
Extract of Calyces from Physalis peruviana Reduces Insulin Resistance and Oxidative Stress in Streptozotocin-Induced Diabetic Mice

 , , ,
, , ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Plant Material
Preparation of the Extract and Fractions
2.3. Dereplication Analysis
2.4. Animals
2.5. Diabetic Chronic Model Induction and Treatments
2.5.1. Oral Glucose Tolerance Test (OGTT)
2.5.2. HOMA-IR
2.6. Biochemical Parameters
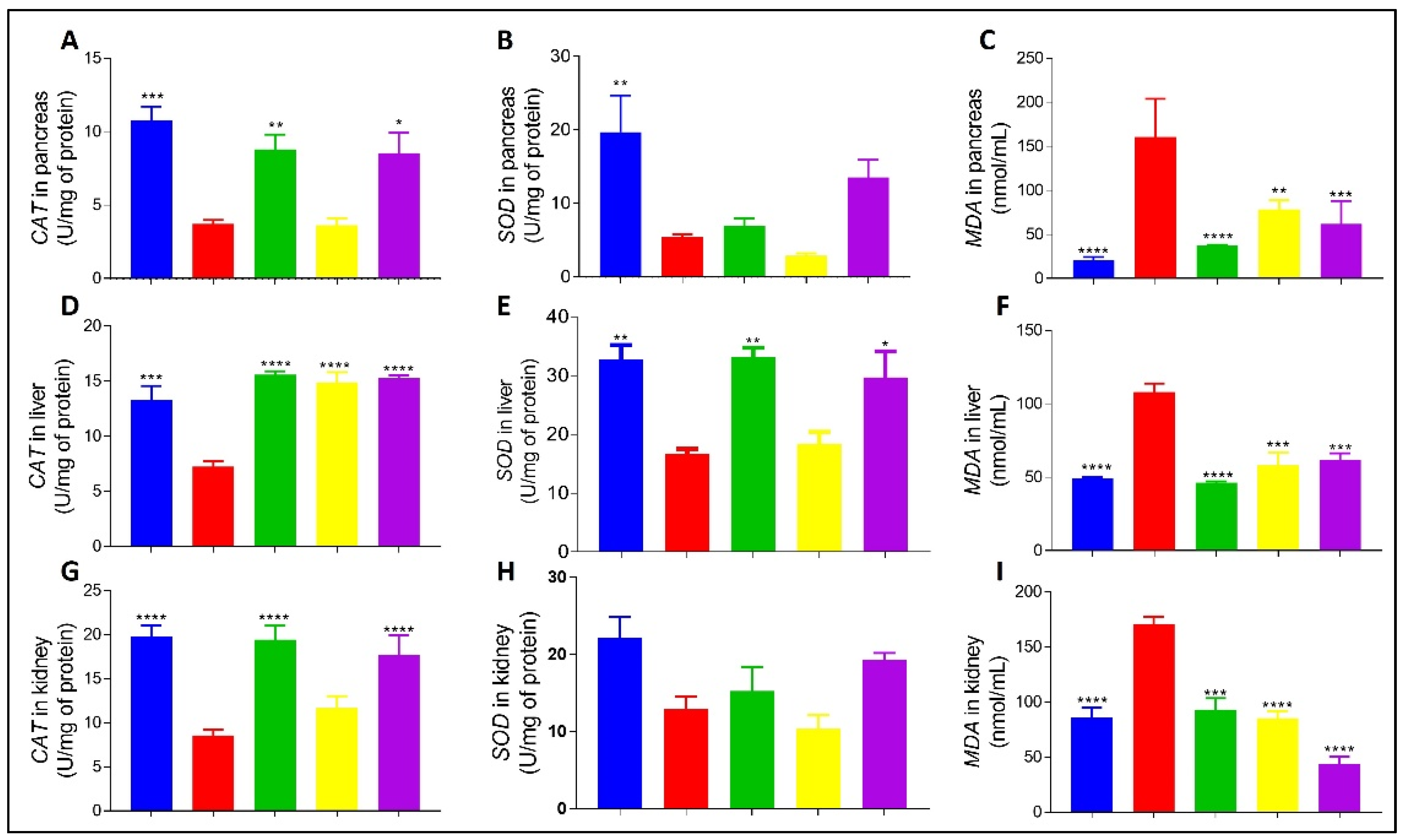
2.6.1. Oxidative Stress Parameters
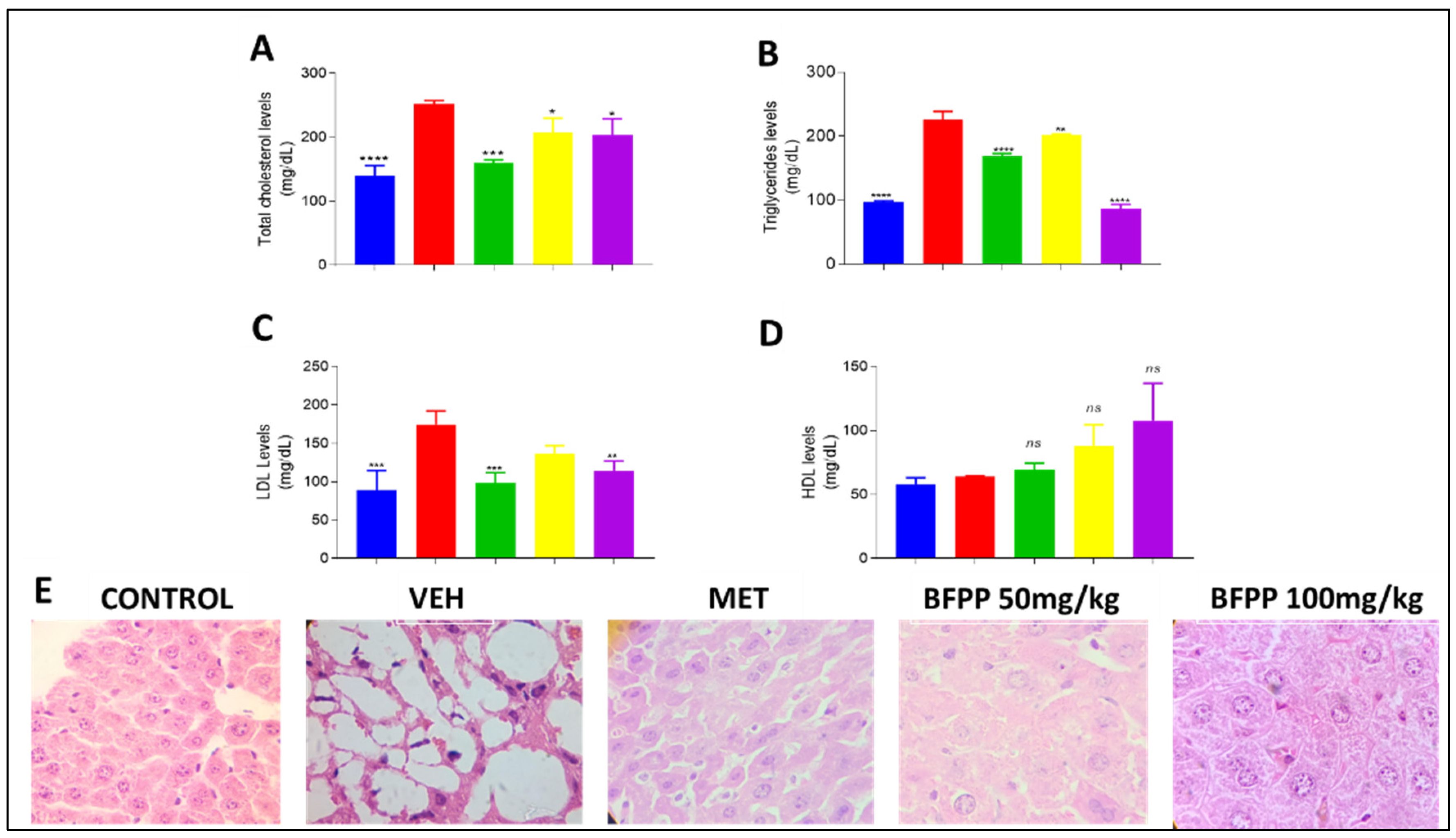
2.6.2. Serum Lipid Profile
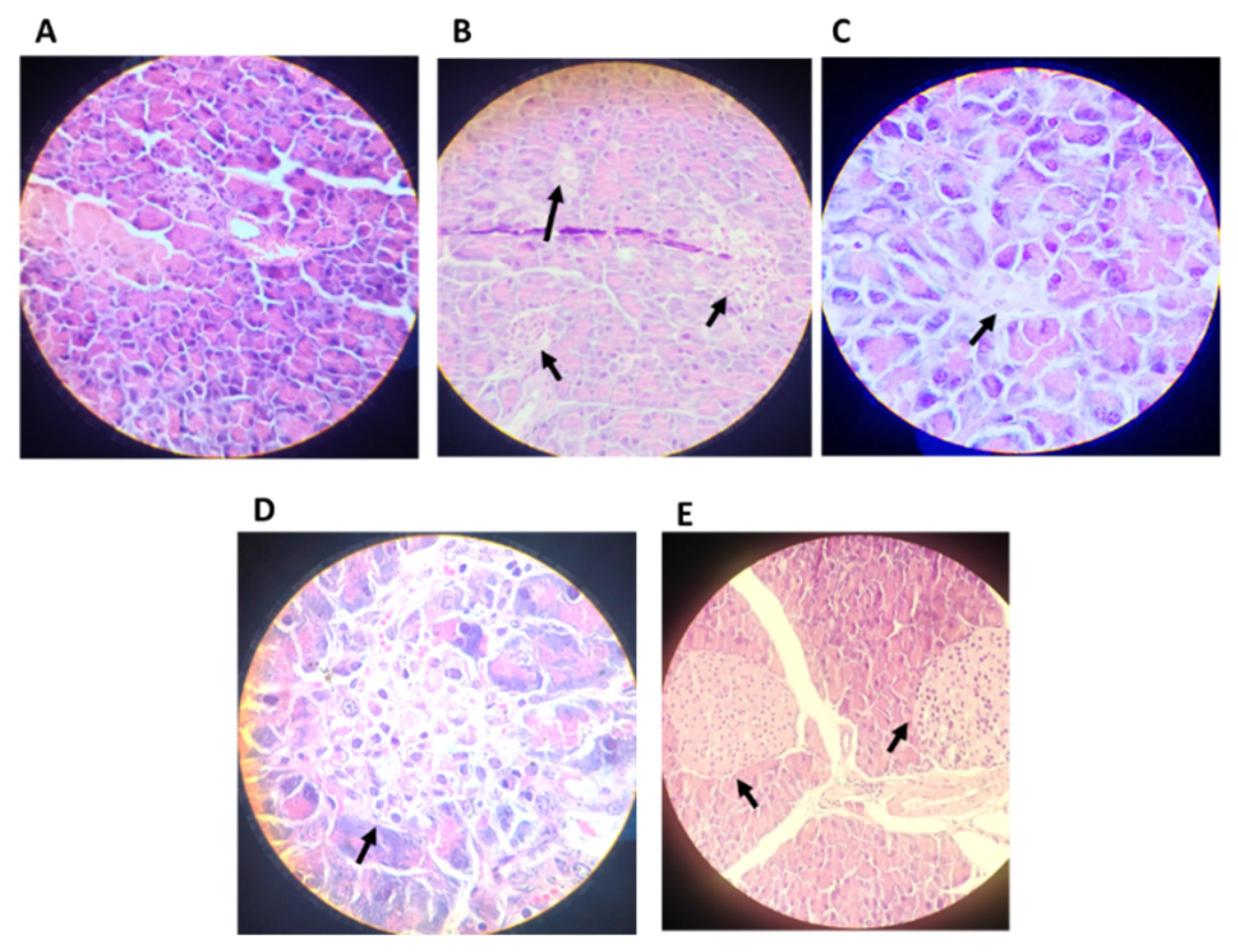
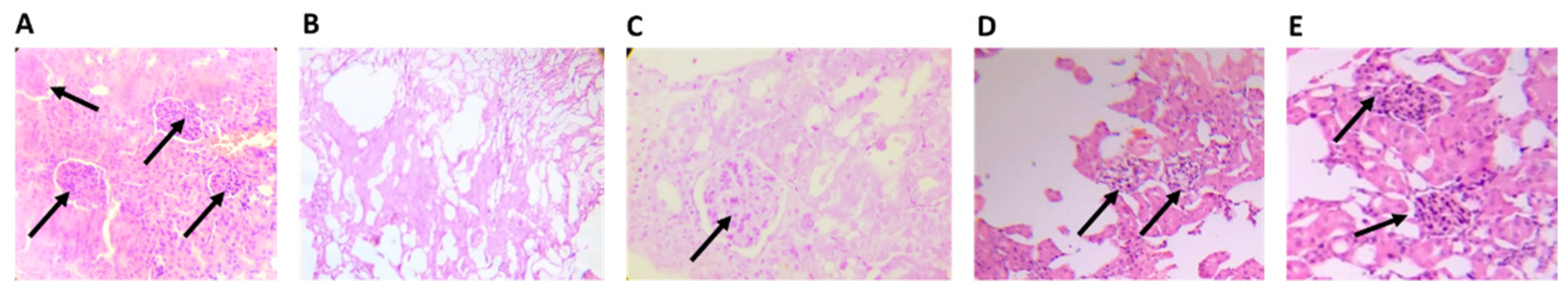
2.7. Histological Analysis
3. Statistical Analysis
4. Results
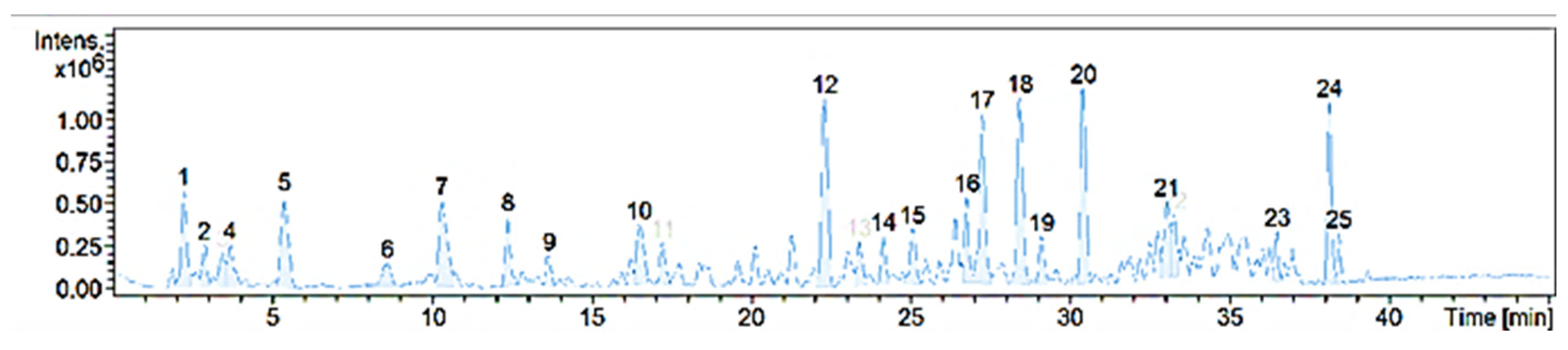
4.1. Chemical Characterization
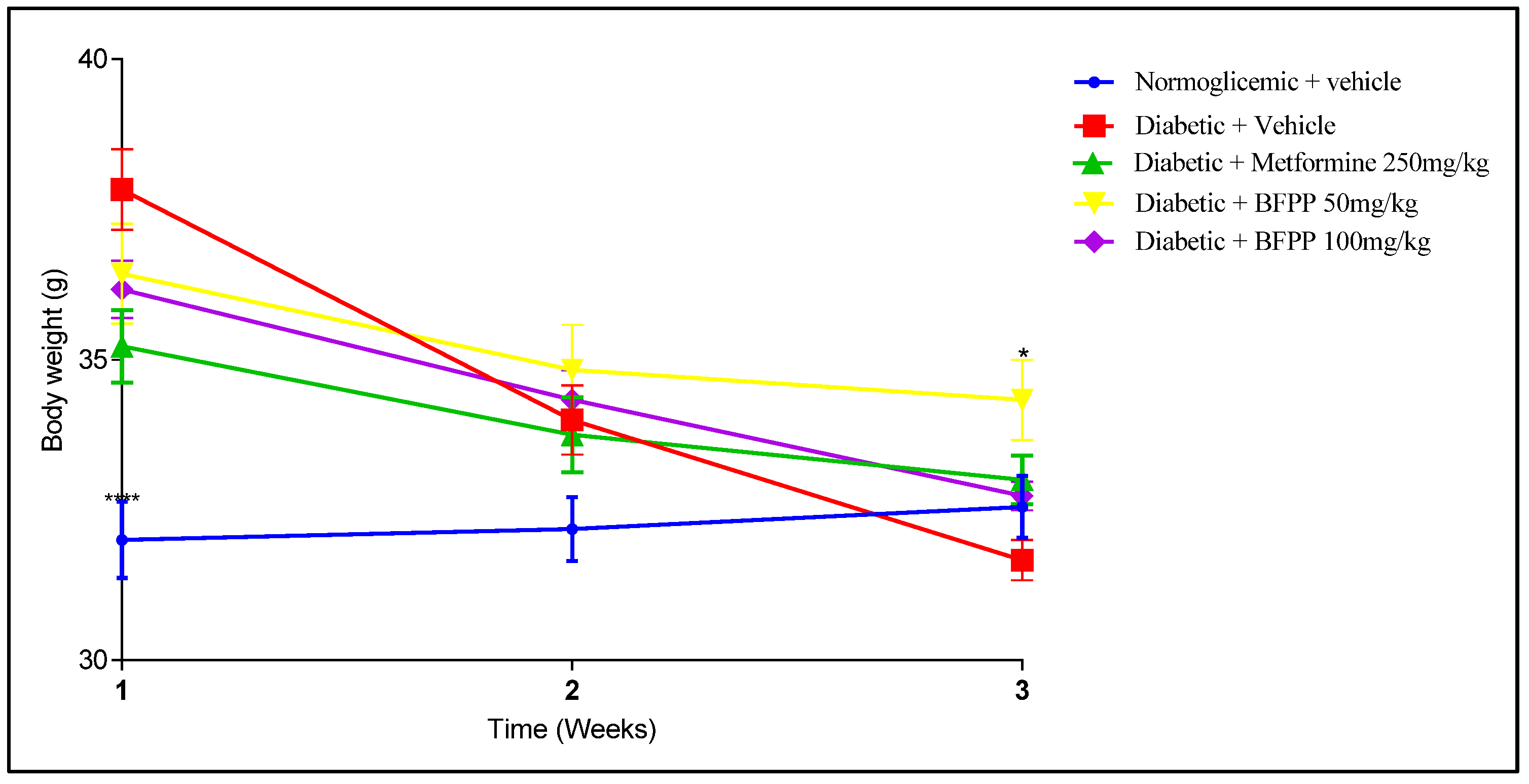
4.2. Diabetic Model
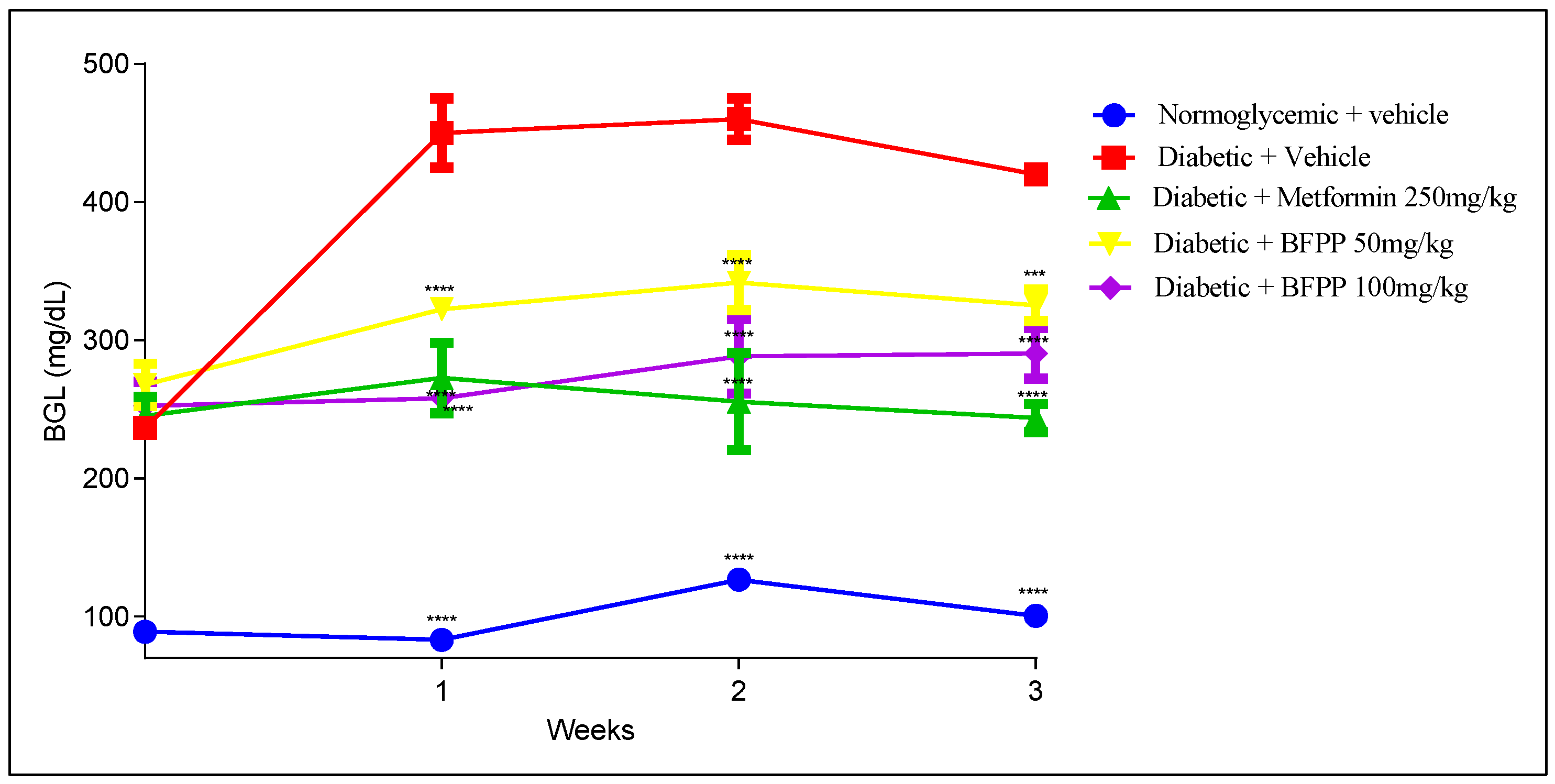
4.2.1. Effect of Butanol Fraction from P. peruviana on Blood Glucose Levels
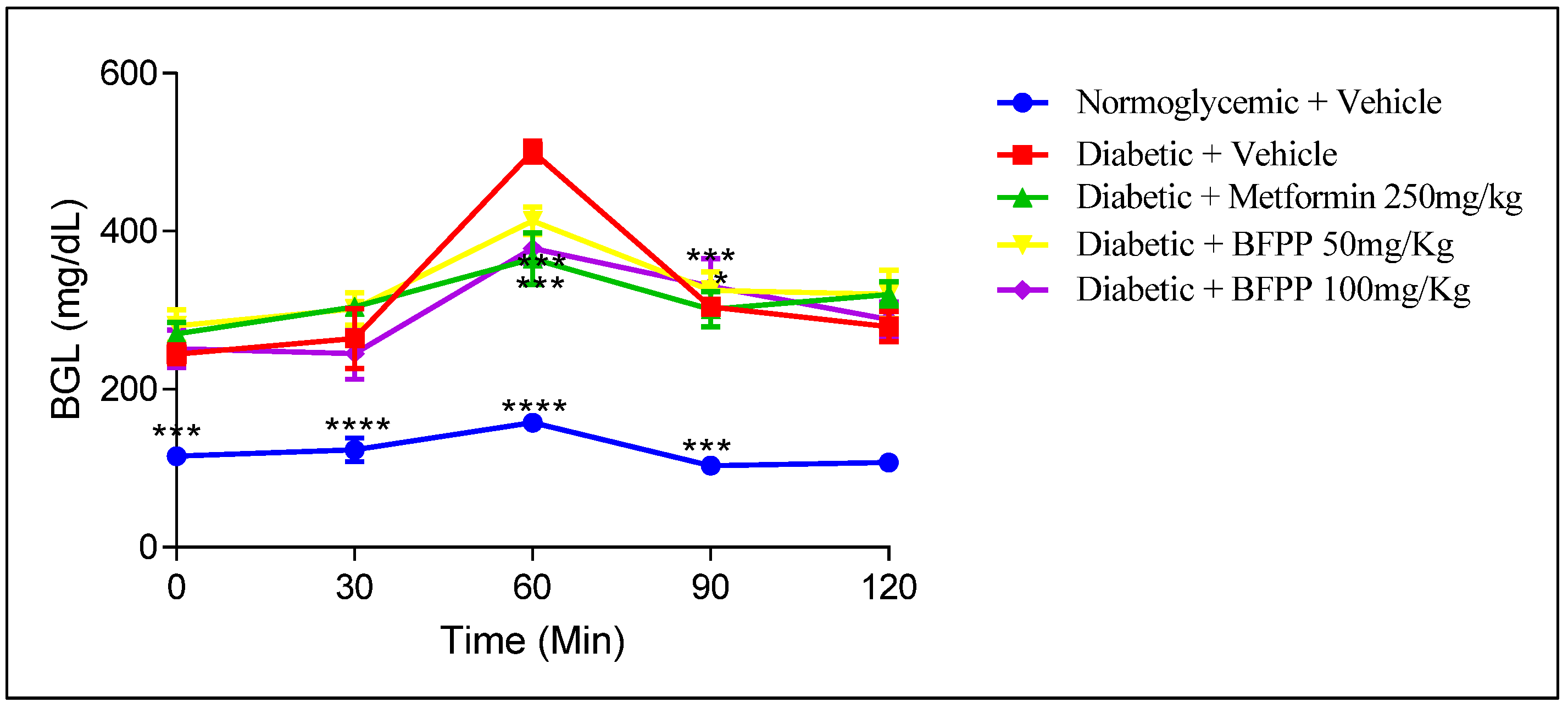
4.2.2. Oral Glucose Tolerance Test (OGTT)
4.2.3. Homa-IR
4.2.4. Effect of BFPP on Lipid Profile
4.2.5. Effect of BFPP on Oxidative Stress Markers
4.2.6. Histopathological Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Fischer, G.; Melgarejo, L.U.Z.M. The ecophysiology of cape gooseberry (Physalis peruviana L.)—An Andean fruit crop. A review. La ecofisiología de uchuva (Physalis peruviana L.)—Un frutal andino. Una revisión. Rev. Colomb. Cienc. Hortíc. 2020, 14, 76–89. [Google Scholar]
- Granados Pérez, W.; Muñoz Vanegas, C.A.; Aguillón Mayorga, D.M. Cadena de la uchuva. Dir Cadenas Agrícolas y For [Internet]. 2019, pp. 1–18. Available online: https://bit.ly/3Ik0gE2 (accessed on 1 December 2022).
- Saltveit, M.E. Postharvest Biology and Technology of Tropical and Subtropical Fruits; Woodhead Publishing: Sawston, UK, 2011. [Google Scholar] [CrossRef]
- Fischer, G.; Almanza-Merchán, P.J.; Miranda, D. Importancia y cultivo de la uchuva (Physalis peruviana L.). Rev. Bras. Frutic. 2014, 36, 1–15. [Google Scholar] [CrossRef]
- Kasali, F.M.; Tusiimire, J.; Kadima, J.N.; Tolo, C.U.; Weisheit, A.; Agaba, A.G. Ethnotherapeutic Uses and Phytochemical Composition of Physalis peruviana L.: An Overview. Sci. World J. 2021, 2021, 5212348. [Google Scholar] [CrossRef] [PubMed]
- Mora, Á.C.; Aragón, D.M.; Ospina, L.F. Effects of Physalis peruviana fruit extract on stress oxidative parameters in streptozotocin-diabetic rats. Lat. Am. J. Pharm. 2010, 29, 1132–1136. [Google Scholar]
- Rey, D.P.; Ospina, L.F.; Aragón, D.M. Inhibitory effects of an extract of fruits of Physalis peruviana on some intestinal carbohydrases. Rev. Colomb. Cienc. Quim.-Farm. 2015, 44, 72–89. [Google Scholar] [CrossRef]
- Wu, S.J.; Tsai, J.Y.; Chang, S.P.; Lin, D.L.; Wang, S.S.; Huang, S.N.; Ng, L.T. Supercritical carbon dioxide extract exhibits enhanced antioxidant and anti-inflammatory activities of Physalis peruviana. J. Ethnopharmacol. 2006, 108, 407–413. [Google Scholar] [CrossRef]
- Franco, L.A.; Ocampo, Y.C.; Gómez, H.A.; de la Puerta, R.; Espartero, J.L.; Ospina, L.F. Sucrose esters from Physalis peruviana calyces with anti-inflammatory activity. Planta Med. 2014, 80, 1605–1614. [Google Scholar] [CrossRef]
- Toro, R.M.; Aragón, D.M.; Ospina, L.F.; Ramos, F.A.; Castellanos, L. Phytochemical analysis, antioxidant and anti-inflammatory activity of calyces from Physalis peruviana. Nat. Prod. Commun. 2014, 9, 1573–1575. [Google Scholar] [CrossRef] [PubMed]
- Toro, R.; Arangon, M. Propuesta de un Marcador Analítico Como Herramienta en la Microencapsulación de un Extracto con Actividad Antioxidante de Cálices de Physalis peruviana. Master’s Thesis, Universidad Nacional de Colombia, Bogota, Colombia, 2014. [Google Scholar]
- Gutierres, J.M.; Lencina, C.L.; Carvalho, F.B.; Soares, M.S.P.; Bona, N.P.; Stefanello, F.M.; Vieira, A.; Chaves, V.C.; Spanevello, R.M.; Reginatto, F.H.; et al. Southern Brazilian native fruit shows neurochemical, metabolic and behavioral benefits in an animal model of metabolic syndrome. Metab. Brain Dis. 2018, 33, 1551–1562. [Google Scholar] [CrossRef]
- Rendon, L.M.; Zuluaga, A.F.; Rodríguez, C.A.; Agudelo, M.; Vesga, O.; Ospina, L.F. Obtaining a Murine Model of Streptozotocin-Induced Diabetes Useful in the Pharmacodynamic Evaluation of Regular Insulin. Biomedica 2017, 2, 11–18. [Google Scholar] [CrossRef]
- Cardona, M.I.; Toro, R.M.; Costa, G.M.; Ospina, L.F.; Castellanos, L.; Ramos, F.A.; Aragón, D.M. Influence of extraction process on antioxidant activity and rutin content in Physalis peruviana calyces extract. J. Appl. Pharm. Sci. 2017, 7, 164–168. [Google Scholar] [CrossRef][Green Version]
- Costa, G.M. Vitexin Derivatives as Chemical Markers in the Differentiation of the Closely Related Species Passiflora quadrangularis Linn. J. Liq. Chromatogr. 2013, 36, 37–41. [Google Scholar]
- Miranda, C.A.; Schönholzer, T.E.; Klöppel, E.; Sinzato, Y.K.; Volpato, G.T.; Damasceno, D.C.; Campos, E.K. Repercussions of low fructose-drinking water in male rats. An. Acad. Bras. Cienc. 2019, 91, e20170705. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Zhao, P.; Pang, K.; Ma, Y.; Huang, H.; Zhou, T.; Yang, X. Antidiabetic effect of a flavonoid-rich extract from Sophora alopecuroides L. in HFD- and STZ-induced diabetic mice through PKC/GLUT4 pathway and regulating PPARα and PPARγ expression. J. Ethnopharmacol. 2021, 268, 113654. [Google Scholar] [CrossRef]
- Furman, B.L. Streptozotocin-Induced Diabetic Models in Mice and Rats. Curr. Protoc. Pharmacol. 2015, 70, 1–20. [Google Scholar] [CrossRef]
- Janssen, B.J.A.; de Celle, T.; Debets, J.J.M.; Brouns, A.E.; Callahan, M.F.; Smith, T.L. Effects of anesthetics on systemic hemodynamics in mice. Am. J. Physiol.—Heart Circ. Physiol. 2004, 287, 1618–1625. [Google Scholar] [CrossRef]
- Aragón, D.M.; Echeverry, S.M.; Valderrama, I.H.; Costa, G.M.; Ospina, L.F. Development and optimization of microparticles containing a hypoglycemic fraction of calyces from Physalis peruviana. J. Appl. Pharm. Sci. 2018, 8, 10–18. [Google Scholar] [CrossRef]
- Abdelhameed, R.F.A.; Ibrahim, A.K.; Elfaky, M.A.; Habib, E.S.; Mahamed, M.I.; Mehanna, E.T.; Darwish, K.M.; Khodeer, D.M.; Ahmed, S.A.; Elhady, S.S. Antioxidant and anti-inflammatory activity of Cynanchum acutum L. isolated flavonoids using experimentally induced type 2 diabetes mellitus: Biological and in silico investigation for nf-κb pathway/mir-146a expression modulation. Antioxidants 2021, 10, 1713. [Google Scholar] [CrossRef]
- Abu-Amsha, R.; Croft, K.D.; Puddey, I.B.; Proudfoot, J.M.; Beilin, L.J. Phenolic content of various beverages determines the extent of inhibition of human serum and low-density lipoprotein oxidation in vitro: Identification and mechanism of action of some cinnamic acid derivatives from red wine. Clin. Sci. 1996, 91, 449–458. [Google Scholar] [CrossRef]
- Ou, L.; Kong, L.Y.; Zhang, X.M.; Niwa, M. Oxidation of ferulic acid by Momordica charantia peroxidase and related anti-inflammation activity changes. Biol. Pharm. Bull. 2003, 26, 1511–1516. [Google Scholar] [CrossRef]
- Kolodziejczyk-Czepas, J.; Kozachok, S.; Pecio, Ł.; Marchyshyn, S.; Oleszek, W. Determination of phenolic profiles of Herniaria polygama and Herniaria incana fractions and their in vitro antioxidant and anti-inflammatory effects. Phytochemistry 2021, 190, 112861. [Google Scholar] [CrossRef]
- Gironés-Vilaplana, A.; Baenas, N.; Villaño, D.; Speisky, H.; García-Viguera, C.; Moreno, D.A. Evaluation of Latin-American fruits rich in phytochemicals with biological effects. J. Funct. Foods 2014, 7, 599–608. [Google Scholar] [CrossRef]
- Medina, S.; Collado-gonzález, J.; Londoño-londoño, J.; Gil-izquierdo, A. Potential of Physalis peruviana calyces as a low-cost valuable resource of phytoprostanes and phenolic compounds. J. Sci. Food Agric. 2019, 99, 2194–2204. [Google Scholar] [CrossRef]
- Franco, L.A.O.L.; Matiz, G.E.; Calle, J.; Pinzón, R. Actividad antinflamatoria de extractos y fracciones obtenidas de cálices de Physalis peruviana L. Luis. Biomedica 2007, 27, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Manzano, S.; Williamson, G. Polyphenols and phenolic acids from strawberry and apple decrease glucose uptake and transport by human intestinal Caco-2 cells. Mol. Nutr. Food Res. 2010, 54, 1773–1780. [Google Scholar] [CrossRef]
- Liu, Z.; Hu, M. Natural polyphenol disposition via coupled metabolic pathways. Expert Opin. Drug Metab. Toxicol. 2007, 3, 389–406. [Google Scholar] [CrossRef]
- Moré, G.P.D.; Cardona, M.I.; Sepúlveda, P.M.; Echeverry, S.M.; Simões, C.M.O.; Aragón, D.M. Matrix effects of the hydroethanolic extract of calyces of Physalis peruviana L. On rutin pharmacokinetics in wistar rats using population modeling. Pharmaceutics 2021, 13, 535. [Google Scholar] [CrossRef] [PubMed]
- Monzón Daza, G.; Meneses Macías, C.; Forero, A.M.; Rodríguez, J.; Aragón, M.; Jiménez, C.; Ramos, F.A.; Castellanos, L. Identification of α-Amylase and α-Glucosidase Inhibitors and Ligularoside A, a New Triterpenoid Saponin from Passiflora ligularis Juss (Sweet Granadilla) Leaves, by a Nuclear Magnetic Resonance-Based Metabolomic Study. J. Agric. Food Chem. 2021, 69, 2919–2931. [Google Scholar] [CrossRef]
- Novoa, D.M.A.; Giraldo, L.F.O.; Rodríguez, F.A.R.; Hernández, L.C.; Modesti, G.C.; Silva, F.R.M.B. Passiflora ligularis Juss. (Granadilla): Farmacológicos de Una Estudios Químicos y Planta con Potencial Terapéutico; Coordinaci, Universidad Nacional de Colombia: Bogotá, Colombia, 2021. [Google Scholar] [CrossRef]
- Kappel, V.D.; Cazarolli, L.H.; Pereira, D.F.; Postal, B.G.; Madoglio, F.A.; Buss, Z.d.; Reginatto, F.H.; Silva, F.R.M.B. Beneficial effects of banana leaves (Musa x paradisiaca) on glucose homeostasis: Multiple sites of action. Braz. J. Pharmacogn. 2013, 23, 706–715. [Google Scholar] [CrossRef]
- Wang, Y.; Alkhalidy, H.; Liu, D. The emerging role of polyphenols in the management of type 2 diabetes. Molecules 2021, 26, 703. [Google Scholar] [CrossRef] [PubMed]
- Qu, H.-Q.; Li, Q.; Rentfro, A.R.; Fisher-Hoch, S.P.; Mccormick, J.B. The Definition of Insulin Resistance Using HOMA-IR for Americans of Mexican Descent Using Machine Learning. PLoS ONE 2011, 6, e21041. [Google Scholar] [CrossRef]
- Antunes, L.C.; Elkfury, J.L.; Jornada, M.N.; Foletto, K.C.; Bertoluci, M.C. Validation of HOMA-IR in a model of insulin-resistance induced by a high-fat diet in Wistar rats. Arch. Endocrinol. Metab. 2016, 60, 138–142. [Google Scholar] [CrossRef]
- Mather, K.J.; Verma, S.; Anderson, T.J. Improved endothelial function with metformin in type 2 diabetes mellitus. J. Am. Coll. Cardiol. 2001, 37, 1344–1350. [Google Scholar] [CrossRef]
- Assadi, S.; Shafiee, S.M.; Erfani, M.; Akmali, M. Antioxidative and antidiabetic effects of Capparis spinosa fruit extract on high-fat diet and low-dose streptozotocin-induced type 2 diabetic rats. Biomed. Pharmacother. 2021, 138, 111391. [Google Scholar] [CrossRef]
- Ji, S.; Zhu, C.; Gao, S.; Shao, X.; Chen, X.; Zhang, H.; Tang, D. Morus alba leaves ethanol extract protects pancreatic islet cells against dysfunction and death by inducing autophagy in type 2 diabetes. Phytomedicine 2021, 83, 153478. [Google Scholar] [CrossRef]
- Li, T.; Chen, S.; Feng, T.; Dong, J.; Li, Y.; Li, H. Rutin protects against aging-related metabolic dysfunction. Food Funct. 2016, 7, 1147–1154. [Google Scholar] [CrossRef]
- Imran, A.; Butt, M.S.; Arshad, M.S.; Arshad, M.U.; Saeed, F.; Sohaib, M.; Munir, R. Exploring the potential of black tea based flavonoids against hyperlipidemia related disorders. Lipids Health Dis. 2018, 17, 57. [Google Scholar] [CrossRef] [PubMed]
- De Paiva, L.B.; Goldbeck, R.; Santos, W.D.d.; Squina, F.M. Ferulic acid and derivatives: Molecules with potential application in the pharmaceutical field. Braz. J. Pharm. Sci. 2013, 49, 395–411. [Google Scholar] [CrossRef]
- Tariq, S.; Imran, M.; Mushtaq, Z.; Asghar, N. Phytopreventive antihypercholesterolmic and antilipidemic perspectives of zedoary (Curcuma zedoaria Roscoe.) herbal tea. Lipids Health Dis. 2016, 15, 39. [Google Scholar] [CrossRef]
- Goss, M.J.; Nunes, M.L.O.; Machado, I.D.; Merlin, L.; Macedo, N.B.; Silva, A.M.O.; Bresolin, T.M.B.; Santin, J.R. Peel flour of Passiflora edulis Var. Flavicarpa supplementation prevents the insulin resistance and hepatic steatosis induced by low-fructose-diet in young rats. Biomed. Pharmacother. 2018, 102, 848–854. [Google Scholar] [CrossRef]
- Albu, E.; Lupaşcu, D.; Filip, C.; Jaba, I.M.; Zamosteanu, N. The influence of a new rutin derivative on homocysteine, cholesterol and total antioxidative status in experimental diabetes in rat. Farmacia 2013, 61, 1167–1177. Available online: https://pdfs.semanticscholar.org/2e5c/6e7f3fde85fcc7591dc13e4f1ab8604e731a.pdf (accessed on 12 March 2018).
- Ramadan, M.F. Physalis peruviana pomace suppresses high-cholesterol diet-induced hypercholesterolemia in rats. Grasas Aceites 2012, 63, 411–422. [Google Scholar] [CrossRef]
- Darwish, A.G.; Mahmoud, H.I.; Refaat, I. Antioxidative and Antidiabetic Effect of Goldenberries juice and pomace on Experimental Rats Induced with streptozotocin In Vitro. J. Food Dairy Sci. 2020, 11, 277–283. [Google Scholar] [CrossRef]
- Mansuroǧlu, B.; Derman, S.; Yaba, A.; Kizilbey, K. Protective effect of chemically modified SOD on lipid peroxidation and antioxidant status in diabetic rats. Int. J. Biol. Macromol. 2015, 72, 79–87. [Google Scholar] [CrossRef]
- Lenzen, S. The mechanisms of alloxan- and streptozotocin-induced diabetes. Diabetologia 2008, 51, 216–226. [Google Scholar] [CrossRef]
- Toro, A.R.M.; Aragón, N.D.M.; Ospina, G.L.F. Hepatoprotective effect of calyces extract of Physalis peruviana on hepatotoxicity induced by CCl4 in wistar rats. Vitae 2013, 20, 125–132. [Google Scholar]
- Lenzen, S.; Drinkgern, J.; Tiedge, M. Low antioxidant enzyme gene expression in pancreatic islets compared with various other mouse tissues. Free Radic. Biol. Med. 1996, 20, 463–466. [Google Scholar] [CrossRef]
- Ezzat, S.M.; Abdallah, H.M.I.; Yassen, N.N.; Radwan, R.A.; Mostafa, E.S.; Salama, M.M.; Salem, M.A. Phenolics from Physalis peruviana fruits ameliorate streptozotocin-induced diabetes and diabetic nephropathy in rats via induction of autophagy and apoptosis regression. Biomed. Pharmacother. 2021, 142, 111948. [Google Scholar] [CrossRef]
- Yang, Y.S.; Huang, C.N.; Wang, C.J.; Lee, Y.J.; Chen, M.L.; Peng, C.H. Polyphenols of Hibiscus sabdariffa improved diabetic nephropathy via regulating the pathogenic markers and kidney functions of type 2 diabetic rats. J. Funct. Foods 2013, 5, 810–819. [Google Scholar] [CrossRef]
- Lan, Y.H.; Chang, F.R.; Pan, M.J.; Wu, C.C.; Wu, S.J.; Chen, S.L.; Wang, S.S.; Wu, M.J.; Wu, Y.C. New cytotoxic withanolides from Physalis peruviana. Food Chem. 2009, 116, 462–469. [Google Scholar] [CrossRef]
- Devkar, S.T.; Muthal, A.P.; Patil, P.V.; Mukherjee-Kandhare, A.A.; Kandhare, A.D.; Jagtap, S.D.; Bodhankar, S.L.; Hegde, M.V. Evaluation of the physicochemical stability and biological activity of withanolide rich fraction from withania somnifera root by hptlc and cyclic voltammetry: A simple, reliable, and cost-effective approach. Lat. Am. J. Pharm. 2021, 40, 946–956. [Google Scholar]
- Bharti, S.K.; Krishnan, S.; Kumar, A.; Kumar, A. Antidiabetic phytoconstituents and their mode of action on metabolic pathways. Ther. Adv. Endocrinol. Metab. 2018, 9, 81–100. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Max. m/z | RT. | Area | Possible Compound | Possible ID (CAS.) |
|---|---|---|---|---|---|
| 1 | 116.0708 | 2.2 | 6680138 | Amino Acid: L-Proline | 147-85-3 |
| 2 | 142.1230 | 2.9 | 2810985 | Tropane Alkaloids: | |
| (1) Tropine | 120-29-6 | ||||
| (2) Physoperuvine | 60723-27-5 | ||||
| (3) Cycloheptanone, 3-(methylamino) | 73744-99-7 | ||||
| 3 | 132.1024 | 3.4 | 2759332 | Amino Acid: L-Isoleucine | 61-90-5 |
| 4 | 132.1024 | 3.7 | 3675771 | Amino Acid: S-Leucine | 73-32-5 |
| 5 | 166.0868 | 5.4 | 8761530 | Amino Acid: Phenylalanine | 63-91-2 |
| 6 | 251.1399 | 8.6 | 2877906 | Cinnamic Acid Derivatives: Cinnamamide, N-(4-aminobutyl)-3,4-dihydroxy- | 8CI; 29554-26-5 |
| 7 | 205.0979 | 10.3 | 9139461 | Amino Acid: L-Tryptophan | 73-22-3 |
| 8 | 265.1555 | 12.4 | 45393229 | Cinnamic Acid Derivates: Cinnamamide, N-(4-aminobutyl)-4-hydroxy-3-methoxy- | 7CI,8CI; 501-13-3 |
| 9 | 474.2610 | 13.7 | 2022964 | N/I | N/I |
| 10 | 611.1622 | 16.5 | 5663280 | Flavanoid: Flavone, 2′,3,4′,5,7-pentahydroxy- | 8CI; 480-16-0 |
| 11 | 1037.5117 | 17.2 | 2652622 | Withanolides: Withaperuvin E | 92125-38-7 |
| 12 | 1005.5214 | 22.3 | 14971186 | Withanolides: Withaperuvin M | 1353093-20-5 |
| Phyperunolide A | 1198400-48-4 | ||||
| 13 | 233.1023 | 23.4 | 3245775 | N/I | N/I |
| 14 | 331.0817 | 24.1 | 2913771 | Terpene: Dihydroactinidiolide | 17092-92-1 |
| 15 | 247.1182 | 25.1 | 4087021 | N/I | N/I |
| 16 | 977.5637 | 26.8 | 4565672 | Withanolides: | |
| (1) Withaferin A | 5119-48-2 | ||||
| (2) Withanone | 7CI; 27570-38-3 | ||||
| (3) Withanolide D | 30655-48-2 | ||||
| (4) 27-Hydroxywithanolide B | 60124-17-6 | ||||
| 17 | 203.1797 | 27.2 | 12844741 | Monoterpene: Calamenene | 73209-42-4 |
| 18 | 345.0974 | 28.4 | 14689013 | Retinol | 68-26-8 |
| 19 | 316.2853 | 29.1 | 3375421 | Monoterpene: β-Vetivenene | 27840-40-0 |
| 20 | 345.2429 | 30.4 | 13726374 | N/I | N/I |
| 21 | 289.2535 | 33.0 | 5550203 | Biotin | 58-85-5 |
| 22 | 289.2533 | 33.2 | 3910919 | Fatty Acid: Linolenic acid, ethyl ester | 1783-84-2 |
| 23 | 273.2581 | 36.5 | 2264958 | Flavanoid: Flavanone, 4′,5,7-trihydroxy- | 8CI; 480-41-1 |
| 24 | 621.3086 | 38.1 | 10006030 | N/I | N/I |
| 25 | 621.3088 | 38.4 | 2128388 | N/I | N/I |
| Compounds | Rt | [M-H]− m/z | MSn | Hexose (-162) | Aglicone | Author |
|---|---|---|---|---|---|---|
| Quercetin 7-O-glucoside 3-O-rutinoside | 4.17 | 771 (T) | 609 | 301 | [25,26] | |
| Kaempferol 7-O-glucoside-3-rutinoside | 4.8 | 755 | 593 | [25,26] | ||
| Quercetin 3-O-rutinoside | 6.8 | 609 * | 301 | [10,25,26,27] | ||
| Quercetin-3-O-glucoside | 6.8 | 463 | 301 | [25,26] | ||
| Kaempferol 3-O-rutinoside | 7.7 | 593 * | 285 | [10,25,26,27] | ||
| 9-Flt-PhytoP | 5 | 327.2 (T) | 171.1 | [25] |
| Compounds | Rt | [M-H]− m/z | MSn | [AF-H]− | [AQ-H]− | [AC-H]− | [AF-H-18]− | [AQ-H-18]− | [p.CoA-H]− | Author |
|---|---|---|---|---|---|---|---|---|---|---|
| 3-O-Caffeoylquinic acid | 1.9 | 179 (T) | [26] | |||||||
| 3-p-Cumaroylquinic acid | 2.6 | 337 | 173 (T) | 166 * | [26] | |||||
| 3-O-Feluroylquinic acid | 3.2 | 367 | 193 * | 173 (T) | [26] | |||||
| 5-O-Caffeoylquinic acid | 3.3 | 353 | 191 * | 179 (T) | [25,26] | |||||
| Ferulic acid hexoside | 3.5 | 355 | 193 * | 175 | [26] | |||||
| Ferulic acid hexoside | 3.8 | 355 | 193 * | 175 | [26] | |||||
| Ferulic acid hexoside | 4.1 | 431 | 385 205 153 * | [25] |
| Treatment | Fasting Glucose mg/dL | Fasting Insulin (µUI/mL) | HOMA-IR |
|---|---|---|---|
| Normoglycemic | 113 ± 5 **** | 3.2 ± 0.3 **** | 1 ± 0.1 **** |
| Vehicle | 463 ± 18 | 27 ± 3.5 | 31 ± 4.1 |
| Metformin 250 mg/kg | 339 ± 29 **** | 18 ± 3.9 *** | 15 ± 2.5 **** |
| BFPP 50 mg/kg | 335 ± 37 **** | 11 ± 1.3 **** | 9 ± 0.4 **** |
| BFPP 100 mg/kg | 299 ± 18 **** | 11 ± 1.4 **** | 8 ± 1 **** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valderrama, I.H.; Echeverry, S.M.; Rey, D.P.; Rodríguez, I.A.; Silva, F.R.M.B.; Costa, G.M.; Ospina-Giraldo, L.F.; Aragón, D.M. Extract of Calyces from Physalis peruviana Reduces Insulin Resistance and Oxidative Stress in Streptozotocin-Induced Diabetic Mice. Pharmaceutics 2022, 14, 2758. https://doi.org/10.3390/pharmaceutics14122758
Valderrama IH, Echeverry SM, Rey DP, Rodríguez IA, Silva FRMB, Costa GM, Ospina-Giraldo LF, Aragón DM. Extract of Calyces from Physalis peruviana Reduces Insulin Resistance and Oxidative Stress in Streptozotocin-Induced Diabetic Mice. Pharmaceutics. 2022; 14(12):2758. https://doi.org/10.3390/pharmaceutics14122758
Chicago/Turabian StyleValderrama, Ivonne Helena, Sandra Milena Echeverry, Diana Patricia Rey, Ingrid Andrea Rodríguez, Fátima Regina Mena Barreto Silva, Geison M. Costa, Luis Fernando Ospina-Giraldo, and Diana Marcela Aragón. 2022. "Extract of Calyces from Physalis peruviana Reduces Insulin Resistance and Oxidative Stress in Streptozotocin-Induced Diabetic Mice" Pharmaceutics 14, no. 12: 2758. https://doi.org/10.3390/pharmaceutics14122758
APA StyleValderrama, I. H., Echeverry, S. M., Rey, D. P., Rodríguez, I. A., Silva, F. R. M. B., Costa, G. M., Ospina-Giraldo, L. F., & Aragón, D. M. (2022). Extract of Calyces from Physalis peruviana Reduces Insulin Resistance and Oxidative Stress in Streptozotocin-Induced Diabetic Mice. Pharmaceutics, 14(12), 2758. https://doi.org/10.3390/pharmaceutics14122758