
Isolation and Characterization of the Sulfate Transporter Gene Family and Its Expression Pattern in Response to Selenium and Abiotic Stress in Walnuts (Juglans regia L.)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Genome-Wide Identification and Physicochemical Analysis of JrSULTR Genes
2.2. Phylogenetic Tree and Collinearity Analysis of JrSULTR Genes
2.3. Gene structure, Domain, and Motif Analysis of JrSULTR Genes
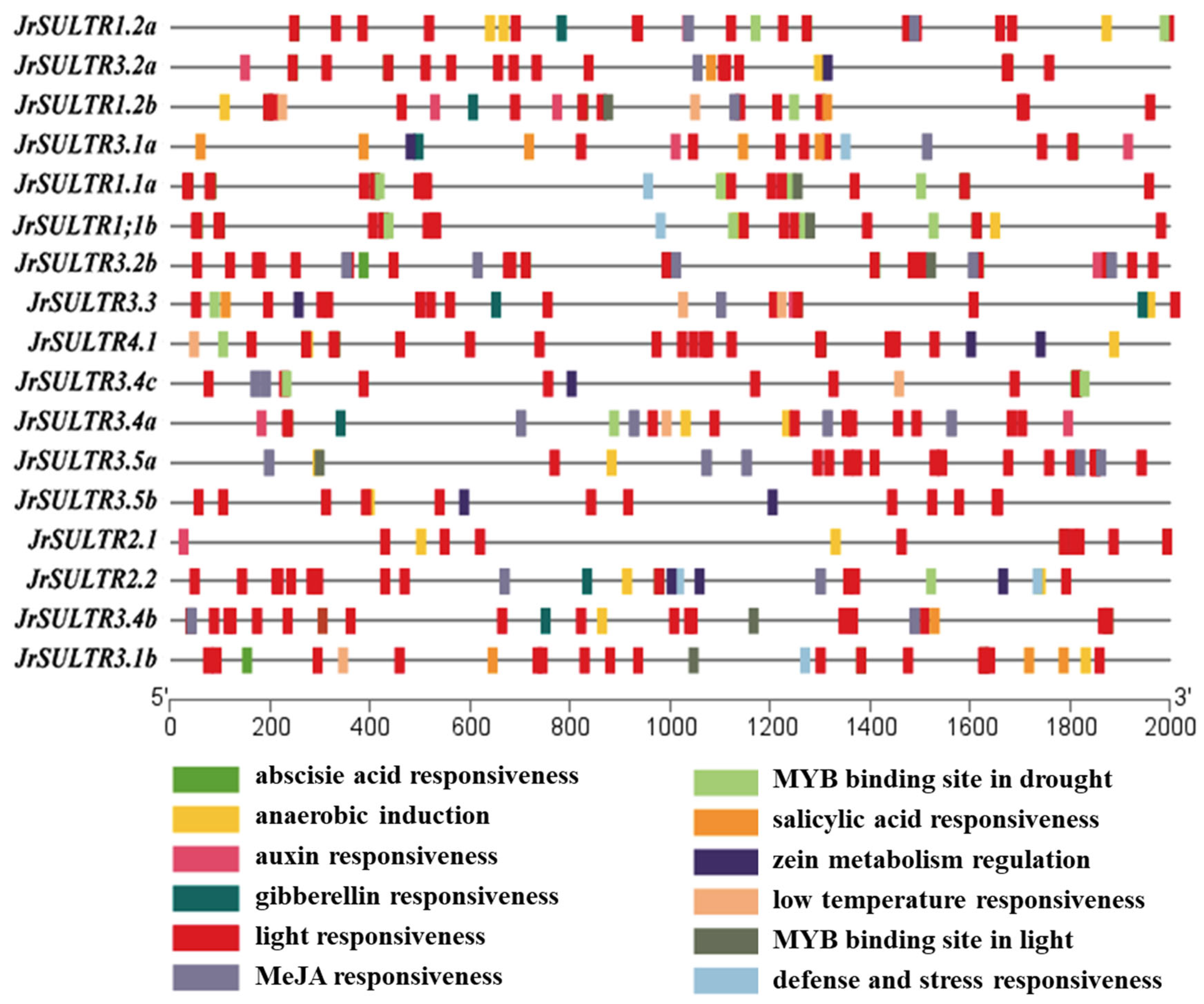
2.4. Analysis of Cis-Acting Elements in the Promoter of JrSULTR Genes
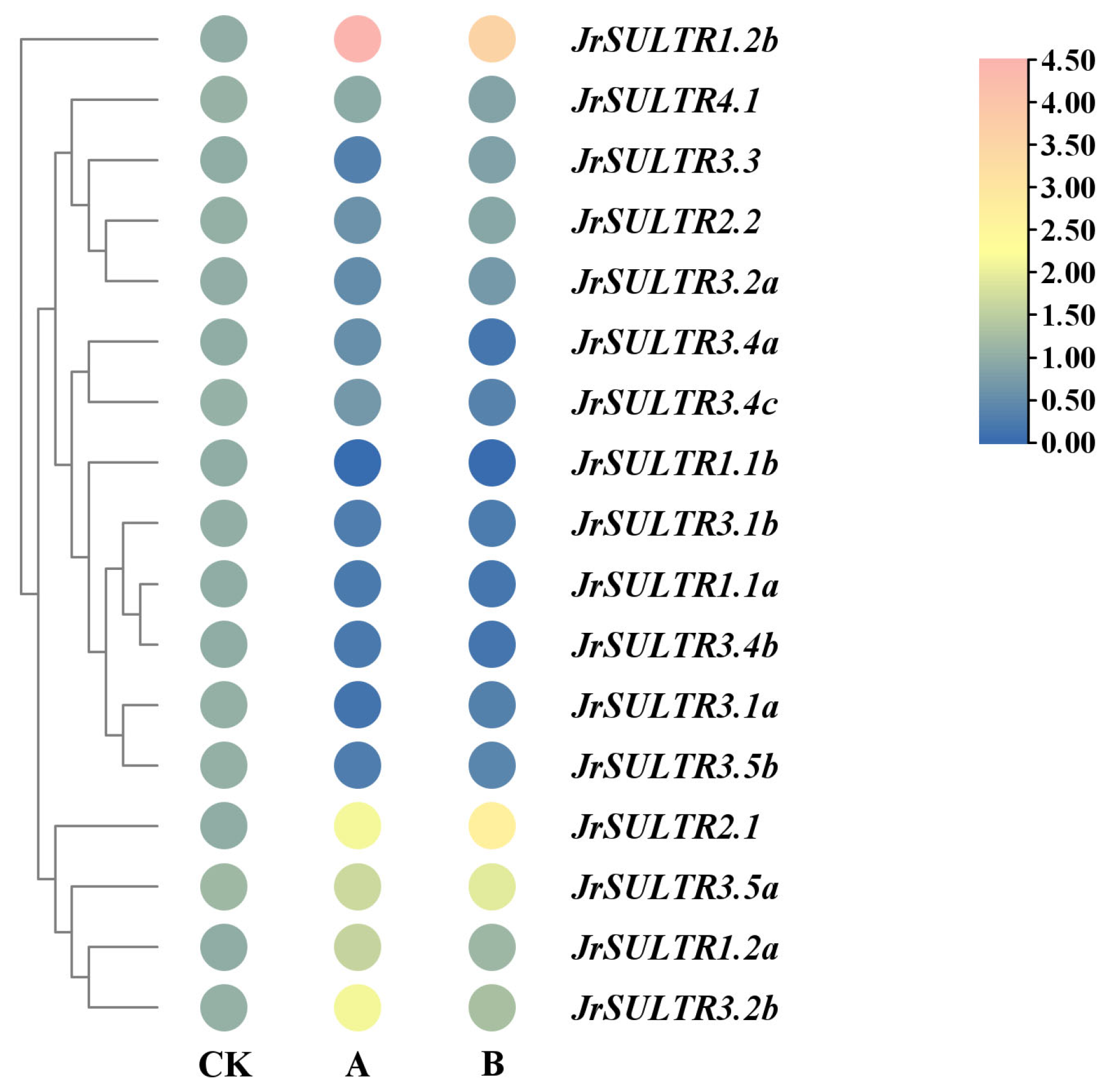
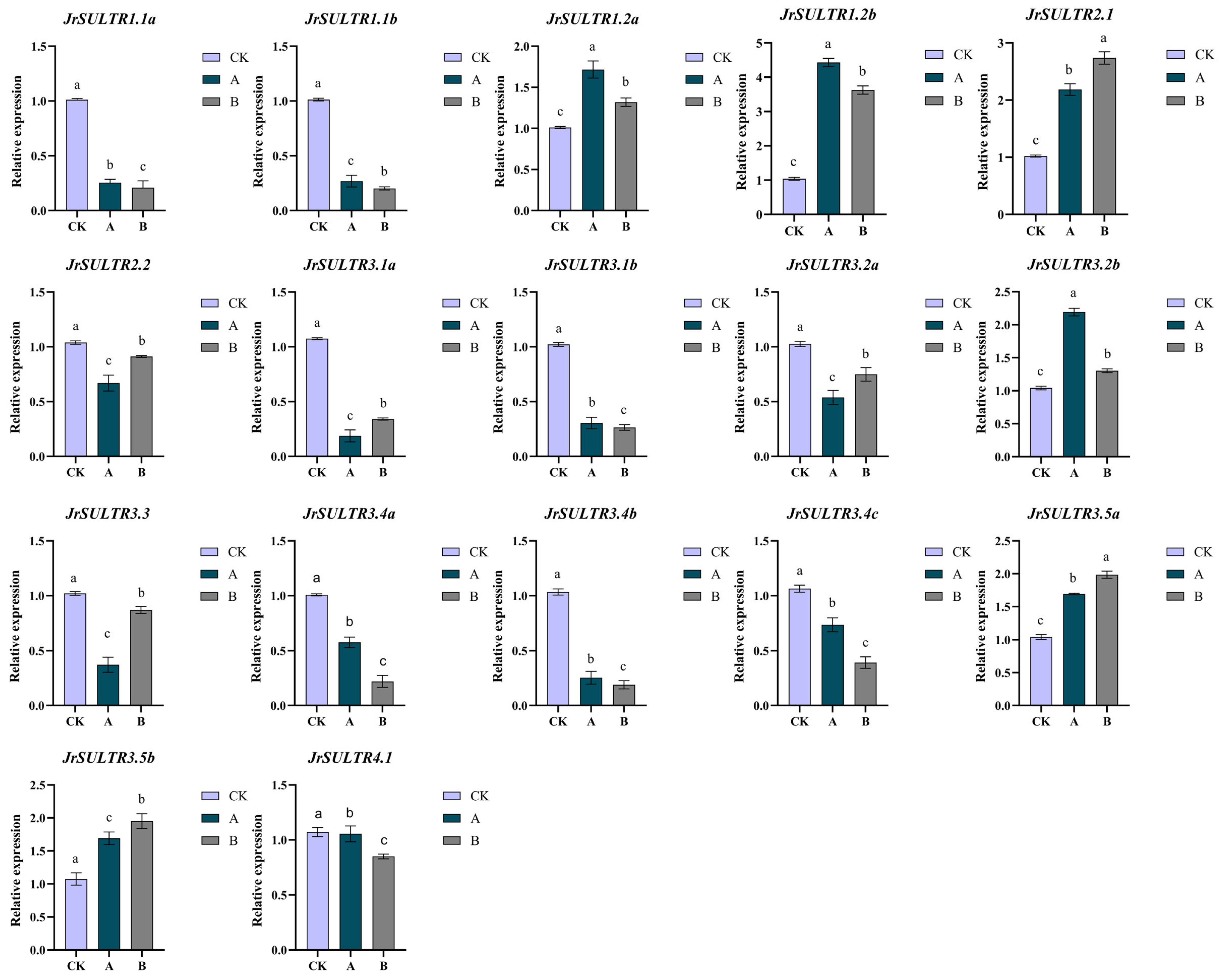
2.5. Transcriptome Data Analysis of the JrSULTR Genes
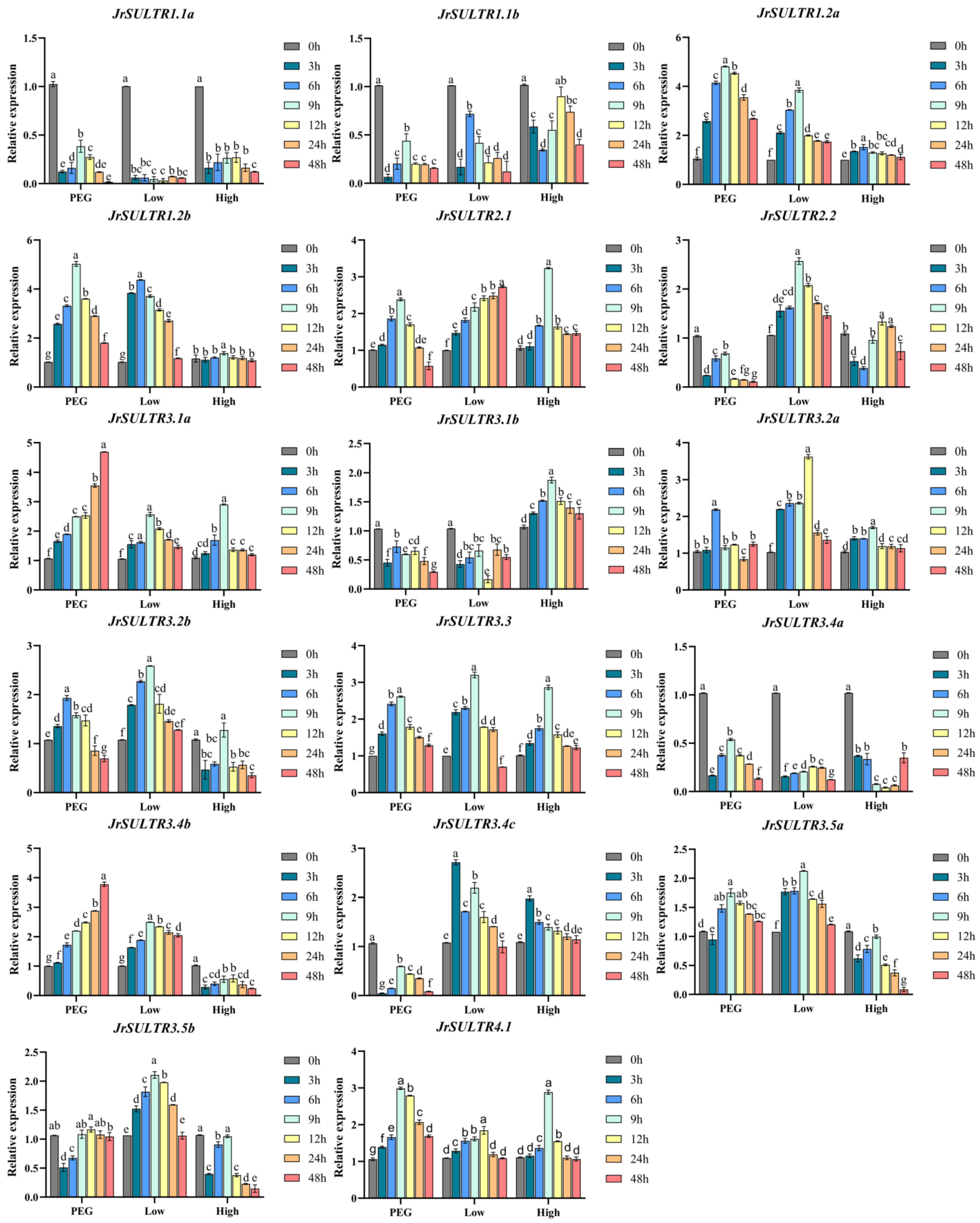
2.6. The Expression Analysis of the JrSULTR Genes under Different Abiotic Stresses
2.7. Subcellular Determination of JrSULTR1.2b and JrSULTR3.1a
3. Results
3.1. Identification and Physicochemical Property Analysis of the JrSULTR Genes
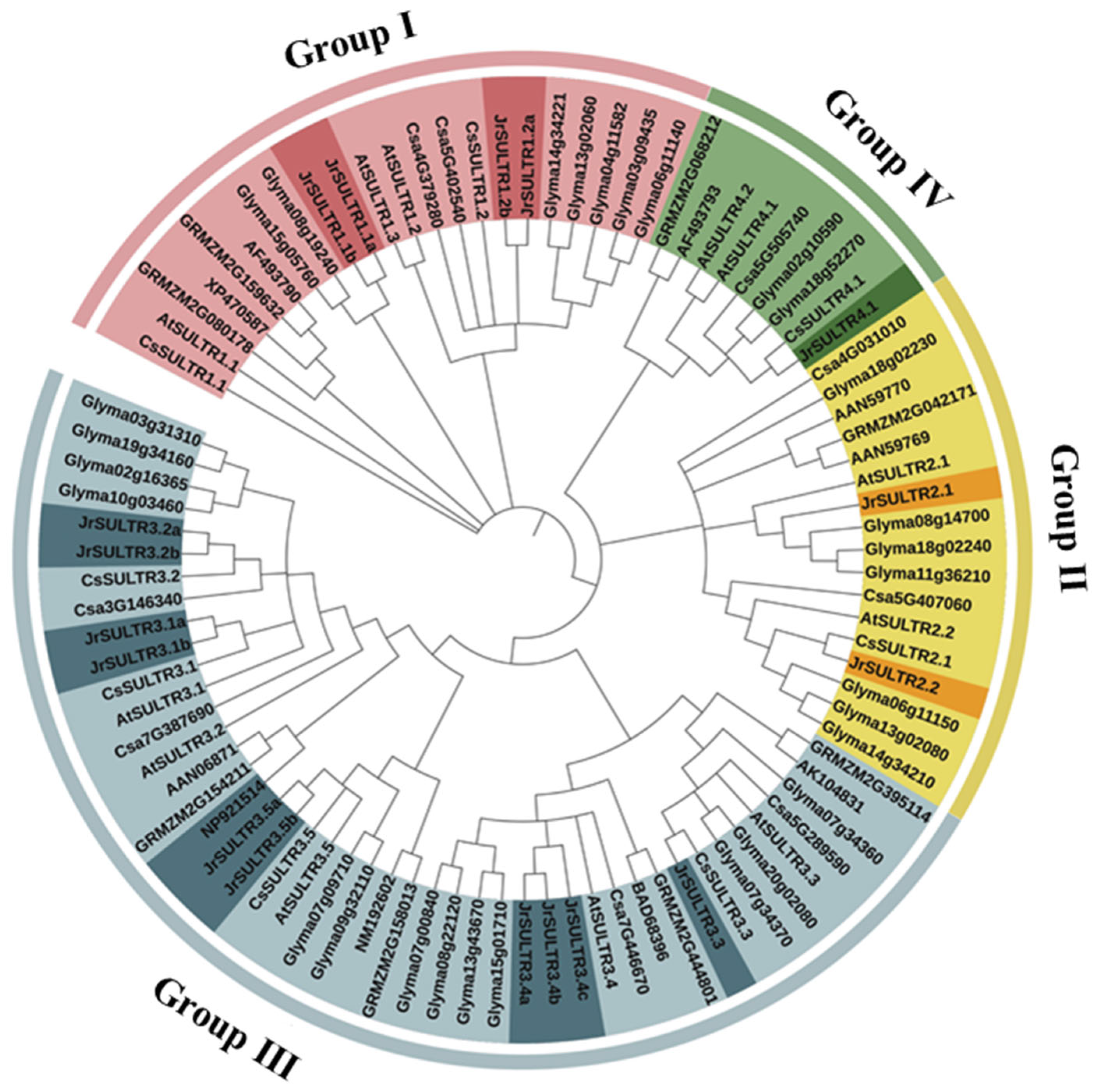
3.2. Phylogenetic Tree Analysis of the JrSULTR Genes
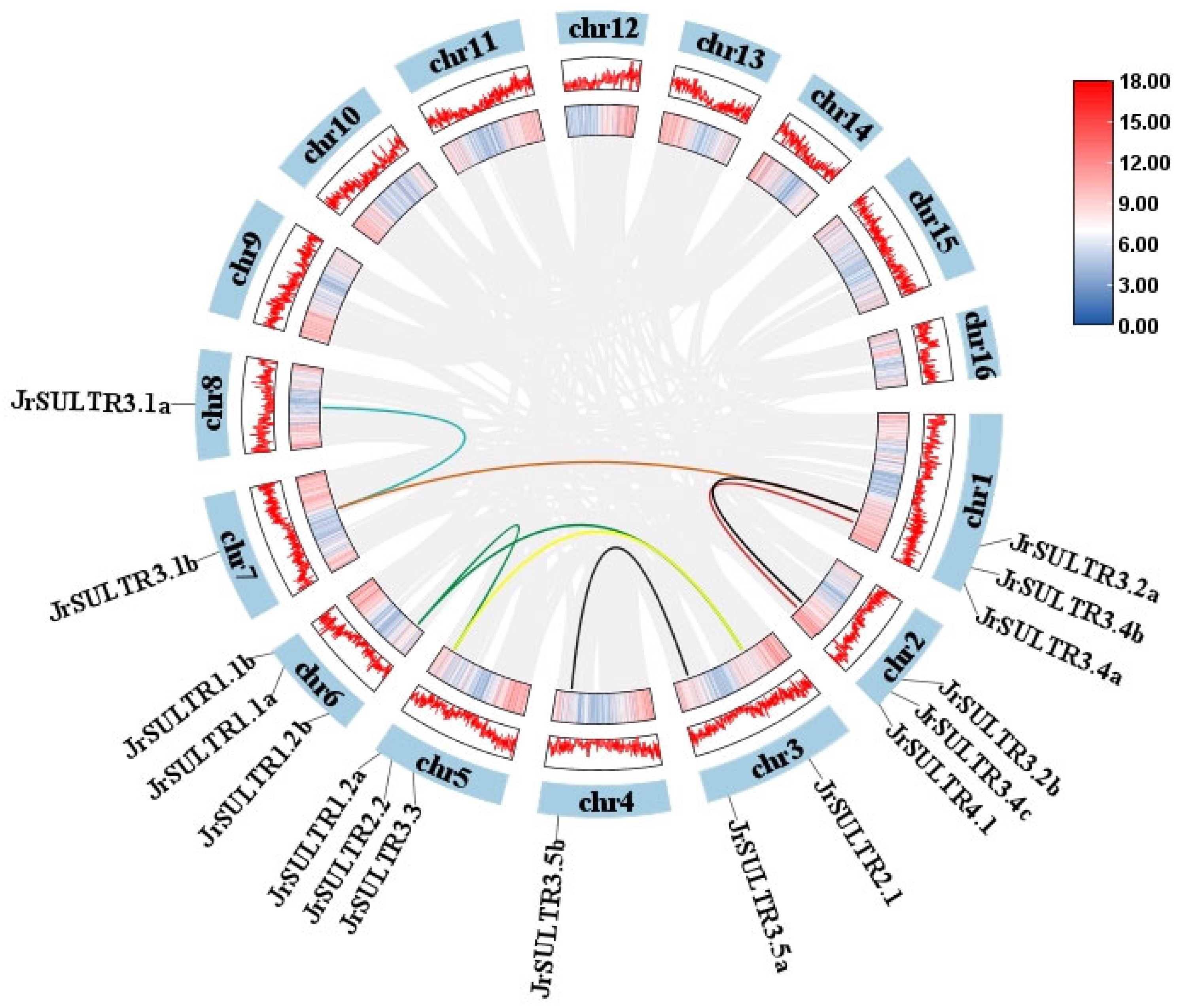
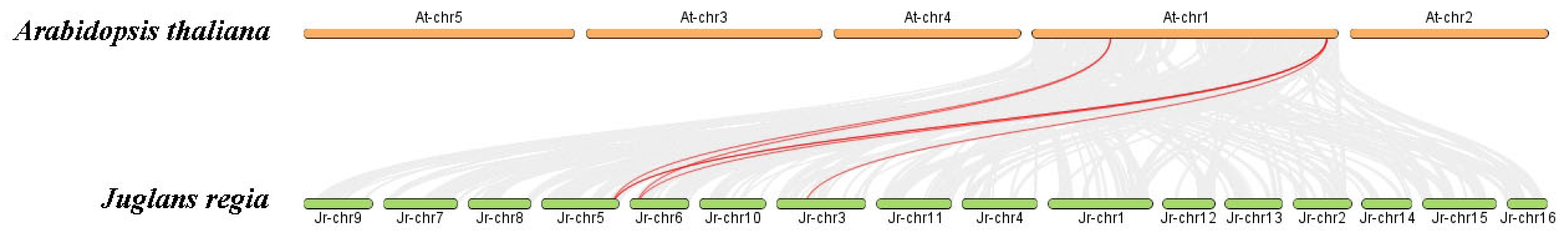
3.3. Chromosomal Localization and Collinearity Analysis of the JrSULTR Genes
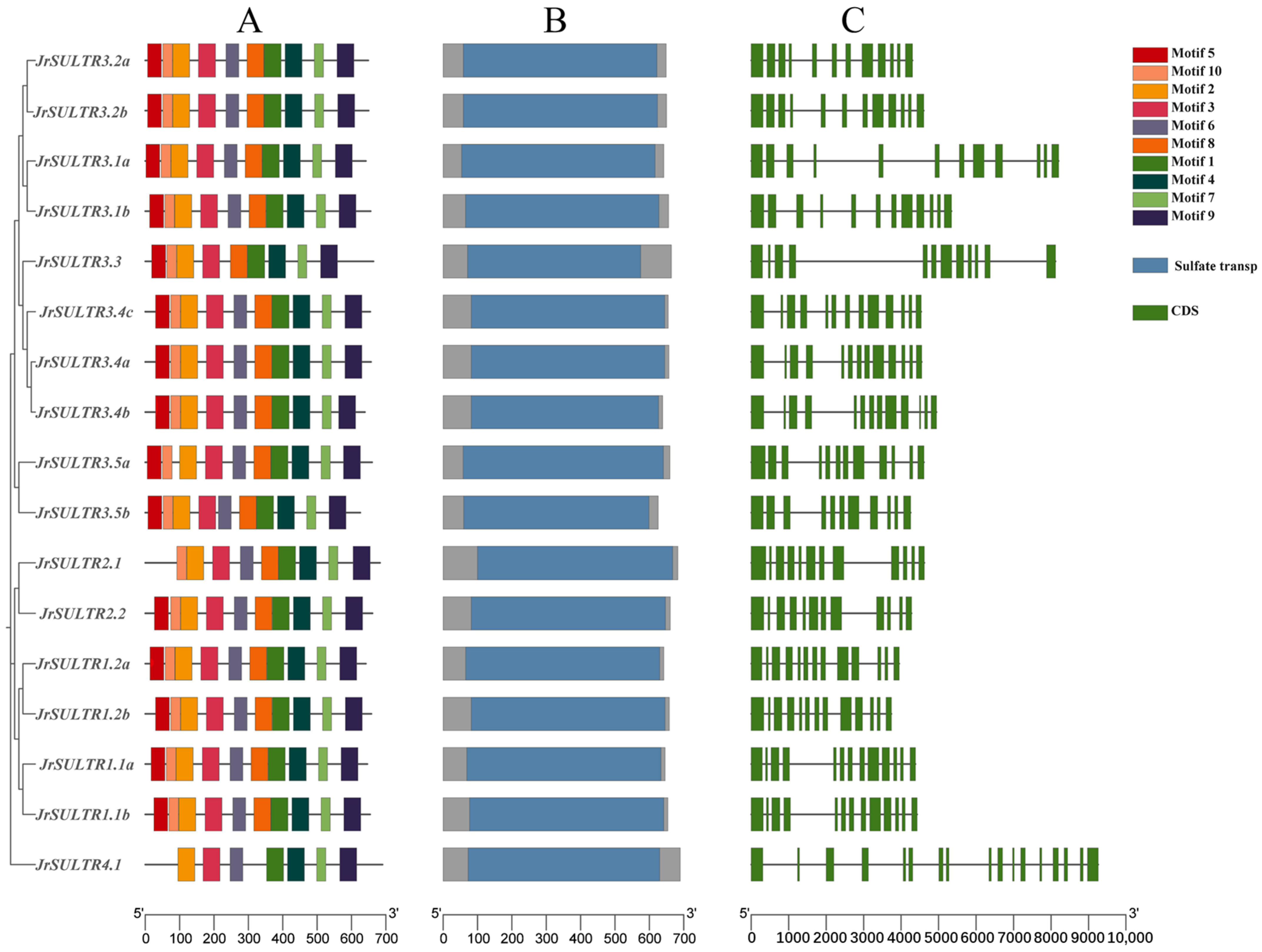
3.4. Gene Structures and Conserved Motif Analysis of the JrSULTR Genes
3.5. Analysis of Cis-Acting Elements in the Promoter of JrSULTR Genes
3.6. Expression Patterns of the JrSULTR Genes in Transcriptome under Sodium Selenite Treatment
3.7. The Expression Level of JrSULTR Genes under Different Abiotic Stresses
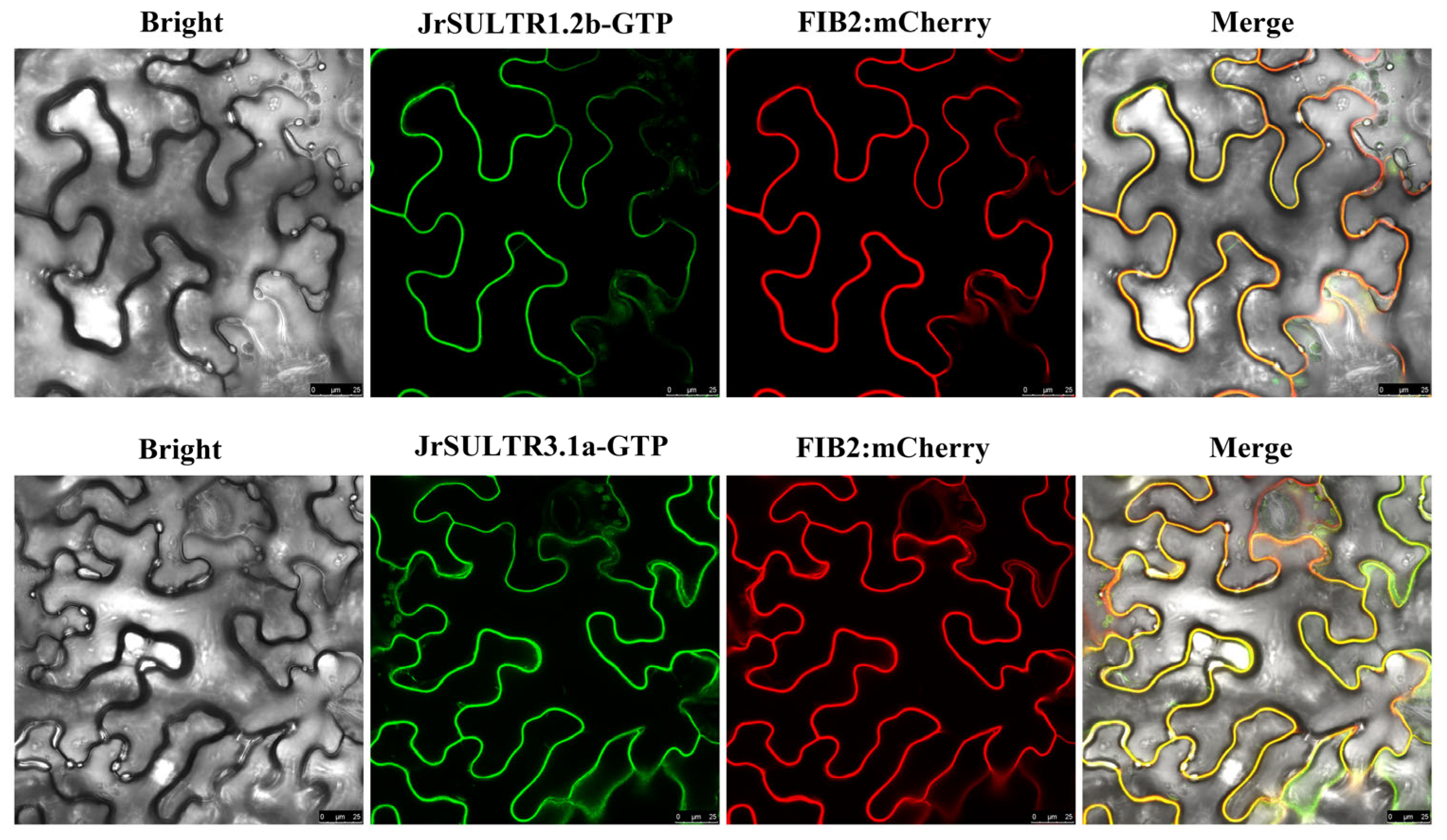
3.8. Subcellular Determination of JrSULTR1.2b and JrSULTR3.1a
4. Discussion
4.1. Functional Diversity and Physicochemical Properties of JrSULTR Genes
4.2. Phylogenetic Relationship and Functional Exploration of JrSULTR Genes
4.3. Expression Analysis of JrSULTR Genes to Selenium Stress
4.4. Response and Function Analysis of JrSULTR Genes to Drought, Low-Temperature, and High-Temperature Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, J.; Duan, X.; Yang, J.; Beeching, J.R.; Zhang, P. Coupled Expression of Cu/Zn-Superoxide Dismutase and Catalase in Cassava Improves Tolerance against Cold and Drought Stresses. Plant Signal. Behav. 2013, 8, e24525. [Google Scholar] [CrossRef] [PubMed]
- Buchner, P.; Stuiver, C.E.E.; Westerman, S.; Wirtz, M.; Hell, R.; Hawkesford, M.J.; De Kok, L.J. Regulation of Sulfate Uptake and Expression of Sulfate Transporter Genes in Brassica Oleracea as Affected by Atmospheric H(2)S and Pedospheric Sulfate Nutrition. Plant Physiol. 2004, 136, 3396–3408. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Watanabe-Takahashi, A.; Smith, F.W.; Blake-Kalff, M.; Hawkesford, M.J.; Saito, K. The Roles of Three Functional Sulphate Transporters Involved in Uptake and Translocation of Sulphate in Arabidopsis Thaliana. Plant J. 2000, 23, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Leggett, J.E.; Epstein, E. Kinetics of Sulfate Absorption by Barley Roots. Plant Physiol. 1956, 31, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Hawkesford, M.J.; Davidian, J.-C.; Grignon, C. Sulphate/Proton Cotransport in Plasma-Membrane Vesicles Isolated from Roots of Brassica napus L.: Increased Transport in Membranes Isolated from Sulphur-Starved Plants. Planta 1993, 190, 297–304. [Google Scholar] [CrossRef]
- Shibagaki, N.; Rose, A.; McDermott, J.P.; Fujiwara, T.; Hayashi, H.; Yoneyama, T.; Davies, J.P. Selenate-Resistant Mutants of Arabidopsis Thaliana Identify Sultr1;2, a Sulfate Transporter Required for Efficient Transport of Sulfate into Roots. Plant J. Cell Mol. Biol. 2002, 29, 475–486. [Google Scholar] [CrossRef]
- Chen, Z.; Zhao, P.-X.; Miao, Z.-Q.; Qi, G.-F.; Wang, Z.; Yuan, Y.; Ahmad, N.; Cao, M.-J.; Hell, R.; Wirtz, M.; et al. SULTR3s Function in Chloroplast Sulfate Uptake and Affect ABA Biosynthesis and the Stress Response. Plant Physiol. 2019, 180, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Akbudak, M.A.; Filiz, E.; Kontbay, K. Genome-Wide Identification and Cadmium Induced Expression Profiling of Sulfate Transporter (SULTR) Genes in Sorghum (Sorghum bicolor L.). Biometals Int. J. Role Met. Ions Biol. Biochem. Med. 2018, 31, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Buchner, P.; Parmar, S.; Kriegel, A.; Carpentier, M.; Hawkesford, M.J. The Sulfate Transporter Family in Wheat: Tissue-Specific Gene Expression in Relation to Nutrition. Mol. Plant 2010, 3, 374–389. [Google Scholar] [CrossRef] [PubMed]
- Xun, M.; Song, J.; Shi, J.; Li, J.; Shi, Y.; Yan, J.; Zhang, W.; Yang, H. Genome-Wide Identification of Sultr Genes in Malus Domestica and Low Sulfur-Induced MhSultr3;1a to Increase Cysteine-Improving Growth. Front. Plant Sci. 2021, 12, 748242. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.; Peng, C.; Quanle, X. Cloning and Expression Analysis of LsSULTR Gene in Lathyrus Sativus. Xibei Zhiwu Xuebao 2021, 41, 1954–1961. [Google Scholar]
- Zhang, H.; Hao, X.; Zhang, J.; Wang, L.; Wang, Y.; Li, N.; Guo, L.; Ren, H.; Zeng, J. Genome-Wide Identification of SULTR Genes in Tea Plant and Analysis of Their Expression in Response to Sulfur and Selenium. Protoplasma 2022, 259, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Adiputra, I.G.K.; Anderson, J.W. Distribution and Redistribution of Sulphur Taken up from Nutrient Solution during Vegetative Growth in Barley. Physiol. Plant. 1992, 85, 453–460. [Google Scholar] [CrossRef]
- Bolchi, A.; Petrucco, S.; Tenca, P.L.; Foroni, C.; Ottonello, S. Coordinate Modulation of Maize Sulfate Permease and ATP Sulfurylase mRNAs in Response to Variations in Sulfur Nutritional Status: Stereospecific down-Regulation by L-Cysteine. Plant Mol. Biol. 1999, 39, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Howarth, J.R.; Fourcroy, P.; Davidian, J.-C.; Smith, F.W.; Hawkesford, M.J. Cloning of Two Contrasting High-Affinity Sulfate Transporters from Tomato Induced by Low Sulfate and Infection by the Vascular Pathogen Verticillium Dahliae. Planta 2003, 218, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-W.; Xiang, C.-P.; Mei, S.-Y. Cloning and Characterization of a Novel Sulfate Transporter Gene from Radish (Raphanus Sativus L.). DNA Seq. J. DNA Seq. Mapp. 2006, 17, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, N.; Takahashi, H.; Smith, F.W.; Yamaya, T.; Saito, K. Two Distinct High-Affinity Sulfate Transporters with Different Inducibilities Mediate Uptake of Sulfate in Arabidopsis Roots. Plant J. 2002, 29, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, N.; Inoue, E.; Saito, K.; Yamaya, T.; Takahashi, H. Phloem-Localizing Sulfate Transporter, Sultr1;3, Mediates Re-Distribution of Sulfur from Source to Sink Organs in Arabidopsis. Plant Physiol. 2003, 131, 1511–1517. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, T.; Hayashi, N.; Yamaya, T.; Takahashi, H. Root-to-Shoot Transport of Sulfate in Arabidopsis. Evidence for the Role of SULTR3;5 as a Component of Low-Affinity Sulfate Transport System in the Root Vasculature. Plant Physiol. 2004, 136, 4198–4204. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.-J.; Wang, Z.; Wirtz, M.; Hell, R.; Oliver, D.J.; Xiang, C.-B. SULTR3;1 Is a Chloroplast-Localized Sulfate Transporter in Arabidopsis Thaliana. Plant J. 2013, 73, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant Drought Stress: Effects, Mechanisms and Management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Aslam, M.; Fakher, B.; Ashraf, M.A.; Cheng, Y.; Wang, B.; Qin, Y. Plant Low-Temperature Stress: Signaling and Response. Agronomy 2022, 12, 702. [Google Scholar] [CrossRef]
- Li, Z.; Cheng, B.; Zhao, Y.; Luo, L.; Zhang, Y.; Feng, G.; Han, L.; Peng, Y.; Zhang, X. Metabolic Regulation and Lipidomic Remodeling in Relation to Spermidine-Induced Stress Tolerance to High Temperature in Plants. Int. J. Mol. Sci. 2022, 23, 12247. [Google Scholar] [CrossRef]
- Gallardo, K.; Courty, P.-E.; Le Signor, C.; Wipf, D.; Vernoud, V. Sulfate Transporters in the Plant’s Response to Drought and Salinity: Regulation and Possible Functions. Front. Plant Sci. 2014, 5, 580. [Google Scholar] [CrossRef] [PubMed]
- El Kassis, E.; Cathala, N.; Rouached, H.; Fourcroy, P.; Berthomieu, P.; Terry, N.; Davidian, J.-C. Characterization of a Selenate-Resistant Arabidopsis Mutant. Root Growth as a Potential Target for Selenate Toxicity. Plant Physiol. 2007, 143, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Barberon, M.; Berthomieu, P.; Clairotte, M.; Shibagaki, N.; Davidian, J.-C.; Gosti, F. Unequal Functional Redundancy between the Two Arabidopsis Thaliana High-Affinity Sulphate Transporters SULTR1;1 and SULTR1;2. New Phytol. 2008, 180, 608–619. [Google Scholar] [CrossRef] [PubMed]
- Terry, N.; Zayed, A.M.; De Souza, M.P.; Tarun, A.S. SELENIUM IN HIGHER PLANTS. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 401–432. [Google Scholar] [CrossRef] [PubMed]
- Burnell, J.N. Selenium Metabolism in Neptunia Amplexicaulis1. Plant Physiol. 1981, 67, 316–324. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Bowen, H.C.; Parmaguru, P.; Fritz, M.; Spracklen, W.P.; Spiby, R.E.; Meacham, M.C.; Mead, A.; Harriman, M.; Trueman, L.J.; et al. Interactions between Selenium and Sulphur Nutrition in Arabidopsis Thaliana. J. Exp. Bot. 2004, 55, 1927–1937. [Google Scholar] [CrossRef]
- Sors, T.G.; Ellis, D.R.; Na, G.N.; Lahner, B.; Lee, S.; Leustek, T.; Pickering, I.J.; Salt, D.E. Analysis of Sulfur and Selenium Assimilation in Astragalus Plants with Varying Capacities to Accumulate Selenium. Plant J. 2005, 42, 785–797. [Google Scholar] [CrossRef] [PubMed]
- Sors, T.G.; Ellis, D.R.; Salt, D.E. Selenium Uptake, Translocation, Assimilation and Metabolic Fate in Plants. Photosynth. Res. 2005, 86, 373–389. [Google Scholar] [CrossRef] [PubMed]
- de Souza, G.A.; de Carvalho, J.G.; Rutzke, M.; Albrecht, J.C.; Guilherme, L.R.G.; Li, L. Evaluation of Germplasm Effect on Fe, Zn and Se Content in Wheat Seedlings. Plant Sci. Int. J. Exp. Plant Biol. 2013, 210, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Popescu, R.; Ionete, R.E.; Botoran, O.R.; Costinel, D.; Bucura, F.; Geana, E.I.; ’Alabedallat, Y.F.J.; Botu, M. 1H-NMR Profiling and Carbon Isotope Discrimination as Tools for the Comparative Assessment of Walnut (Juglans regia L.) Cultivars with Various Geographical and Genetic Origins-A Preliminary Study. Molecules 2019, 24, 1378. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Du, H.; Xue, Y.; Liao, Y.; Zhang, W.; Ye, J.; Wang, Q.; Xu, F. Genome-Wide Identification and Expression Analysis of the Walnut C-Repeat Binding Factor Gene Family under Low-Temperature Stress. Forests 2023, 14, 2274. [Google Scholar] [CrossRef]
- Poggi, V.; Arcioni, A.; Filippini, P.; Pifferi, P.G. Foliar Application of Selenite and Selenate to Potato (Solanum tuberosum): Effect of a Ligand Agent on Selenium Content of Tubers. J. Agric. Food Chem. 2000, 48, 4749–4751. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Chen, L.; Xu, J.; Zhang, Y.; Pan, G. Determination of Selenium Concentration in Rice and the Effect of Foliar Application of Se-Enriched Fertiliser or Sodium Selenite on the Selenium Content of Rice. J. Sci. Food Agric. 2002, 82, 869–872. [Google Scholar] [CrossRef]
- Yang, F.; Chen, L.; Hu, Q.; Pan, G. Effect of the Application of Selenium on Selenium Content of Soybean and Its Products. Biol. Trace Elem. Res. 2003, 93, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Slekovec, M.; Goessler, W. Accumulation of Selenium in Natural Plants and Selenium Supplemented Vegetable and Selenium Speciation by HPLC-ICPMS. Chem. Speciat. Bioavailab. 2005, 17, 63–73. [Google Scholar] [CrossRef]
- Smrkolj, P.; Stibilj, V.; Kreft, I.; Germ, M. Selenium Species in Buckwheat Cultivated with Foliar Addition of Se(VI) and Various Levels of UV-B Radiation. Food Chem. 2006, 96, 675–681. [Google Scholar] [CrossRef]
- Kápolna, E.; Hillestrøm, P.R.; Laursen, K.H.; Husted, S.; Larsen, E.H. Effect of Foliar Application of Selenium on Its Uptake and Speciation in Carrot. Food Chem. 2009, 115, 1357–1363. [Google Scholar] [CrossRef]
- Irish, K.P.; Harvey, M.-A.; Harris, H.H.; Aarts, M.G.M.; Chan, C.X.; Erskine, P.D.; van der Ent, A. Micro-Analytical and Molecular Approaches for Understanding the Distribution, Biochemistry, and Molecular Biology of Selenium in (Hyperaccumulator) Plants. PLANTA 2023, 257, 2. [Google Scholar] [CrossRef] [PubMed]
- Schiavon, M.; Pilon, M.; Malagoli, M.; Pilon-Smits, E.A.H. Exploring the Importance of Sulfate Transporters and ATP Sulphurylases for Selenium Hyperaccumulation—A Comparison of Stanleya Pinnata and Brassica Juncea (Brassicaceae). Front. Plant Sci. 2015, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Yu, T.; Cong, X.; Lai, X.; Xiang, J.; Cao, J.; Liao, X.; Gou, Y.; Chao, W.; Xue, H.; et al. Transcriptome, Proteome, and Metabolome Reveal the Mechanism of Tolerance to Selenate Toxicity in Cardamine Violifolia. J. Hazard. Mater. 2021, 406, 124283. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Guo, L.; Huang, J.; Hao, X.; Li, X.; Li, N.; Wang, Y.; Zhang, K.; Wang, X.; Wang, L.; et al. Comparative Transcriptomics Provides Novel Insights into the Mechanisms of Selenium Accumulation and Transportation in Tea Cultivars (Camellia sinensis (L.) O. Kuntze). Front. Plant Sci. 2023, 14, 1268537. [Google Scholar] [CrossRef] [PubMed]
- Heidari, P.; Hasanzadeh, S.; Faraji, S.; Ercisli, S.; Mora-Poblete, F. Genome-Wide Characterization of the Sulfate Transporter Gene Family in Oilseed Crops: Camelina Sativa and Brassica Napus. Plants 2023, 12, 628. [Google Scholar] [CrossRef]
- Yuan, Z.; Long, W.; Hu, H.; Liang, T.; Luo, X.; Hu, Z.; Zhu, R.; Wu, X. Genome-Wide Identification and Expansion Patterns of SULTR Gene Family in Gramineae Crops and Their Expression Profiles under Abiotic Stress in Oryza Sativa. Genes 2021, 12, 634. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Wang, M.; Xia, Z. The SULTR Gene Family in Maize (Zea Mays L.): Gene Cloning and Expression Analyses under Sulfate Starvation and Abiotic Stress. J. Plant Physiol. 2018, 220, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Geng, J.; Du, Y.; Li, S.; Yuan, X.; Zhu, J.; Zhou, Z.; Wang, Q.; Du, J. The Common Bean (Phaseolus vulgaris) SULTR Gene Family: Genome-Wide Identification, Phylogeny, Evolutionary Expansion and Expression Patterns. Biotechnol. Biotechnol. Equip. 2022, 36, 724–736. [Google Scholar] [CrossRef]
- Cao, M.-J.; Wang, Z.; Zhao, Q.; Mao, J.-L.; Speiser, A.; Wirtz, M.; Hell, R.; Zhu, J.-K.; Xiang, C.-B. Sulfate Availability Affects ABA Levels and Germination Response to ABA and Salt Stress in Arabidopsis Thaliana. Plant J. Cell Mol. Biol. 2014, 77, 604–615. [Google Scholar] [CrossRef] [PubMed]
- ARVY, M.P. Selenate and Selenite Uptake and Translocation in Bean Plants (Phaseolus vulgaris). J. Exp. Bot. 1993, 44, 1083–1087. [Google Scholar] [CrossRef]
- Ding, Y.; Zhou, X.; Zuo, L.; Wang, H.; Yu, D. Identification and Functional Characterization of the Sulfate Transporter Gene GmSULTR1;2b in Soybean. BMC Genom. 2016, 17, 373. [Google Scholar] [CrossRef] [PubMed]
- Rouached, H.; Wirtz, M.; Alary, R.; Hell, R.; Arpat, A.B.; Davidian, J.-C.; Fourcroy, P.; Berthomieu, P. Differential Regulation of the Expression of Two High-Affinity Sulfate Transporters, SULTR1.1 and SULTR1.2, in Arabidopsis. Plant Physiol. 2008, 147, 897–911. [Google Scholar] [CrossRef] [PubMed]
- Vatansever, R.; Koc, I.; Ozyigit, I.I.; Sen, U.; Uras, M.E.; Anjum, N.A.; Pereira, E.; Filiz, E. Genome-Wide Identification and Expression Analysis of Sulfate Transporter (SULTR) Genes in Potato (Solanum tuberosum L.). Planta 2016, 244, 1167–1183. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.R.; Nazir, F.; Asgher, M.; Per, T.S.; Khan, N.A. Selenium and Sulfur Influence Ethylene Formation and Alleviate Cadmium-Induced Oxidative Stress by Improving Proline and Glutathione Production in Wheat. J. Plant Physiol. 2015, 173, 9–18. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Amino Acid Number (aa) | Molecular Weight (kDa) | Theoretical pI | Hydrophilic | Predicted Localization |
|---|---|---|---|---|---|---|
| JreChr06G11410 | JrSULTR1.1a | 646 | 71.10 | 9.3 | 0.376 | Cell membrane |
| JreChr06G11416 | JrSULTR1.1b | 654 | 72.07 | 9.3 | 0.352 | Cell membrane |
| JreChr05G11871 | JrSULTR1.2a | 642 | 70.85 | 9.32 | 0.363 | Cell membrane |
| JreChr06G11781 | JrSULTR1.2b | 658 | 72.66 | 9.02 | 0.312 | Cell membrane |
| JreChr03G10342 | JrSULTR2.1 | 683 | 73.86 | 8.43 | 0.537 | Cell membrane |
| JreChr05G11869 | JrSULTR2.2 | 661 | 71.94 | 9.17 | 0.488 | Cell membrane |
| JreChr08G10436 | JrSULTR3.1a | 642 | 70.76 | 8.53 | 0.461 | Cell membrane |
| JreChr07G10637 | JrSULTR3.1b | 656 | 72.31 | 8.47 | 0.385 | Cell membrane |
| JreChr01G12210 | JrSULTR3.2a | 649 | 70.98 | 8.5 | 0.455 | Cell membrane |
| JreChr02G10983 | JrSULTR3.2b | 650 | 71.42 | 8.89 | 0.445 | Cell membrane |
| JreChr05G11381 | JrSULTR3.3 | 664 | 72.44 | 8.87 | 0.376 | Cell membrane |
| JreChr01G12720 | JrSULTR3.4a | 657 | 72.05 | 9.15 | 0.528 | Cell membrane |
| JreChr01G12712 | JrSULTR3.4b | 639 | 70.03 | 9.41 | 0.538 | Cell membrane |
| JreChr02G11304 | JrSULTR3.4c | 655 | 71.70 | 9.1 | 0.595 | Cell membrane |
| JreChr03G12216 | JrSULTR3.5a | 660 | 73.84 | 9.1 | 0.343 | Cell membrane |
| JreChr04G11407 | JrSULTR3.5b | 626 | 69.11 | 8.75 | 0.423 | Cell membrane |
| JreChr02G11621 | JrSULTR4.1 | 690 | 75.70 | 8.29 | 0.359 | Chloroplast |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Xue, Y.; Liu, N.; Quzhen, D.; Qiong, D.; Liao, Y.; Zhang, W.; Ye, J.; Wang, Q.; Xu, F. Isolation and Characterization of the Sulfate Transporter Gene Family and Its Expression Pattern in Response to Selenium and Abiotic Stress in Walnuts (Juglans regia L.). Forests 2024, 15, 702. https://doi.org/10.3390/f15040702
Zhang S, Xue Y, Liu N, Quzhen D, Qiong D, Liao Y, Zhang W, Ye J, Wang Q, Xu F. Isolation and Characterization of the Sulfate Transporter Gene Family and Its Expression Pattern in Response to Selenium and Abiotic Stress in Walnuts (Juglans regia L.). Forests. 2024; 15(4):702. https://doi.org/10.3390/f15040702
Chicago/Turabian StyleZhang, Simin, Yansheng Xue, Ningfang Liu, Danzeng Quzhen, De Qiong, Yongling Liao, Weiwei Zhang, Jiabao Ye, Qijian Wang, and Feng Xu. 2024. "Isolation and Characterization of the Sulfate Transporter Gene Family and Its Expression Pattern in Response to Selenium and Abiotic Stress in Walnuts (Juglans regia L.)" Forests 15, no. 4: 702. https://doi.org/10.3390/f15040702
APA StyleZhang, S., Xue, Y., Liu, N., Quzhen, D., Qiong, D., Liao, Y., Zhang, W., Ye, J., Wang, Q., & Xu, F. (2024). Isolation and Characterization of the Sulfate Transporter Gene Family and Its Expression Pattern in Response to Selenium and Abiotic Stress in Walnuts (Juglans regia L.). Forests, 15(4), 702. https://doi.org/10.3390/f15040702