Abstract
Understanding the effects of above- and below-ground interactions on seedling growth is pivotal for identifying the key drivers of secondary forest succession. However, it is still unclear whether the effects of above- and below-ground interactions of plants are consistent for seedling growth of deciduous and evergreen species. There are two types of broadleaved forests (i.e., tropical lowland rainforest and tropical deciduous monsoon rainforest) in the low-elevation (<800 m) areas of Hainan Island in China. Here, 32 seedling transplanting plots (1 × 1 m2) were established in the tropical lowland rainforest and the tropical deciduous monsoon rainforest, respectively. Four treatments (each with 16 replicates) were carried out to reduce above- and below-ground interactions of plants in the low-elevation forests: removal of vegetation (R), root trenching (T), removal of vegetation and root trenching (R + T), and no vegetation removal or trenching (as the control) (C). Seedlings of four deciduous species and four evergreen species were planted to observe their performance in the experiments. The relative growth rates (RGR) of the seedlings were measured to distinguish the relative effects of above- and below-ground interactions. The photosynthetically active radiation (PAR) was measured as a proxy for above-ground interaction and the root biomass was used as a proxy for below-ground interaction. The relationships between seedling RGR and PAR/root biomass were examined. Results showed that: (1) R and R+T treatments significantly increased the seedlings RGR, but T treatment had no effect on the RGR; (2) the growth rates of deciduous species were greater than those of the evergreen species; and (3) seedling growth rates were increased with more PAR. Our study suggests that above-ground vegetation removal had a stronger effect than trenching on the growth and assembly of tree seedlings in the low-elevation tropical rainforests.
1. Introduction
Interaction between seedlings is nearly universal in forests. Interactions of plants, can be a critical factor in secondary succession by affecting the speed of forest regeneration [1]. In tropical forests, most seedlings compete with both mature trees and other seedlings, not only in above-ground but also in below-ground parts [2]. It is true that light is usually the most important factor limiting tree seedling growth in tropical forests [3,4]; however, below-ground resources are also important [5]. The result of both the two types of competition between seedlings and adult trees determines the composition of the advanced renewal and relative growth of the seedlings [6]. These, finally, strongly influence the species composition of the forest. Identifying the influences of above- and below-ground interactions of plants on the relative growth of deciduous and evergreen species is necessary to improve our understanding of forest succession.
Seedling relative growth rate (RGR) is a critical factor, reflecting shade tolerance, the regeneration niche, and successional status of plants in their natural habitat [7,8]. It is well known that species differences in seedling RGR across environmental or resources (i.e., light, water, and nutrients) gradients result in the maintenance of species biodiversity in forest ecosystems [7]. Many works have examined the effects of gaps on seedling survival and growth in different types of tropical rainforest, but there have been very few studies on the integrative effects of vegetation removal and root trenching on seedling growth and performance [9,10,11]. Besides, the research on the response of deciduous and evergreen species seedlings’ RGR to above- and below-ground interactions of plants is less frequent.
In most tropical evergreen and monsoon forests, the removal of vegetation can promote seedling growth from the seedling bank [12,13]. Hence, the factors limiting seedling survival and growth are of great importance in determining the species presence, density, and relative size, which, in turn, can be crucial in determining the trajectories of forest succession. The relative competitiveness of deciduous and evergreen species seedlings determines the successional direction of tropical forests [14]. In the early stage of tropical rainforest succession, there are many deciduous species seedlings which can tolerate intense light, drought, and harsh soil conditions [15]. As the canopy density increased, the evergreen species appeared and replaced the deciduous species eventually in the tropical forest.
Tropical lowland rainforest and tropical deciduous monsoon rainforest, which are similar in terms of elevation (≤800 m), are two typical forest types on Hainan Island, China. Tropical lowland rainforest occupies the largest area at low elevations, while tropical deciduous monsoon rainforest occurs in drought and hot habitats. In the previous studies, we found that the species are characterized by deciduous species in tropical deciduous monsoon rainforest and by evergreen in the tropical lowland rainforest [16,17,18]. Liu [14] found that both the richness and abundance of deciduous species in the tropical deciduous monsoon rainforest were higher than those in the tropical lowland rainforest. Studying the response of deciduous and evergreen species to different treatments of reducing plant interactions in both above- and below-ground parts will help us to uncover the mechanism of species succession in tropical rainforests in the low-elevation areas of Hainan Island. In this study, plant interactions in the above-ground part were reduced by clearing understory vegetation, while plant interactions in the below-ground parts were reduced by trenching the soil around the tree seedlings. Four representative deciduous species and four evergreen species in the tropical low-elevation areas were planted in four treatments to reduce interactions of plants in the above- and below-ground parts. The relative growth rates of seedlings were measured. We hypothesize that the deciduous species seedlings regenerate earlier and grow better than the evergreen species in the lowland rainforests in our study area. Our two objectives are: (1) to distinguish the relative effects of above- and below-ground interactions on seedling performance, and (2) to explore whether deciduous or evergreen species will predominate in the natural recovery process of the disturbed lowland secondary forests.
2. Material and Methods
2.1. Study Area
This study was conducted in the Bawangling Forest Region (BFR; 18°52′−19°12′ N, 108°53′–109°20′ E) on Hainan Island, China. The area of BFR is 500 km2 with an elevation range of 100–1654 m.a.s.l. The precipitation is seasonally distributed, with a wet season (monthly precipitation > 100 mm) from May to October and a dry season from November to April. The annual precipitation is 1677 mm. The mean temperature is 23.6 °C. The lowland area of the study site is mainly dominated by secondary forest recovering from shifting cultivation and logging, while some fragments of old growth forest are scattered across the landscape in low-elevation areas.
2.2. Data Collection
Interactions of plants in the above-ground parts were reduced by clearing understory vegetation and interactions of plants in the below-ground parts were reduced by trenching the soil around the tree seedlings. A total of 32 seedling transplanting plots were established in the tropical lowland rainforest and the tropical deciduous monsoon rainforest, respectively. In total, sixty-four 1 × 1 m2 plots were established in our study area. The replicates of plots for each treatment was 16. The following four kinds of treatments in these plots were performed: removal of vegetation, root trenching, removal of vegetation + root trenching, and control. Removal of vegetation treatment included the removal of all woody stems within 0.5 m of the plot edge to improve light transmittance. On all four sides of each trenched plot, we dug trenches 50 cm deep (at this depth, 93% of the live roots can be removed [4,19]) and 40 cm wide. We covered it with a thick plastic sheet to prevent neighboring roots from invading, and then refilled it.
Photosynthetically active radiation (PAR) of the plot can surrogate above-ground interactions of plants and root biomass, whereas can indicate below-ground interactions of plants. To test the effects of the four treatments on above- and below-ground interactions, we measured the PAR and root biomass in each plot. We measured the PAR every 2 h from 8:00 a.m. to 18:00 p.m., with Field SCOUT light quantum illuminance radio meter (3415FSE, Spectrum Technologies, INc., Plainfield, IL, USA) in each plot on a sunny day. We set 5 points uniformly along the diagonal of each plot, and then measured the PAR at the height of 0 cm, 50 cm, and 150 cm from the ground at each point. Meanwhile, 4 soil scores (100 cm2) were collected using cutting rings to measure fine root biomass in each plot. We washed the fine root mass carefully by hand with clean water. The fine roots were dried at 70 °C in an oven for 48 h and then weighed.
Seedlings of eight tree species were chosen for our experiment: Sapium discolor (Champ. ex Benth.) Muell. Arg, Lannea coromandelica (Houtt.) Merr, Toxicodendron succedaneum (L.) O. Kuntze, Peltophorum tonkinense (Pierre). Gagnep are canopy dominant species in tropical deciduous monsoon rainforest, and Castanopsis carlesii (Hemsl.) Hayata, Machilus suaveolens S. Lee, Syzygium cumini (L.) Skeels, Vatica mangachapoi Blanco dominate the canopy of the tropical lowland rainforest. Among the eight species, Sapium discolor, Lannea coromandelica, Toxicodendron succedaneum, and Peltophorum tonkinense are dry-season deciduous species, and Castanopsis carlesi, Machilus suaveolens, Syzygium cumini, and Vatica mangachapoi are evergreen species. The annual seedlings of these species were transplanted from the wild to nursery sites where the environmental conditions were more suitable for seedling growth on 5 May 2010. We watered and shaded the seedlings to ensure their survival, and then transplanted the seedlings to the treated test plots before the rainy season (mid-June). We randomly planted one individual of each plant in each plot. Therefore, a total of 512 seedlings were planted (1 individual seedlings × 8 species × 16 plots ×4 treatments). In the first two weeks after transplanting, we replaced the dead seedlings by new individuals.
In August 2010, the heights of all seedlings were measured. During the vegetation removal process in December 2010 and May 2011, we removed all the germinated stems. The height of all the surviving seedlings was measured on 10 January and 10 April 2012, respectively.
2.3. Data Statistical Analysis
We compared two factors (PAR and fine root mass) between different treatments using one-way ANOVA. The relative growth rate (RGR) of each seedling was calculated by the Equation (1) [20]:
where H1 is the seedling height at the end of census, H0 is the beginning height of census, t1 is the end time of census, and t0 is the beginning time of census. After one-way ANOVA analysis, the significant differences in RGR among the four different treatments were found by using Turkey’s multiple comparisons. We used one-way ANOVA to compare the RGR of deciduous and evergreen species. PAR, fine-root biomass, and RGR were log transformed to improve normality and the equality of variances. We used PAR as the independent variable and RGR as the dependent variable. Linear regression was used to analyze the relationship between seedling RGR and PAR, and between RGR and root biomass. Statistical analyses were conducted in R [21].
3. Results
3.1. Effects of Treatment on Photosynthetically Active Radiation and Root Biomass
Compared with the control and root trenching treatments, removal of vegetation treatment significantly increased PAR (Table 1). The trenching treatment significantly reduced root mass (p < 0.001). The PAR serves as a proxy for above-ground interactions of plants and the root dry matter mass serves as proxy for below-ground interactions of plants. These results show that the treatments had a significant effect on the above- and below-ground interactions of plants.

Table 1.
Comparison of photosynthetic effective radiation and root biomass of roots under different treatments. Different letters (a, b) indicate significant difference (p ≤ 0.05).
3.2. Effects of Treatment on the Seedling Relative Growth Rate
The vegetation removal and vegetation removal + trenching treatment significantly increased the seedling RGR for all deciduous and evergreen species (Figure 1). Trenching alone had no significant effects on the seedling RGR. The seedling RGR of deciduous species was significantly higher than that of evergreen species in all of the four treatments (p < 0.05).
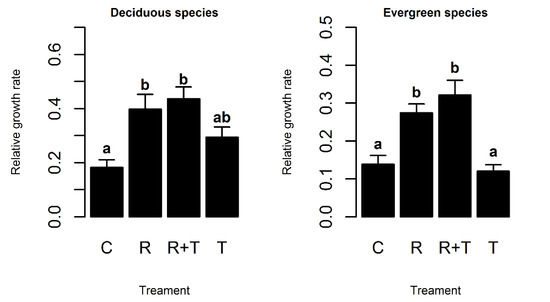
Figure 1.
Effects of different treatments on seedling survival rates. Different letters (a, b) indicate significant difference (p ≤ 0.05).
3.3. Relationship between Photosynthetically Active Radiation and Seedling RGR
The seedling RGR of deciduous and evergreen species was positively correlated with PAR (Figure 2, Table 2). The seedling RGR increased significantly with the increase in PAR in all four of the treatments. The seedling RGR of deciduous species was higher than that of evergreen species, especially in the vegetation removal treatment.
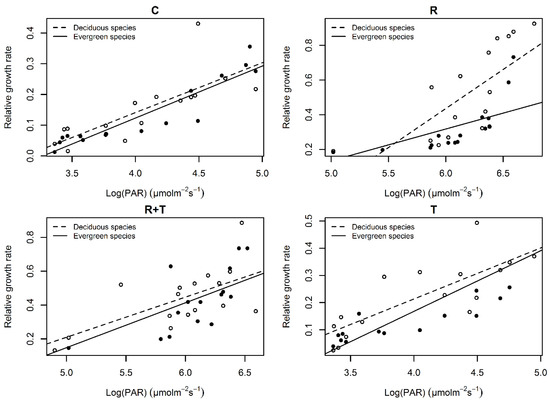
Figure 2.
Relationships between the seedling relative growth rate and photosynthetically active radiation under different treatments.
Table 2.
Linear regression equation parameters for the seedling relative growth rate and photosynthetically active radiation.
3.4. Relationship between Root Biomass and Seedling RGR
The seedling RGR of deciduous and evergreen species was not related to root biomass (Table 3). In all the treatments, the correlations between seedling RGR and root biomass were non-significant (p > 0.05). These results indicate that the effect of below-ground interactions on seedling growth was not significant.
Table 3.
Linear regression equation parameters of the seedling relative growth rate and root biomass.
4. Discussion
4.1. The Seedling Relative Growth Rate in Different Treatments
In our field experiment, we found that removal of vegetation treatment had a greater effect on the seedling relative growth rate than root trenching (Figure 1). The vegetation removal and vegetation removal + trenching treatment significantly increased the seedling RGR for all seedlings. Removal of vegetation treatment improved the light environment under the forest and reduced seedling competition for light, thus significantly increasing the seedling relative growth rate. Our results seemed to be incongruent with other studies in a secondary seasonally dry tropical forest in Panama [22], whose method of trenching was the same as ours. While their results show that trenching treatment significantly improves the growth rate of seedlings in tropical forests, our data suggest that above-ground vegetation removal is a more important driver of seedling growth. The reason for this difference may be that the purpose of their study was just to analyze the effect of trenching on seedling growth in Panama. They did not compare the effects of above-ground vegetation removal and trenching on seedlings growth simultaneously. In their studied system, the underground resources, such as water or nutrients, were probably in shortage, resulting in a strong effect on seedlings. However, in our studied area, understory vegetation was dense, which might pose a serious constraint on the growth of tree seedlings in terms of light availability. In addition, the below-ground resources, such as water and nutrients, were still ample enough for the utilization of all the competing seedlings of plants. The growth of seedling is jointly controlled by factors such as light, water and nutrition [23,24]. Above-ground light intensity can affect seedlings carbon fixation, while underground water, nutrients, and space can affect the seedlings’ survival and growth [25,26]. It has been hypothesized that underground nutrients dominates in the initial stage of succession, but light becomes more important as stand biomass increases [27]. Light is thought to be predominantly asymmetric, because larger plants shade smaller plants and capture a disproportionately larger share of the available light. In contrast, soil moisture and nutrients are often considered symmetric because the root system is assumed to produce consumption zones on its surface, thereby limiting the resource availability of other roots relative to plant size.
These results suggested that light is a limiting factor for seedling growth in the tropical rainforest. In natural forests, light decreased rapidly with depth in the stand, thus a slight increase in height leads to an increase in the amount of intercepting light. Removing the understory vegetation improved the light conditions with most seedlings responded positively. Therefore, this study demonstrated that the understory layer is one of the major limiting factors for seedling growth. In tropical forests, the ability to adapt to low-light conditions is one of the key factors affecting seedling growth, survival, and regeneration. Therefore, light acts as an environmental filter to influence the species composition and the direction of future development of tropical forests. Our results show that light is the most important factor restricting the growth of seedling in the tropical lowland rainforests in the studied forest region, and that the relative growth rate of seedling increases significantly with the increase in photosynthetic effective radiation.
4.2. The Relative Growth Rate of Deciduous and Evergreen Species
The relative growth rate tended to be greater in seedlings of deciduous species than evergreen species across these four treatments. This is consistent with previous studies [8,28]. The difference in the relative growth rates between deciduous and evergreen species may be related to leaf functional traits [29]. Most studies to date have found that a greater RGR of deciduous species seems to be determined by their higher specific leaf area. The higher specific leaf area enhances the carbon capture efficiency of leaves. Consequently, deciduous species with a higher specific leaf area grow rapidly [30]. The difference in growth rate between deciduous and evergreen species suggests that the growth of shade-intolerant deciduous species is more significantly affected by the competition from contiguous trees, while the evergreen species with lower light demands are less affected by the shade from contiguous individuals (Figure 1). These results are consistent with the findings conducted in other forest ecosystems by Zhang et al. [12].
The different relative growth rates among deciduous and evergreen species may have a strong effect on species dynamics on a community level. Therefore, understanding their growth is pivotal for a better understanding of secondary forest dynamics after disturbance. Deciduous species can tolerate intense light, drought, and poor soil, can adapt to the early growth environment of early succession, and can easily establish in disturbed habitats. With the increase in canopy density, shade-tolerant species of tropical lowland species appear and dominate in the lowland tropical forest [31,32]. The change of dominant species likely play an important role in controlling the forest succession dynamics. In early forest restoration, deciduous and evergreen species with significantly different functional traits play different roles and jointly determine the trend of secondary forest succession [12]. Over the past century, the BFR has experienced extensive shifting cultivation and logging in low-elevation areas. These anthropogenic disturbances have greatly altered the forest environment, especially by increasing the amount of light. After human disturbance, deciduous species enter the forest earlier to form deciduous monsoon forest, then evergreen species enter the forest, and finally form tropical lowland rainforest. These findings improve our understanding of the self-recovery mechanisms of tropical forests in secondary succession and provide important theoretical support to accelerate the future restoration of secondary forests.
5. Conclusions
The vegetation removal and vegetation removal + trenching treatment significantly increased the RGR for all seedlings, but trenching alone had no significant effects on seedling RGR. The relationships between light and the relative growth rate of seedling in the different treatments, both for deciduous and evergreen species, suggest that light is a strong limiting factor for seedling growth. Our study suggests that above-ground vegetation removal had a stronger effect than trenching on the growth and assembly of tree seedlings in the low-elevation tropical rainforests.
Author Contributions
Conceptualization, R.Z. and X.L.; methodology, R.Z.; formal analysis, Y.X. and S.Y.; investigation, X.L.; writing—original draft preparation, X.L.; writing—review and editing, R.Z., X.L and H.Z.; visualization, X.L.; project administration, R.Z.; funding acquisition, R.Z. and X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a grant from the Research Institute of Forest Ecology, Environment, and Protection, Chinese Academy of Forestry CAFYBB2016QA006 and by the National Natural Science Foundation of China 31901210.
Acknowledgments
We would like to thank Xiusen Yang, Rucai Li, and Yaocai Tang for their assistance in field work.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Báez, S.; Homeier, J. Functional traits determine tree growth and ecosystem productivity of a tropical montane forest: Insights from a long-term nutrient manipulation experiment. Glob. Chang. Biol. 2018, 24, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Comita, L.S.; Johnson, D.; Chen, M.; Wu, S. Biotic vs abiotic drivers of seedling persistence in a tropical karst forest. J. Veg. Sci. 2017, 28, 206–217. [Google Scholar] [CrossRef]
- Pizano, C.; Mangan, S.A.; Herre, E.A.; Eom, A.-H.; Dalling, J.W. Above- and belowground interactions drive habitat segregation between two cryptic species of tropical trees. Ecology 2011, 92, 47–56. [Google Scholar] [CrossRef]
- Ding, Y.; Zang, R.; Letcher, S.G.; Liu, W.; Lu, X. Aboveground and belowground competition affect seedling performance and allometry in a tropical monsoon forest. New For. 2016, 47, 529–540. [Google Scholar] [CrossRef]
- Larson, J.E.; Funk, J.L. Seedling root responses to soil moisture and the identification of a belowground trait spectrum across three growth forms. New Phytol. 2016, 210, 827–838. [Google Scholar] [CrossRef]
- Ma, Z.; Guo, D.; Xu, X.; Lu, M.; Bardgett, R.D.; Eissenstat, D.M.; McCormack, M.L.; Hedin, L.O. Evolutionary history resolves global organization of root functional traits. Nature 2018, 555, 94–97. [Google Scholar] [CrossRef]
- Xiong, G.; Zhang, A.; Fan, D.; Ge, J.; Yang, D.; Xie, Z.; Zhang, W. Functional coordination between leaf traits and biomass allocation and growth of four herbaceous species in a newly established reservoir riparian ecosystem in China. Ecol. Evol. 2018, 8, 11372–11384. [Google Scholar] [CrossRef]
- Modrzyński, J.; Chmura, D.J.; Tjoelker, M.G. Seedling growth and biomass allocation in relation to leaf habit and shade tolerance among 10 temperate tree species. Tree Physiol. 2015, 35, 879–893. [Google Scholar] [CrossRef]
- De Deyn, G.B. Plant life history and above-belowground interactions: Missing links. Oikos 2017, 126, 497–507. [Google Scholar] [CrossRef]
- Tingstad, L.; Olsen, S.L.; Klanderud, K.; Vandvik, V.; Ohlson, M. Temperature, precipitation and biotic interactions as determinants of tree seedling recruitment across the tree line ecotone. Oecologia 2015, 179, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Goodale, U.M.; Berlyn, G.P.; Gregoire, T.G.; Tennakoon, K.U.; Ashton, M.S. Differences in survival and growth among tropical rain forest pioneer tree seedlings in relation to canopy openness and herbivory. Biotropica 2014, 46, 183–193. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, Q.; Zhou, D.; Xu, W.; Gao, J.; Wang, Z. How evergreen and deciduous trees coexist during secondary forest succession: Insights into forest restoration mechanisms in Chinese subtropical forest. Glob. Ecol. Conserv. 2021, 25, e01418. [Google Scholar] [CrossRef]
- Bai, K.; He, C.; Wan, X.; Jiang, D. Leaf economics of evergreen and deciduous tree species along an elevational gradient in a subtropical mountain. AoB PLANTS 2015, 7. [Google Scholar] [CrossRef]
- Liu, W. Community Ecology of Tropical Monsoon Forest in Hainan Island, China. PhD Dissertation, Chinese Academy of Forestry, Beijing, China, 2009. [Google Scholar]
- Condit, R.; Hubbell, S.P.; Foster, R.B. Changes in tree species abundance in a Neotropical forest: Impact of climate change. J. Trop. Ecol. 1996, 12, 231–256. [Google Scholar] [CrossRef]
- Do, H.T.T.; Grant, J.C.; Trinh, N.B.; Zimmer, H.C.; Tran, L.D.; Nichols, J.D. Recovery of tropical moist deciduous dipterocarp forest in Southern Vietnam. For. Ecol. Manag. 2019, 433, 184–204. [Google Scholar] [CrossRef]
- Liu, W.; Zang, R.; Ding, Y.; Zhang, W. Species-area relationships of different plant functional groups in tropical monsoon rainforests (Hainan Island, China). Pol. J. Ecol. 2013, 61, 3–11. [Google Scholar]
- Ding, Y.; Zang, R.; Letcher, S.G.; Liu, S.; He, F. Disturbance regime changes the trait distribution, phylogenetic structure and community assembly of tropical rain forests. Oikos 2012, 121, 1263–1270. [Google Scholar] [CrossRef]
- Ding, Y.; Zang, R.; Lu, X.; Huang, J.; Xu, Y. The effect of environmental filtering on variation in functional diversity along a tropical elevational gradient. J. Veg. Sci. 2019, 30, 973–983. [Google Scholar] [CrossRef]
- Santiago, L.S.; Wright, S.J.; Harms, K.E.; Yavitt, J.B.; Korine, C.; Garcia, M.N.; Turner, B.L. Tropical tree seedling growth responses to nitrogen, phosphorus and potassium addition. J. Ecol. 2012, 100, 309–316. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Tanner, E.V.J.; Barberis, I.M. Trenching increased growth, and irrigation increased survival of tree seedlings in the understory of a semi-evergreen rain forest in Panama. J. Trop. Ecol. 2007, 23, 257–268. [Google Scholar] [CrossRef][Green Version]
- Baldeck, C.A.; Harms, K.E.; Yavitt, J.B.; John, R.; Turner, B.; Valencia, R.; Navarrete, H.; Bunyavejchewin, S.; Kiratiprayoon, S.; Yaacob, A.; et al. Habitat filtering across tree life stages in tropical forest communities. Proc. R. Soc. B Biol. Sci. 2013, 280. [Google Scholar] [CrossRef]
- Pizano, C.; Mangan, S.A.; Graham, J.H.; Kitajima, K. Habitat-specific positive and negative effects of soil biota on seedling growth in a fragmented tropical montane landscape. Oikos 2014, 123, 846–856. [Google Scholar] [CrossRef]
- Van der Putten, W.; Bardgett, R.; de Ruiter, P.; Hol, W.; Meyer, K.; Bezemer, T.; Bradford, M.; Christensen, S.; Eppinga, M.; Fukami, T.; et al. Empirical and theoretical challenges in aboveground–belowground ecology. Oecologia 2009, 161, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Paz, H. Root/Shoot Allocation and Root Architecture in Seedlings: Variation among Forest Sites, Microhabitats, and Ecological Groups. Biotropica 2003, 35, 318–332. [Google Scholar] [CrossRef]
- Huston, M. Soil Nutrients and Tree Species Richness in Costa Rican Forests. J. Biogeogr. 1980, 7, 147–157. [Google Scholar] [CrossRef]
- Poorter, L.; Rose, S.A. Light-dependent changes in the relationship between seed mass and seedling traits: A meta-analysis for rain forest tree species. Oecologia 2005, 142, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, R.K.; Raghubanshi, A.S.; Singh, J.S. Leaf attributes and tree growth in a tropical dry forest. J. Veg. Sci. 2011, 22, 917–931. [Google Scholar] [CrossRef]
- Antúnez, I.; Retamosa, E.C.; Villar, R. Relative growth rate in phylogenetically related deciduous and evergreen woody species. Oecologia 2001, 128, 172–180. [Google Scholar] [CrossRef]
- De Souza, B.C.; Carvalho, E.C.D.; Oliveira, R.S.; de Araujo, F.S.; de Lima, A.L.A.; Rodal, M.J.N. Drought response strategies of deciduous and evergreen woody species in a seasonally dry neotropical forest. Oecologia 2020, 194, 221–236. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Chi, X.; Tang, Z.; Jiang, M. Analysing tree–neighbourhood interactions in ecotones of montane evergreen and deciduous forests in China. J. Veg. Sci. 2019, 30, 654–663. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).