; Jha, K.K. Antidiabetic activity of ethanolic extract of Cyperus rotundus rhizomes in streptozotocin-induced diabetic mice. J. Pharm. Bioallied Sci. 2015, 7, 289–292. [Google Scholar] [CrossRef]
- Seo, W.G.; Pae, H.O.; Oh, G.S.; Chai, K.Y.; Kwon, T.O.; Yun, Y.G.; Kim, N.Y.; Chung, H.T. Inhibitory effects of methanol extract of Cyperus rotundus rhizomes on nitric oxide and superoxide productions by murine macrophage cell line, RAW 264.7 cells. J. Ethnopharmacol. 2001, 76, 59–64. [Google Scholar] [CrossRef]
- Ahn, J.H.; Lee, T.W.; Kim, K.H.; Byun, H.; Ryu, B.; Lee, K.T.; Jang, D.S.; Choi, J.H. 6-Acetoxy Cyperene, a Patchoulane-type Sesquiterpene Isolated from Cyperus rotundus Rhizomes Induces Caspase-dependent Apoptosis in Human Ovarian Cancer Cells. Phytother. Res. 2015, 29, 1330–1338. [Google Scholar] [CrossRef]
- Zhu, M.; Luk, H.H.; Fung, H.S.; Luk, C.T. Cytoprotective effects of Cyperus rotundus against ethanol induced gastric ulceration in rats. Phytother. Res. 1997, 11, 392–394. [Google Scholar] [CrossRef]
- Kamala, A.; Middha, S.K.; Gopinath, C.; Sindhura, H.S.; Karigar, C.S. In vitro Antioxidant Potentials of Cyperus rotundus L. Rhizome Extracts and Their Phytochemical Analysis. Pharmacogn. Mag. 2018, 14, 261–267. [Google Scholar] [CrossRef]
- Mannarreddy, P.; Denis, M.; Munireddy, D.; Pandurangan, R.; Thangavelu, K.P.; Venkatesan, K. Cytotoxic effect of Cyperus rotundus rhizome extract on human cancer cell lines. Biomed. Pharmacother. 2017, 95, 1375–1387. [Google Scholar] [CrossRef]
- Jia, H.; Liu, Y.; Yu, M.; Shang, H.; Zhang, H.; Ma, L.; Zhang, T.; Zou, Z. Neuroprotective Effect of Cyperi rhizome against Corticosterone-Induced PC12 Cell Injury via Suppression of Ca2+ Overloading. Metabolites 2019, 9, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Yan, Y.; Liu, Z.; Wang, Y. Inflammation and endometriosis. Front. Biosci. 2016, 21, 941–948. [Google Scholar]
- Saunders, P.T.K.; Horne, A.W. Endometriosis: Etiology, pathobiology, and therapeutic prospects. Cell 2021, 184, 2807–2824. [Google Scholar] [CrossRef]
- Zeitvogel, A.; Baumann, R.; Starzinski-Powitz, A. Identification of an invasive, N-cadherin-expressing epithelial cell type in endometriosis using a new cell culture model. Am. J. Pathol. 2001, 159, 1839–1852. [Google Scholar] [CrossRef] [Green Version]
- Vercellini, P.; Vigano, P.; Somigliana, E.; Fedele, L. Endometriosis: Pathogenesis and treatment. Nat. Rev. Endocrinol. 2014, 10, 261–275. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Yamada, Y.; Morioka, S.; Niiro, E.; Shigemitsu, A.; Ito, F. Mechanism of pain generation for endometriosis-associated pelvic pain. Arch. Gynecol. Obstet. 2014, 289, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Fabi, F.; Grenier, K.; Parent, S.; Adam, P.; Tardif, L.; Leblanc, V.; Asselin, E. Regulation of the PI3K/Akt pathway during decidualization of endometrial stromal cells. PLoS ONE 2017, 12, e0177387. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Ramos, R.; Van Langendonckt, A.; Defrere, S.; Lousse, J.C.; Colette, S.; Devoto, L.; Donnez, J. Involvement of the nuclear factor-kappaB pathway in the pathogenesis of endometriosis. Fertil. Steril. 2010, 94, 1985–1994. [Google Scholar] [CrossRef] [PubMed]
- Hung, Y.C.; Kao, C.W.; Lin, C.C.; Liao, Y.N.; Wu, B.Y.; Hung, I.L.; Hu, W.L. Chinese Herbal Products for Female Infertility in Taiwan: A Population-Based Cohort Study. Medicine 2016, 95, e3075. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, N.K. Folk herbal remedies for diarrhoea and dysentery in central Nepal. Fitoterapia 1993, 64, 243–250. [Google Scholar] [CrossRef]
- Choi, H.J.; Chung, T.W.; Park, M.J.; Jung, Y.S.; Lee, S.O.; Kim, K.J.; Ha, K.T. Water-extracted tubers of Cyperus rotundus L. enhance endometrial receptivity through leukemia inhibitory factor-mediated expression of integrin αVβ3 and αVβ5. J. Ethnopharmacol. 2017, 208, 16–23. [Google Scholar] [CrossRef]
- Su, S.Y.; Muo, C.H.; Sung, F.C.; Morisky, D.E. Reduction of surgery rate in endometriosis patients who take Chinese medicine: A population-based retrospective cohort study. Complement. Ther. Med. 2014, 22, 632–639. [Google Scholar] [CrossRef]
- Kim, H.W.; Yoo, J.E. Inhibitory effect of traditional Korean medicine on the recurrent endometriosis after laparoscopic excision: A case report. Integr. Med. Res. 2018, 7, 296–301. [Google Scholar] [CrossRef]
- Xu, X.T.; Deng, Z.P.; Zhong, H.; Yao, Q.Q. Research progress on chemical constituents and pharmacological activities of Cyperus rotundus L. Qilu Pharm. Aff. 2012, 31, 473–475. [Google Scholar]
- Gamal, M.A.; Hani, K.M.K.; Sabrin, I.R.M. A Review: Compounds Isolated from Cyperus Species (Part II):Terpenoidal. Int. J. Pharmacogn. Phytochem. Res. 2015, 7, 83–99. [Google Scholar]
- Seo, Y.J.; Jeong, M.; Lee, K.T.; Jang, D.S.; Choi, J.H. Isocyperol, isolated from the rhizomes of Cyperus rotundus, inhibits LPS-induced inflammatory responses via suppression of the NF-kappaB and STAT3 pathways and ROS stress in LPS-stimulated RAW 264.7 cells. Int. Immunopharmacol. 2016, 38, 61–69. [Google Scholar] [CrossRef]
- Jung, S.H.; Kim, S.J.; Jun, B.G.; Lee, K.T.; Hong, S.P.; Oh, M.S.; Jang, D.S.; Choi, J.H. alpha-Cyperone, isolated from the rhizomes of Cyperus rotundus, inhibits LPS-induced COX-2 expression and PGE2 production through the negative regulation of NFkappaB signalling in RAW 264.7 cells. J. Ethnopharmacol. 2013, 147, 208–214. [Google Scholar] [CrossRef]
- Luo, M.; Qiu, J.; Zhang, Y.; Wang, J.; Dong, J.; Li, H.; Leng, B.; Zhang, Q.; Dai, X.; Niu, X.; et al. alpha-cyperone alleviates lung cell injury caused by Staphylococcus aureus via attenuation of alpha-hemolysin expression. J. Microbiol. Biotechnol. 2012, 22, 1170–1176. [Google Scholar] [CrossRef]
- Singh, N.; Kulshrestha, V.K.; Gupta, M.B.; Bhargava, K.P. Apharmacological study of Cyperus rotundus. Indian J. Med. Res. 1970, 58, 103–109. [Google Scholar]
- Jin, J.H.; Lee, D.U.; Kim, Y.S.; Kim, H.P. Anti-allergic activity of sesquiterpenes from the rhizomes of Cyperus rotundus. Arch. Pharm. Res. 2011, 34, 223–228. [Google Scholar] [CrossRef]
- Xia, B.; Tong, Y.; Xia, C.; Chen, C.; Shan, X. alpha-Cyperone Confers Antidepressant-Like Effects in Mice via Neuroplasticity Enhancement by SIRT3/ROS Mediated NLRP3 Inflammasome Deactivation. Front. Pharmacol. 2020, 11, 577062. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Lv, S.; Wu, S.C.; Guo, X.; Xia, F.; Hu, X.R.; Song, Z.; Zhang, C.; Qin, Q.Q.; Fu, B.D.; et al. Inhibitory effects of alpha-cyperone on adherence and invasion of avian pathogenic Escherichia coli O78 to chicken type II pneumocytes. Vet. Immunol. Immunopathol. 2014, 159, 50–57. [Google Scholar] [CrossRef]
- Kim, S.J.; Kim, H.J.; Kim, H.J.; Jang, Y.P.; Oh, M.S.; Jang, D.S. New patchoulane-type sesquiterpenes from the rhizomes of Cyperus rotundus. Bull. Korean Chem. Soc. 2012, 33, 3115–3118. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Ryu, B.; Kim, H.-Y.; Yang, Y.-I.; Ham, J.; Choi, J.-H.; Jang, D.S. Sesquiterpenes from the rhizomes of Cyperus rotundus and their potential to inhibit LPS-induced nitric oxide production. Bull. Korean Chem. Soc. 2013, 34, 2207–2210. [Google Scholar] [CrossRef] [Green Version]
- Ryu, B.; Kim, H.M.; Lee, J.-S.; Cho, Y.J.; Oh, M.S.; Choi, J.-H.; Jang, D.S. Sesquiterpenes from rhizomes of Cyperus rotundus with cytotoxic activities on human cancer cells in vitro. Helv. Chim. Acta 2015, 98, 1372–1380. [Google Scholar] [CrossRef]
- Sim, Y.; Choi, J.G.; Gu, P.S.; Ryu, B.; Kim, J.H.; Kang, I.; Jang, D.S.; Oh, M.S. Identification of Neuroactive Constituents of the Ethyl Acetate Fraction from Cyperi Rhizoma Using Bioactivity-Guided Fractionation. Biomol. Ther. 2016, 24, 438–445. [Google Scholar] [CrossRef] [Green Version]
- Osteen, K.G.; Yeaman, G.R.; Bruner-Tran, K.L. Matrix metalloproteinases and endometriosis. Semin. Reprod. Med. 2003, 21, 155–164. [Google Scholar] [CrossRef]
- Yang, W.V.; Au, H.K.; Chang, C.W.; Chen, H.W.; Chen, P.H.; Chen, C.C.; Tang, Y.L.; Wang, I.T.; Tzeng, C.R. Matrix remodeling and endometriosis. Reprod. Med. Biol. 2005, 4, 93–99. [Google Scholar] [CrossRef]
- Ribeiro, A.S.; Albergaria, A.; Sousa, B.; Correia, A.L.; Bracke, M.; Seruca, R.; Schmitt, F.C.; Paredes, J. Extracellular cleavage and shedding of P-cadherin: A mechanism underlying the invasive behaviour of breast cancer cells. Oncogene 2010, 29, 392–402. [Google Scholar] [CrossRef] [Green Version]
- Howard, F.M. Endometriosis and mechanisms of pelvic pain. J. Minim. Invasive Gynecol. 2009, 16, 540–550. [Google Scholar] [CrossRef] [PubMed]
- Howe, D.C.; Pullen, N.; Shelton, D.L. Treatment of Endometriosis. 2010. Available online: https://patents.google.com/patent/WO2010029497A1/zh (accessed on 1 October 2020).
- Hennessy, B.T.; Smith, D.L.; Ram, P.T.; Lu, Y.; Mills, G.B. Exploiting the PI3K/AKT pathway for cancer drug discovery. Nat. Rev. Drug Discov. 2005, 4, 988–1004. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Pavone, M.E.; Lu, Z.; Wei, J.; Kim, J.J. Increased activation of the PI3K/AKT pathway compromises decidualization of stromal cells from endometriosis. J. Clin. Endocrinol. Metab. 2012, 97, E35–E43. [Google Scholar] [CrossRef] [Green Version]
- Gentilini, D.; Busacca, M.; Di Francesco, S.; Vignali, M.; Vigano, P.; Di Blasio, A.M. PI3K/Akt and ERK1/2 signalling pathways are involved in endometrial cell migration induced by 17beta-estradiol and growth factors. Mol. Hum. Reprod. 2007, 13, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, X.; Li, C.; He, C.; Ren, B.; Deng, Q.; Gao, W.; Wang, B. Aurora-A modulates MMP-2 expression via AKT/NF-kappaB pathway in esophageal squamous cell carcinoma cells. Acta Biochim. Biophys. Sin. 2016, 48, 520–527. [Google Scholar] [CrossRef] [Green Version]
- Vieira, A.F.; Ribeiro, A.S.; Dionisio, M.R.; Sousa, B.; Nobre, A.R.; Albergaria, A.; Santiago-Gomez, A.; Mendes, N.; Gerhard, R.; Schmitt, F.; et al. P-cadherin signals through the laminin receptor alpha6beta4 integrin to induce stem cell and invasive properties in basal-like breast cancer cells. Oncotarget 2014, 5, 679–692. [Google Scholar] [CrossRef] [Green Version]
- Ozes, O.N.; Mayo, L.D.; Gustin, J.A.; Pfeffer, S.R.; Pfeffer, L.M.; Donner, D.B. NF-kappaB activation by tumour necrosis factor requires the Akt serine-threonine kinase. Nature 1999, 401, 82–85. [Google Scholar] [CrossRef]
- Sizemore, N.; Leung, S.; Stark, G.R. Activation of phosphatidylinositol 3-kinase in response to interleukin-1 leads to phosphorylation and activation of the NF-kappaB p65/RelA subunit. Mol. Cell. Biol. 1999, 19, 4798–4805. [Google Scholar] [CrossRef] [Green Version]
- Cao, W.G.; Morin, M.; Sengers, V.; Metz, C.; Roger, T.; Maheux, R.; Akoum, A. Tumour necrosis factor-alpha up-regulates macrophage migration inhibitory factor expression in endometrial stromal cells via the nuclear transcription factor NF-kappaB. Hum. Reprod. 2006, 21, 421–428. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Ramos, R.; Donnez, J.; Defrere, S.; Leclercq, I.; Squifflet, J.; Lousse, J.C.; Van Langendonckt, A. Nuclear factor-kappa B is constitutively activated in peritoneal endometriosis. Mol. Hum. Reprod. 2007, 13, 503–509. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Ramos, R.; Rocco, J.; Rojas, C.; Sovino, H.; Poch, A.; Kohen, P.; Alvarado-Diaz, C.; Devoto, L. Physiologic activation of nuclear factor kappa-B in the endometrium during the menstrual cycle is altered in endometriosis patients. Fertil. Steril. 2012, 97, 645–651. [Google Scholar] [CrossRef]
- Grund, E.M.; Kagan, D.; Tran, C.A.; Zeitvogel, A.; Starzinski-Powitz, A.; Nataraja, S.; Palmer, S.S. Tumor necrosis factor-alpha regulates inflammatory and mesenchymal responses via mitogen-activated protein kinase kinase, p38, and nuclear factor kappaB in human endometriotic epithelial cells. Mol. Pharmacol. 2008, 73, 1394–1404. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahn, J.-H.; Choi, J.-M.; Kang, E.-S.; Yoo, J.-H.; Cho, Y.-J.; Jang, D.S.; Choi, J.-H. The Anti-Endometriotic Effect of Cyperi Rhizoma Extract, Inhibiting Cell Adhesion and the Expression of Pain-Related Factors through Akt and NF-kB Pathways. Medicina 2022, 58, 335. https://doi.org/10.3390/medicina58030335
Ahn J-H, Choi J-M, Kang E-S, Yoo J-H, Cho Y-J, Jang DS, Choi J-H. The Anti-Endometriotic Effect of Cyperi Rhizoma Extract, Inhibiting Cell Adhesion and the Expression of Pain-Related Factors through Akt and NF-kB Pathways. Medicina. 2022; 58(3):335. https://doi.org/10.3390/medicina58030335
Chicago/Turabian StyleAhn, Ji-Hye, Jun-Myeong Choi, Eun-Sol Kang, Jae-Hyeon Yoo, Yoon-Jin Cho, Dae Sik Jang, and Jung-Hye Choi. 2022. "The Anti-Endometriotic Effect of Cyperi Rhizoma Extract, Inhibiting Cell Adhesion and the Expression of Pain-Related Factors through Akt and NF-kB Pathways" Medicina 58, no. 3: 335. https://doi.org/10.3390/medicina58030335
APA StyleAhn, J.-H., Choi, J.-M., Kang, E.-S., Yoo, J.-H., Cho, Y.-J., Jang, D. S., & Choi, J.-H. (2022). The Anti-Endometriotic Effect of Cyperi Rhizoma Extract, Inhibiting Cell Adhesion and the Expression of Pain-Related Factors through Akt and NF-kB Pathways. Medicina, 58(3), 335. https://doi.org/10.3390/medicina58030335