Growth and Production of Lipids in Raphidocelis subcapitata Immobilized in Sodium Alginate Beads

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemical Products
2.2. Microalga and Culture Medium
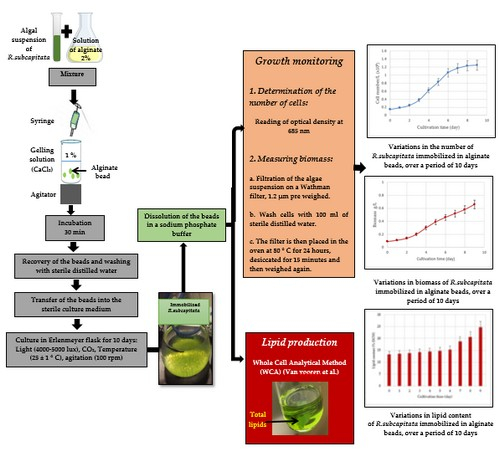
2.3. Cell Immobilization
2.4. Culture Conditions
2.5. Growth Monitoring
2.6. Calculation of Growth Parameters
2.7. Lipid Extraction from Immobilized Microalgae
2.8. Calculating Lipid Productivity
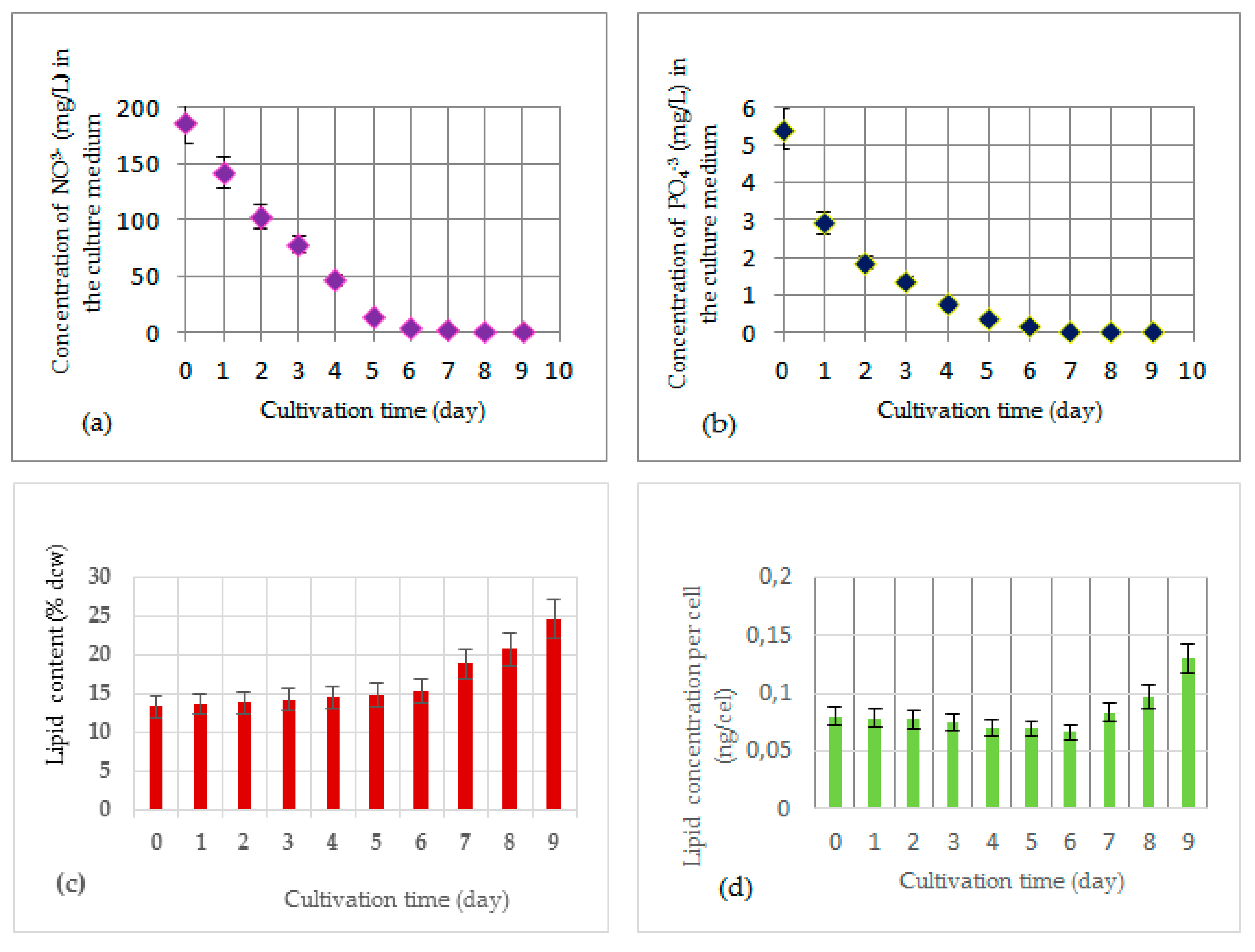
2.9. Measurement of Nitrates in the Culture Medium
2.10. Measurement of Phosphates in the Culture Medium
2.10.1. Principle of the Method
2.10.2. Reagents
- -
- Sulfuric acid, H2SO4, 5N.
- -
- Potassium antimonyl tartrate solution, K(SbO)C4H4O6, 1/2H2O, 3.42 g/L.
- -
- Ammonium molybdate solution, (NH4)6Mo7O24, 4H2O, 40 g/L.
- -
- Ascorbic acid, 0.1 M.
2.10.3. Procedure
3. Results
3.1. Growth of R. subcapitata Immobilized in Alginate Beads
3.2. Production of Total Lipids in R. subcapitata Immobilized in Alginate Beads
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gavrilescu, M.; Chisti, Y. Biotechnology—A sustainable alternative for chemical industry. Biotechnol. Adv. 2005, 23, 471–499. [Google Scholar] [CrossRef] [PubMed]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; Hussain, M.Z.; Prasad, S.; Banerjee, U.C. Prospects of biodiesel production from microalgae in India. Renew. Sustain. Energy Rev. 2009, 13, 2361–2372. [Google Scholar] [CrossRef]
- Li, Y.; Horsman, M.; Wu, N.; Lan, C.Q.; Dubois-Calero, N. Biocatalysts and bioreactor design. Biotechnol. Prog. 2008, 24, 815–820. [Google Scholar]
- Lam, M.K.; Lee, K.T. Renewable and sustainable bioenergies production from palm oil mill effluent (POME): Win–win strategies toward better environmental protection. Biotechnol. Adv. 2011, 29, 124–141. [Google Scholar] [CrossRef]
- Doré-Deschênes, F. Utilisation des Microalgues Comme Source d’énergie Durable; Université de Sherbrooke: Sherbrooke, QC, Canada, 2009. [Google Scholar]
- Cantin, I. La Production de Biodiesel à Partir des Microalgues Ayant un Métabolisme Hétérotrophe; Canada Centre Universitaire de Formation en Environnement Université de Sherbrooke: Sherbrooke, QC, Canada, 2010. [Google Scholar]
- Patil, V.; Källqvist, T.; Olsen, E.; Vogt, G.; Gislerød, H.R. Fatty acid composition of 12 microalgae for possible use in aquaculture feed. Aquac. Int. 2007, 15, 1–9. [Google Scholar] [CrossRef]
- Gonçalves, A.L.; Pires, J.C.; Simões, M. Lipid production of Chlorella vulgaris and Pseudokirchneriella subcapitata. Int. J. Energy Environ. Eng. 2013, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Sander, K.; Murthy, G.S. Life cycle analysis of algae biodiesel. Int. J. Life Cycle Assess. 2010, 15, 704–714. [Google Scholar] [CrossRef]
- Moreira, S.M.; Moreira-Santos, M.; Guilhermino, L.; Ribeiro, R. Immobilization of the marine microalga Phaeodactylum tricornutum in alginate for in situ experiments: Bead stability and suitability. Enzym. Microb. Technol. 2006, 38, 135–141. [Google Scholar] [CrossRef]
- Cao, Y.R.; Liu, Z.; Cheng, G.L.; Jing, X.B.; Xu, H. Exploring single and multi-metal biosorption by immobilized spent Tricholoma lobayense using multi-step response surface methodology. Chem. Eng. J. 2010, 164, 183–195. [Google Scholar] [CrossRef]
- Moreno-Garrido, I. Microalgae immobilization: Current techniques and uses. Bioresour. Technol. 2008, 99, 3949–3964. [Google Scholar] [CrossRef] [PubMed]
- Draget, K.I.; Steinsvåg, K.; Onsøyen, E.; Smidsrød, O. Na- and K-alginate; effect on Ca2+-gelation. Carbohydr. Polym. 1998, 35, 1–6. [Google Scholar] [CrossRef]
- Pires, J.; Gonçalves, A.; Martins, F.; Alvim-Ferraz, M.; Simões, M. Effect of light supply on CO2 capture from atmosphere by Chlorella vulgaris and Pseudokirchneriella subcapitata. Mitig. Adapt. Strateg. Glob. Chang. 2014, 19, 1109–1117. [Google Scholar] [CrossRef] [Green Version]
- Taleb, A. Production de Biodiesel à Partir des Microalgues: Recherche des Souches Accumulatrices des Lipides et Optimisation des Conditions de Culture en Photobioréacteurs. Ph.D. Thesis, Degree-Granting University, Nantes, France, 2015. [Google Scholar]
- Rahbani, J.N.; Salameh, D.F.; Rahbani, J.N. Identifier des microalgues pour le traitement des eaux usées et la production de biocarburant. Available online: https://scholar.google.com/scholar?hl=fr&as_sdt=0%2C5&q=17.%09Rahbani%2C+J.N.%3B+Salameh%2C+D.F.%3B+Rahbani% (accessed on 18 December 2019).
- Chader, S.; Mahmah, B.; Chetehouna, K.; Mignolet, E. Biodiesel production using Chlorella sorokiniana a green microalga. Rev. Energ. Renouvelables 2011, 14, 21–26. [Google Scholar]
- Béliveau, M.-O.; Bertrand-Lehouillier, V.; Carrier, J.; Tremblay, J.; Larrivée, M. Optimisation de la Culture de Microalgues Pour la Production de Biodiesel. 2013. Available online: https://scholar.google.com/scholar?hl=fr&as_sdt=0%2C5&q=19.%09B%C3%A9liveau%2C+M.-O.%3B+Bertrand-Lehouillier%2C+V.%3B+Carrier%2C+J.%3B+Tremblay%2C+J.%3B+Larriv%C3%A9e%2C+M.+Optimisation+de+la+Culture+de+Microalgues+Pour+la+Production+de+Biodiesel%3B+2013.&btnG= (accessed on 18 December 2019).
- Chamoumi, M.; Faucheux, N.; Jones, J.P.; Brunel, D. Optimisation de la Production du Biodiesel à Partir d’huiles de Microalgues et d’huiles Usées. Producteur non Identifié. Ph.D. Thesis, Centre universitaire de Formation en Environnement, Sherbrooke, QC, Canada, 2013. [Google Scholar]
- Massart, A.; Aubry, É.; Hantson, A.-L. Étude de stratégies de culture de Dunaliella tertiolecta combinant haute densité cellulaire et accumulation de lipides en vue de produire du biodiesel. BASE 2010, 14. Available online: https://scholar.google.com/scholar?hl=fr&as_sdt=0%2C5&q=21.%09Massart%2C+A.%3B+Aubry%2C+%C3%89.%3B+Hantson%2C+A.-L.+%C3%89tude+de+strat%C3%A9gies+de+culture+de+Dunaliella+tertiolecta+combinant+haute+densit%C3%A9+cellulaire+et+accumulation+de+lipides+en+vue+de+produire+du+biodiesel.+BASE+2010%2C+14.&btnG= (accessed on 18 December 2019).
- Olofsson, M.; Lamela, T.; Nilsson, E.; Bergé, J.P.; Del Pino, V.; Uronen, P. Seasonal variation of lipids and fatty acids of the microalgae Nannochloropsis oculata grown in outdoor large-scale photobioreactors. Energies 2012, 5, 1577–1592. [Google Scholar] [CrossRef] [Green Version]
- Branco-Vieira, M.; San Martin, S.; Agurto, C.; Santos, M.A.; Freitas, M.A.; Mata, T.M. Potential of Phaeodactylum tricornutum for biodiesel production under natural conditions in Chile. Energies 2018, 11, 54. [Google Scholar] [CrossRef] [Green Version]
- Casadevall, E.; Bailliez, C.; Berkaloff, C.; Brenckmann, F.; Chirac, C.; Corre, B. Production of renewable hydrocarbons through the culture of green unicellular alga Botryococcus: Study of the factors affecting the production of hydrocarbons: Final report. Production d’hydrocarbures renouvelables par la culture de l’algue verte botryococcus braunii: Etude des facteurs affectant la production des hydrocarbures: Rapport final. 1985. Available online: https://www.osti.gov/etdeweb/biblio/7769795 (accessed on 18 December 2019).
- Panouillé, M.; Larreta-Garde, V. Gelation behaviour of gelatin and alginate mixtures. Food Hydrocoll. 2009, 23, 1074–1080. [Google Scholar]
- Khromova, Y.L. The effect of chlorides on alginate gelation in the presence of calcium sulfate. Colloid J. 2006, 68, 115–119. [Google Scholar] [CrossRef]
- Polona, S.; Marija, B.; Anamarija, Z.; Odon, P.; Ales, M. Shape optimization and characterization of polysaccharide beads prepared by ionotropic gelation. J. Microencapsul. 2007, 25, 90–105. [Google Scholar]
- Benasla, A.; Hausler, R. Optimisation of Growth of Raphidocelis subcapitata Immobilised for Biofuel Production: Influence of Alginate and CaCl2 Concentrations on Growth. Environments 2018, 5, 60. [Google Scholar] [CrossRef] [Green Version]
- Environnement et changement climatique Canada. Méthode d’essai Biologique: Essai d’inhibition de la Croissance d’une Algue d’eau Douce.Centre des Sciences et Technologies Environnementales; Environnement et changement climatique Canada: Ottawa, ON, Canada, 2007.
- Santos, M.M.D.; Moreno-Garrido, I.; Gonçalves, F.; Soares, A.M.; Ribeiro, R. An in situ bioassay for estuarine environments using the microalga Phaeodactylum tricornutum. Environ. Toxicol. Chem. 2002, 21, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Voo, W.-P.; Lee, B.-B.; Idris, A.; Islam, A.; Tey, B.-T.; Chan, E.-S. Production of ultra-high concentration calcium alginate beads with prolonged dissolution profile. RSC Adv. 2015, 5, 36687–36695. [Google Scholar] [CrossRef]
- Lam, M.K.; Lee, K.T. Immobilization as a feasible method to simplify the separation of microalgae from water for biodiesel production. Chem. Eng. J. 2012, 191, 263–268. [Google Scholar] [CrossRef]
- Richmond, A. Handbook of Microalgal Culture: Biotechnology and Applied Phycology; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Van Vooren, G.; Le Grand, F.; Legrand, J.; Cuiné, S.; Peltier, G.; Pruvost, J. Investigation of fatty acids accumulation in Nannochloropsis oculata for biodiesel application. Bioresour. Technol. 2012, 124, 421–432. [Google Scholar] [CrossRef] [Green Version]
- Folch, J.; Lees, M.; Stanley, G.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Eaton, A.D.; Clesceri, L.S.; Rice, E.W.; Greenberg, A.E.; Franson, M. APHA: Standard Methods for the Examination of Water and Wastewater, Centennial ed.; APHA: Washington, DC, USA, 2005. [Google Scholar]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Del Río, E.; Armendáriz, A.; García-Gómez, E.; García-González, M.; Guerrero, M.G. Continuous culture methodology for the screening of microalgae for oil. J. Biotechnol. 2015, 195, 103–107. [Google Scholar] [CrossRef] [Green Version]
- Rodolfi, L.; Chini Zittelli, G.; Bassi, N.; Padovani, G.; Biondi, N.; Bonini, G. Microalgae for oil: Strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor. Biotechnol. Bioeng. 2009, 102, 100–112. [Google Scholar] [CrossRef]
- Bernard, O.; Blonz, C.; Hidoine, B. Projet Shamash: Production de Biocarburants Lipidiques par des Microalgues. 2008. Available online: https://scholar.google.com/scholar?hl=fr&as_sdt=0%2C5&q=41.%09Bernard%2C+O.%3B+Blonz%2C+C.%3B+Hidoine%2C+B.+Projet+Shamash%3A+Production+de+Biocarburants+Lipidiques+par+des+Microalgues%3B+&btnG= (accessed on 17 December 2019).
- Dejoye, C. Eco-Extraction et Analyse de Lipides de Micro-Algues Pour la Production D’algo-Carburant; Université d’Avignon: Avignon, France, 2013. [Google Scholar]
- Hersant, G.; Tremblay, A.; Berthelot, L.; Rousseau, J.; Bernier, R. Merinov; ÉPAQ: Grande-Rivière, QC, Canada, 2012. [Google Scholar]
- Del Río, E.; García-Gómez, E.; Moreno, J.; Guerrero, M.G.; García-González, M. Microalgae for oil. Assessment of fatty acid productivity in continuous culture by two high-yield strains, Chlorococcum oleofaciens and Pseudokirchneriella subcapitata. Algal Res. 2017, 23, 37–42. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Microalgae | Type of Culture | Reference |
|---|---|---|
| Parachlorella kessleri UTEX2229 | Free | [16] |
| Chlorella sp. | Free | [17] |
| Chlorella sorokiniana | Free | [18] |
| Chlorella vulgaris | Immobilized | [19] |
| Tetraselmis sp. | Free | [20] |
| Dunaliella tertiolecta | Free | [21] |
| Nannochloropsis oculata | Free | [22] |
| Phaeodactylum tricornutum | Free | [23] |
| Pseudokirchneriella subcapitata | Free | [9] |
| Botryococcus braunii | Immobilized | [24] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benasla, A.; Hausler, R. Growth and Production of Lipids in Raphidocelis subcapitata Immobilized in Sodium Alginate Beads. Energies 2020, 13, 506. https://doi.org/10.3390/en13020506
Benasla A, Hausler R. Growth and Production of Lipids in Raphidocelis subcapitata Immobilized in Sodium Alginate Beads. Energies. 2020; 13(2):506. https://doi.org/10.3390/en13020506
Chicago/Turabian StyleBenasla, Amel, and Robert Hausler. 2020. "Growth and Production of Lipids in Raphidocelis subcapitata Immobilized in Sodium Alginate Beads" Energies 13, no. 2: 506. https://doi.org/10.3390/en13020506
APA StyleBenasla, A., & Hausler, R. (2020). Growth and Production of Lipids in Raphidocelis subcapitata Immobilized in Sodium Alginate Beads. Energies, 13(2), 506. https://doi.org/10.3390/en13020506