
Enhancing In Vivo Electroporation Efficiency through Hyaluronidase: Insights into Plasmid Distribution and Optimization Strategies

 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. pDNA Constructs
2.2. Animals
2.3. Intramuscular Electroporation
2.4. Bioimpedance Measurements and Fitting
2.5. PNA-pDNA Labeling
2.6. Blood Collection, Tissue Processing, and Sectioning
2.7. Microscopy and Image Acquisition
2.8. ELISA Analysis
2.9. Statistical Analysis
3. Results
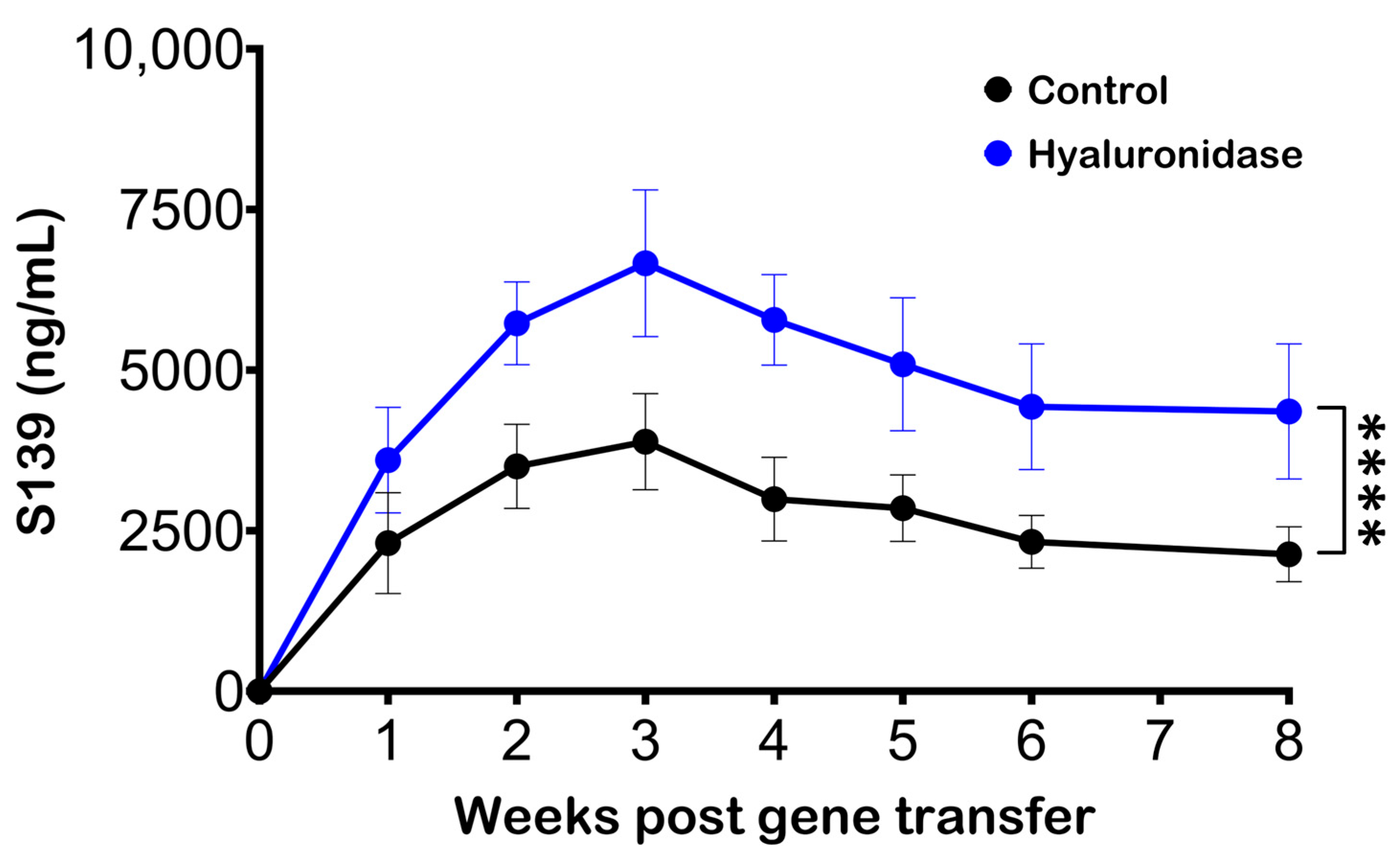
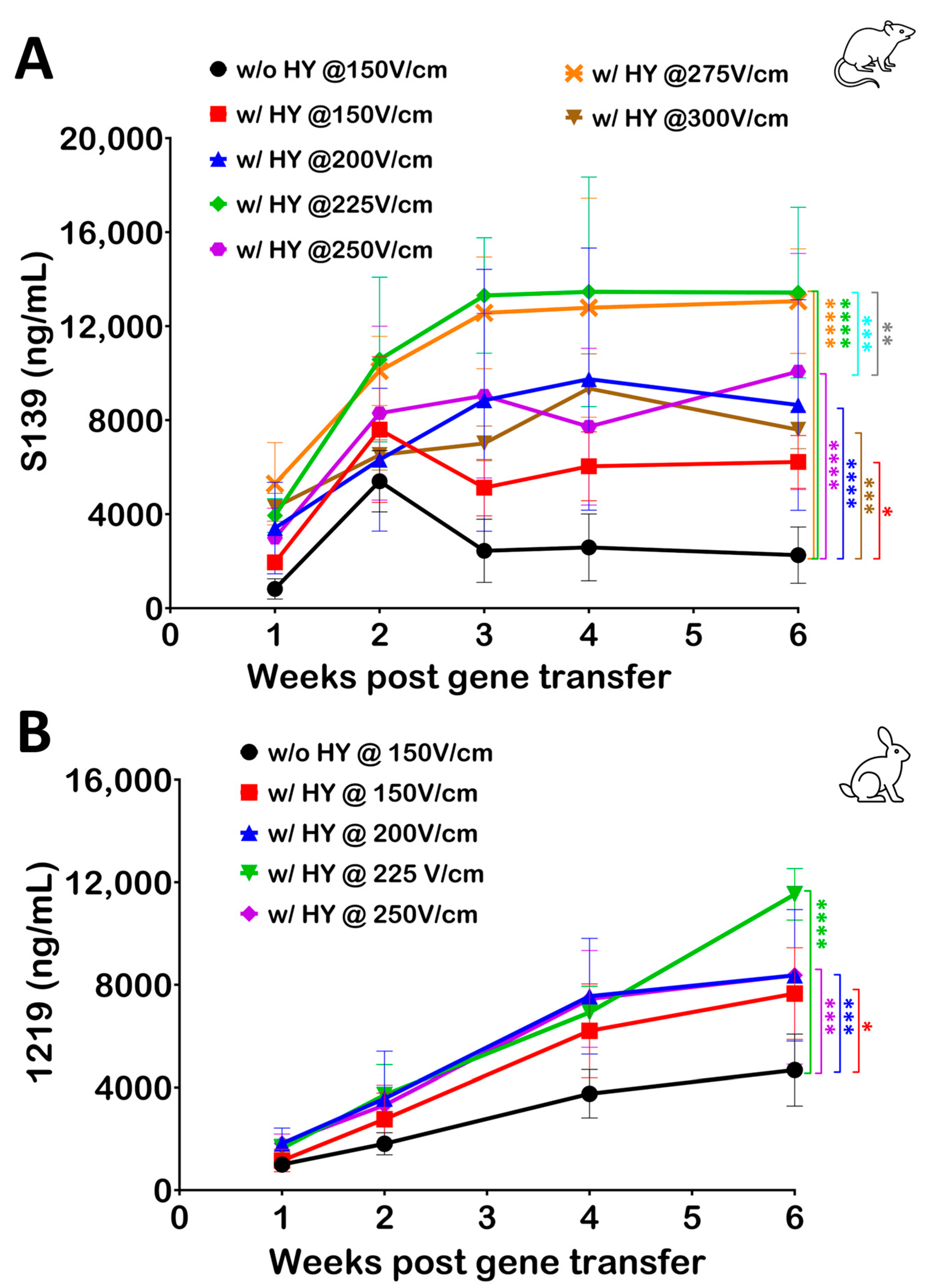
3.1. Hyaluronidase Treatment Improves Serum Antibody Levels following Electroporation of pDNA
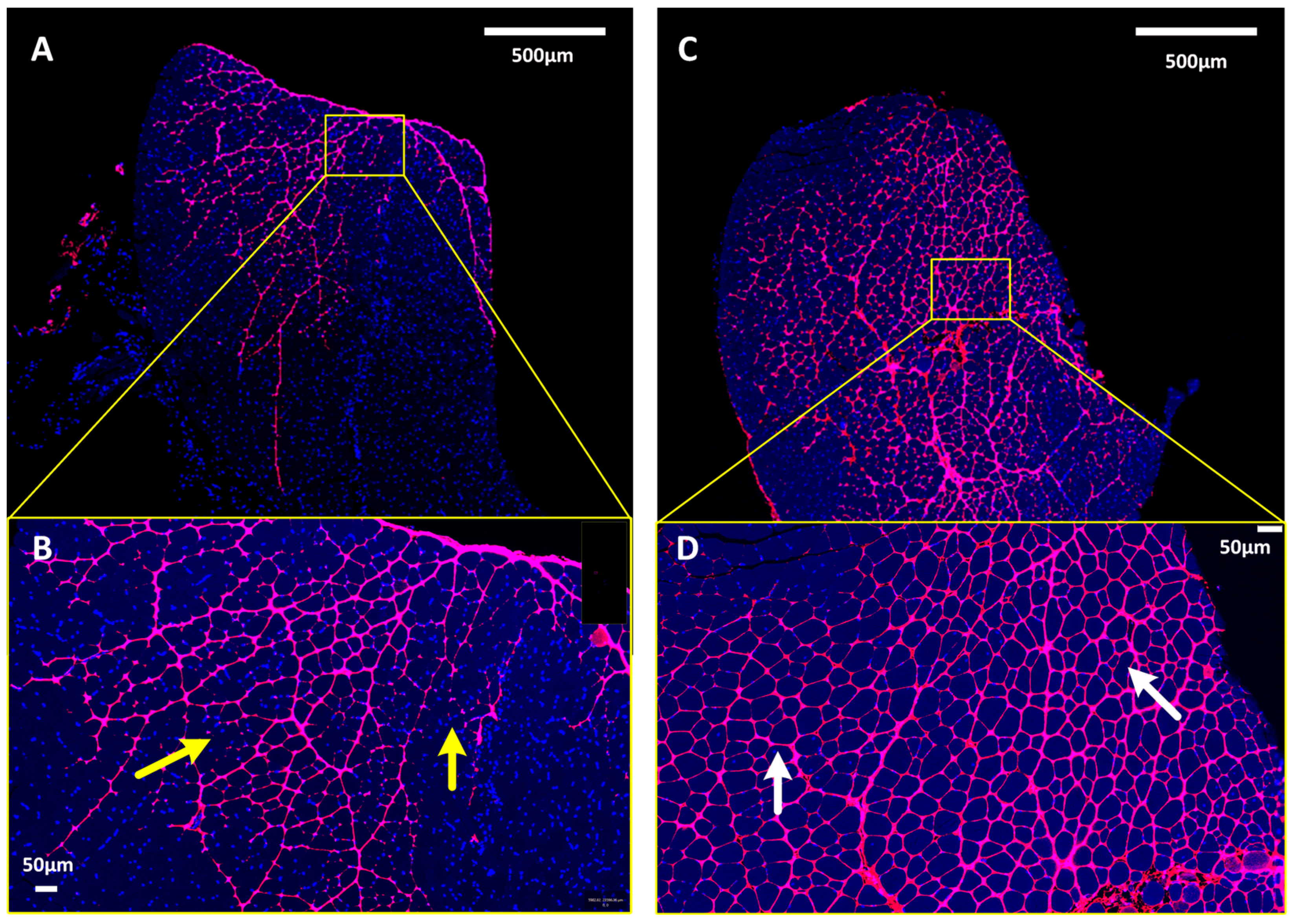
3.2. pDNA Distribution in Skeletal Muscle
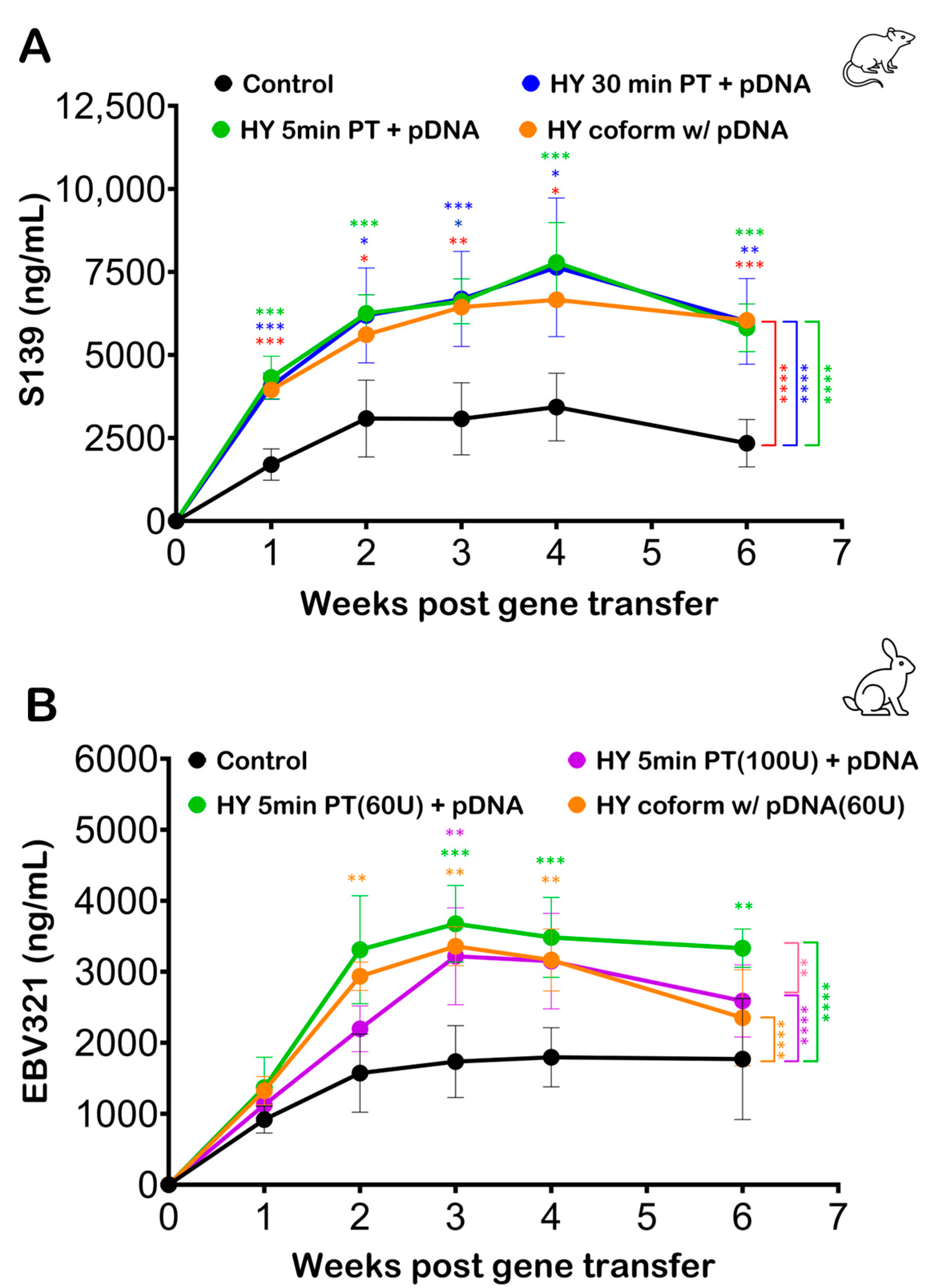
3.3. Effect of Hyaluronidase Pretreatment on Electroporation Efficiency
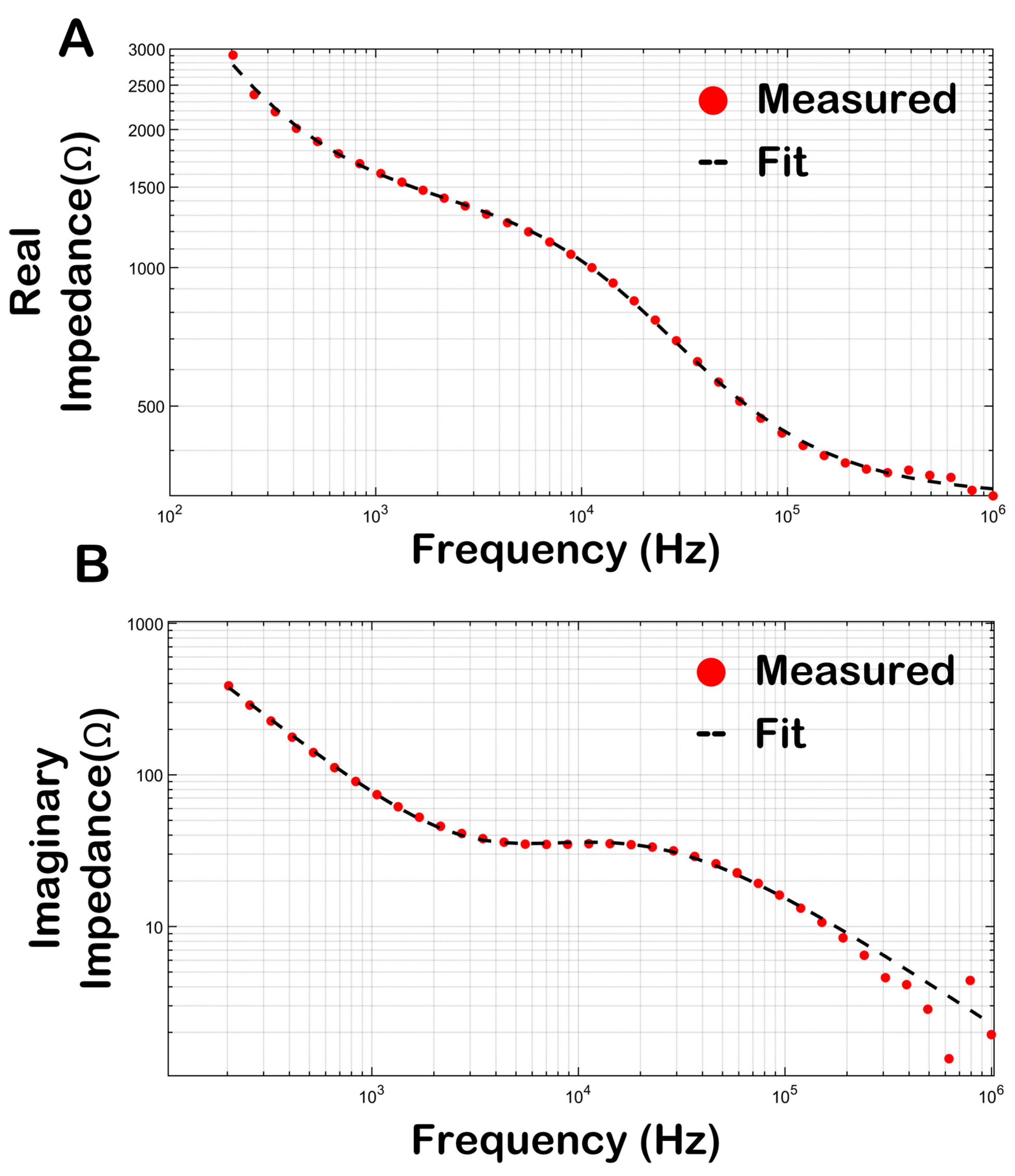
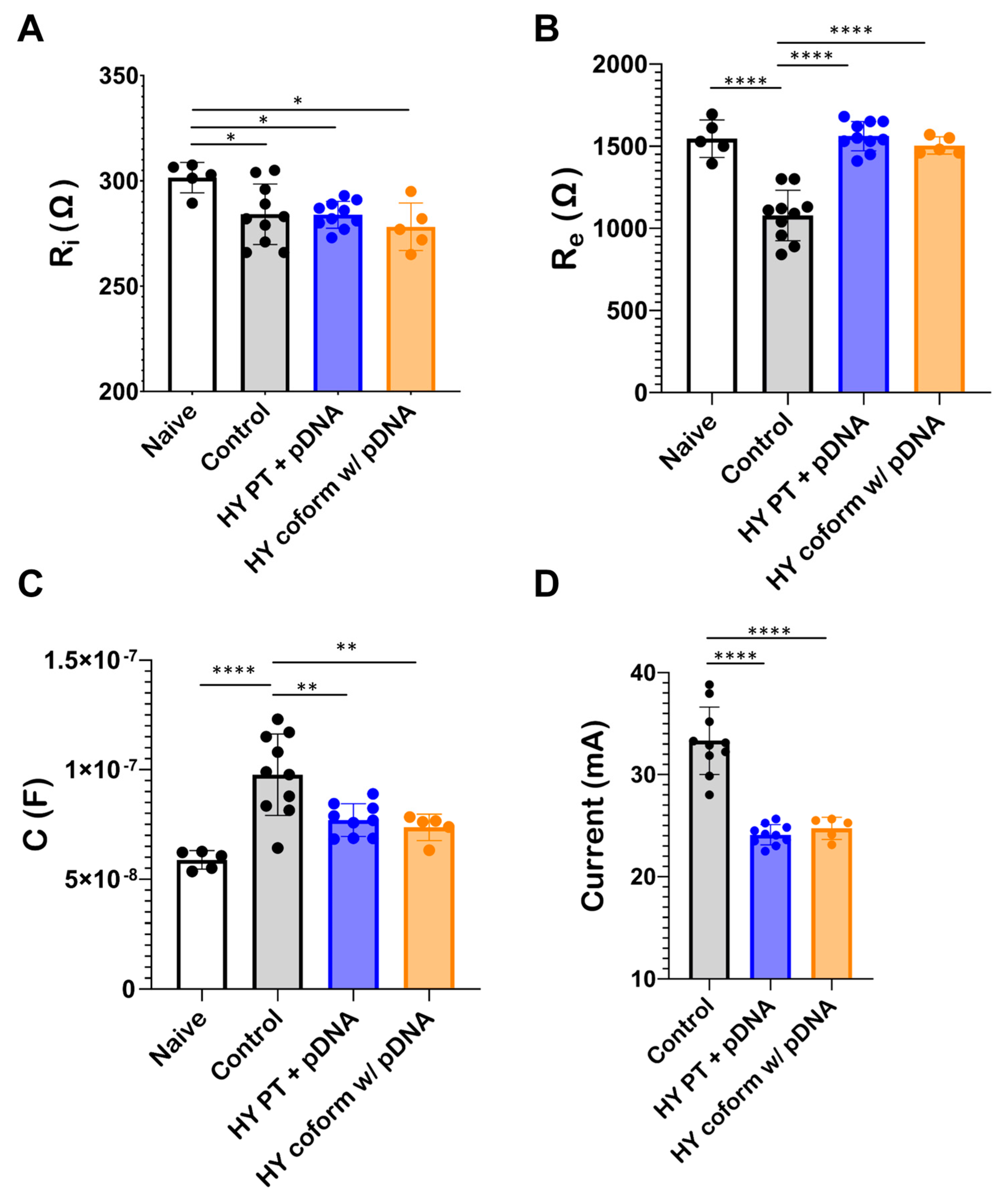
3.4. Bioimpedance of Skeletal Muscle
3.5. Optimizing Electroporation Parameters following Hyaluronidase Treatment
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wolff, J.A.; Malone, R.W.; Williams, P.; Chong, W.; Acsadi, G.; Jani, A.; Felgner, P.L. Direct Gene Transfer into Mouse Muscle in Vivo. Science 1990, 247, 1465–1468. [Google Scholar] [CrossRef] [PubMed]
- Sung, Y.; Kim, S. Recent advances in the development of gene delivery systems. Biomater. Res. 2019, 23, 8. [Google Scholar] [CrossRef] [PubMed]
- Al-Dosari, M.S.; Gao, X. Nonviral Gene Delivery: Principle, Limitations, and Recent Progress. AAPS J. 2009, 11, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Felgner, P.L.; Gadek, T.R.; Holm, M.; Roman, R.; Chan, H.W.; Wenz, M.; Northrop, J.P.; Ringold, G.M.; Danielsen, M. Lipofection: A highly efficient, lipid-mediated DNA-transfection procedure. Proc. Natl. Acad. Sci. USA 1987, 84, 7413–7417. [Google Scholar] [CrossRef] [PubMed]
- Boussif, O.; Lezoualc’h, F.; Zanta, M.A.; Mergny, M.D.; Scherman, D.; Demeneix, B.; Behr, J.P. A versatile vector for gene and oligonucleotide transfer into cells in culture and in vivo: Polyethylenimine. Proc. Natl. Acad. Sci. USA 1995, 92, 7297–7301. [Google Scholar] [CrossRef] [PubMed]
- Nathwani, A.C.; Tuddenham, E.G.D.; Rangarajan, S.; Rosales, C.; McIntosh, J.; Linch, D.C.; Chowdary, P.; Riddell, A.; Pie, A.J.; Harrington, C.; et al. Adenovirus-Associated Virus Vector–Mediated Gene Transfer in Hemophilia B. N. Engl. J. Med. 2011, 365, 2357–2365. [Google Scholar] [CrossRef] [PubMed]
- Dull, T.; Zufferey, R.; Kelly, M.; Mandel, R.J.; Nguyen, M.; Trono, D.; Naldini, L. A Third-Generation Lentivirus Vector with a Conditional Packaging System. J. Virol. 1998, 72, 8463–8471. [Google Scholar] [CrossRef] [PubMed]
- Neumann, E.; Schaefer-Ridder, M.; Wang, Y.; Hofschneider, P.H. Gene transfer into mouse lyoma cells by electroporation in high electric fields. EMBO J. 1982, 1, 841–845. [Google Scholar] [CrossRef]
- Aihara, H.; Miyazaki, J. Gene transfer into muscle by electroporation in vivo. Nat. Biotechnol. 1998, 16, 867–870. [Google Scholar] [CrossRef]
- Meaking, W.S.; Edgerton, J.; Wharton, C.W.; Meldrum, R.A. Electroporation-induced damage in mammalian cell DNA. Biochim. Biophys. Acta (BBA)-Gene Struct. Expr. 1995, 1264, 357–362. [Google Scholar] [CrossRef]
- Weaver, J.C. Electroporation: A general phenomenon for manipulating cells and tissues. J. Cell. Biochem. 1993, 51, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Napotnik, T.B.; Polajžer, T.; Miklavčič, D. Cell death due to electroporation—A review. Bioelectrochemistry 2021, 141, 107871. [Google Scholar]
- Gehl, J. Electroporation: Theory and methods, perspectives for drug delivery, gene therapy and research. Acta Physiol. Scand. 2003, 177, 437–447. [Google Scholar] [CrossRef]
- Kandušer, M.; Ušaj, M. Cell electrofusion: Past and future perspectives for antibody production and cancer cell vaccines. Expert Opin. Drug Deliv. 2014, 11, 1885–1898. [Google Scholar] [CrossRef] [PubMed]
- Weaver, J.C.; Chizmadzhev, Y.A. Theory of electroporation: A review. Bioelectrochem. Bioenerg. 1996, 41, 135–160. [Google Scholar] [CrossRef]
- De Robertis, M.; Pasquet, L.; Loiacono, L.; Bellard, E.; Messina, L.; Vaccaro, S.; Di Pasquale, R.; Fazio, V.M.; Rols, M.P.; Teissie, J.; et al. In Vivo Evaluation of a New Recombinant Hyaluronidase to Improve Gene Electro-Transfer Protocols for DNA-Based Drug Delivery against Cancer. Cancers 2018, 10, 405. [Google Scholar] [CrossRef]
- Andrews, C.D.; Luo, Y.; Sun, M.; Yu, J.; Goff, A.J.; Glass, P.J.; Padte, N.N.; Huang, Y.; Ho, D.D. In Vivo Production of Monoclonal Antibodies by Gene Transfer via Electroporation Protects against Lethal Influenza and Ebola Infections. Mol. Ther. Methods Clin. Dev. 2017, 7, 74–82. [Google Scholar] [CrossRef]
- Yu, Y.; Lee, P.; Ke, Y.; Zhang, Y.; Yu, Q.; Lee, J.; Li, M.; Song, J.; Chen, J.; Dai, J.; et al. A Humanized Anti-VEGF Rabbit Monoclonal Antibody Inhibits Angiogenesis and Blocks Tumor Growth in Xenograft Models. PLoS ONE 2010, 5, e9072. [Google Scholar] [CrossRef]
- Akerstrom, T.; Vedel, K.; Needham, J.; Hojman, P.; Kontou, E.; Hellsten, Y.; Wojtaszewski, J.F. Optimizing hyaluronidase dose and plasmid DNA delivery greatly improves gene electrotransfer efficiency in rat skeletal muscle. Biochem. Biophys. Rep. 2015, 4, 342–350. [Google Scholar] [CrossRef]
- Peri, D.; Deville, M.; Poignard, C.; Signori, E.; Natalini, R. Numerical optimization of plasmid DNA delivery combined with hyaluronidase injection for electroporation protocol. Comput. Methods Programs Biomed. 2020, 186, 105204. [Google Scholar] [CrossRef]
- Cole Kenneth, S. Permeability and Impermeability of Cell Membranes for Ions. Cold Spring Harb. Symp. Quant. Biol. 1940, 8, 110–122. [Google Scholar] [CrossRef]
- Freeborn Todd, J. A Survey of Fractional-Order Circuit Models for Biology and Biomedicine. IEEE J. Emerg. Sel. Top. Circuits Syst. 2013, 3, 416–424. [Google Scholar] [CrossRef]
- Egholm, M.; Christensen, L.; Deuholm, K.L.; Buchardt, O.; Coull, J.; Nielsen, P.E. Efficient pH-independent sequence-specific DNA binding by pseudoisocytosine-containing bis-PNA. Nucleic Acids Res. 1995, 23, 217–222. [Google Scholar] [CrossRef]
- Zelphati, O.; Liang, X.; Hobart, P.; Felgner, P.L. Gene Chemistry: Functionally and Conformationally Intact Fluorescent Plasmid DNA. Hum. Gene Ther. 1999, 10, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Zelphati, O.; Liang, X.; Nguyen, C.; Barlow, S.; Sheng, S.; Shao, Z.; Felgner, P. PNA-Dependent Gene Chemistry: Stable Coupling of Peptides and Oligonucleotides to Plasmid DNA. BioTechniques 2000, 28, 304–316. [Google Scholar] [CrossRef]
- Mennuni, C.; Calvaruso, F.; Zampaglione, I.; Rizzuto, G.; Rinaudo, D.; Dammassa, E.; Ciliberto, G.; Fattori, E.; La Monica, N. Hyaluronidase Increases Electrogene Transfer Efficiency in Skeletal Muscle. Hum. Gene Ther. 2002, 13, 355–365. [Google Scholar] [CrossRef] [PubMed]
- McMahon, J.M.; Signori, E.; Wells, K.E.; Fazio, V.M.; Wells, D.J. Optimisation of electrotransfer of plasmid into skeletal muscle by pretreatment with hyaluronidase–increased expression with reduced muscle damage. Gene Ther. 2001, 8, 1264–1270. [Google Scholar] [CrossRef] [PubMed]
- Buhren, B.A.; Schrumpf, H.; Hoff, N.-P.; Bölke, E.; Hilton, S.; Gerber, P.A. Hyaluronidase: From clinical applications to molecular and cellular mechanisms. Eur. J. Med. Res. 2016, 21, 5. [Google Scholar] [CrossRef]
- DeLorenzi, C. New High Dose Pulsed Hyaluronidase Protocol for Hyaluronic Acid Filler Vascular Adverse Events. Aesthet. Surg. J. 2017, 37, 814–825. [Google Scholar] [CrossRef]
- Naranjo-Hernández, D.; Reina-Tosina, J.; Min, M. Fundamentals, Recent Advances, and Future Challenges in Bioimpedance Devices for Healthcare Applications. J. Sens. 2019, 2019, e9210258. [Google Scholar] [CrossRef]
- Pagant, S.; Liberatore, R.A. In Vivo Electroporation of Plasmid DNA: A Promising Strategy for Rapid, Inexpensive, and Flexible Delivery of Anti-Viral Monoclonal Antibodies. Pharmaceutics 2021, 13, 1882. [Google Scholar] [CrossRef] [PubMed]
- Quigley, A.; Lowes, K.; Kornberg, A.J.; Cook, M.J.; Kapsa, R. Therapeutic DNA Delivery to Skeletal Muscle. Curr. Genom. 2006, 7, 179–190. [Google Scholar] [CrossRef]
- Bessis, N.; GarciaCozar, F.J.; Boissier, M.-C. Immune responses to gene therapy vectors: Influence on vector function and effector mechanisms. Gene Ther. 2004, 11 (Suppl. S1), S10–S17. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.E.; Ehrhardt, A.; Kay, M.A. Progress and problems with the use of viral vectors for gene therapy. Nat. Rev. Genet. 2003, 4, 346–358. [Google Scholar] [CrossRef] [PubMed]
- Mir, L.M.; Bureau, M.F.; Gehl, J.; Rangara, R.; Rouy, D.; Caillaud, J.M.; Delaere, P.; Branellec, D.; Schwartz, B.; Scherman, D. High-efficiency gene transfer into skeletal muscle mediated by electric pulses. Proc. Natl. Acad. Sci. USA 1999, 96, 4262–4267. [Google Scholar] [CrossRef] [PubMed]
- Mathiesen, I. Electropermeabilization of skeletal muscle enhances gene transfer in vivo. Gene Ther. 1999, 6, 508–514. [Google Scholar] [CrossRef]
- Bureau, M.F.; Naimi, S.; Ibad, R.T.; Seguin, J.; Georger, C.; Arnould, E.; Maton, L.; Blanche, F.; Delaere, P.; Scherman, D. Intramuscular plasmid DNA electrotransfer: Biodistribution and degradation. Biochim. Biophys. Acta (BBA)-Gene Struct. Expr. 2004, 1676, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Wohlrab, J.; Wohlrab, D.; Wohlrab, L.; Wohlrab, C.; Wohlrab, A. Use of Hyaluronidase for Pharmacokinetic Increase in Bioavailability of Intracutaneously Applied Substances. Ski. Pharmacol. Physiol. 2014, 27, 276–282. [Google Scholar] [CrossRef]
- Gollins, H.; McMahon, J.; Wells, K.E.; Wells, D.J. High-efficiency plasmid gene transfer into dystrophic muscle. Gene Ther. 2003, 10, 504–512. [Google Scholar] [CrossRef]
- Favre, D.; Cherel, Y.; Provost, N.; Blouin, V.; Ferry, N.; Moullier, P.; Salvetti, A. Hyaluronidase enhances recombinant adeno-associated virus (rAAV)-mediated gene transfer in the rat skeletal muscle. Gene Ther. 2000, 7, 1417–1420. [Google Scholar] [CrossRef]
- Chiarella, P.; Santis, S.D.; Fazio, V.M.; Signori, E. Hyaluronidase contributes to early inflammatory events induced by electrotransfer in mouse skeletal muscle. Hum. Gene Ther. 2013, 24, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Molnar, M.J.; Gilbert, R.; Lu, Y.; Liu, A.-B.; Guo, A.; Larochelle, N.; Orlopp, K.; Lochmuller, H.; Petrof, B.J.; Nalbantoglu, J.; et al. Factors Influencing the Efficacy, Longevity, and Safety of Electroporation-Assisted Plasmid-Based Gene Transfer into Mouse Muscles. Mol. Ther. 2004, 10, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Schertzer, J.D.; Plant, D.R.; Lynch, G.S. Optimizing Plasmid-Based Gene Transfer for Investigating Skeletal Muscle Structure and Function. Mol. Ther. 2006, 13, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Bruce, C.R.; Brolin, C.; Turner, N.; Cleasby, M.E.; van der Leij, F.R.; Cooney, G.J.; Kraegen, E.W. Overexpression of carnitine palmitoyltransferase I in skeletal muscle in vivo increases fatty acid oxidation and reduces triacylglycerol esterification. Am. J. Physiol.-Endocrinol. Metab. 2007, 292, E1231–E1237. [Google Scholar] [CrossRef] [PubMed]
- Bruce, C.R.; Hoy, A.J.; Turner, N.; Watt, M.J.; Allen, T.L.; Carpenter, K.; Cooney, G.J.; Febbraio, M.A.; Kraegen, E.W. Overexpression of carnitine palmitoyltransferase-1 in skeletal muscle is sufficient to enhance fatty acid oxidation and improve high-fat diet-induced insulin resistance. Diabetes 2009, 58, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Hollevoet, K.; Thomas, D.; Compernolle, G.; Vermeire, G.; De Smidt, E.; De Vleeschauwer, S.; Smith, T.R.F.; Fisher, P.D.; Dewilde, M.; Geukens, N.; et al. Clinically relevant dosing and pharmacokinetics of DNA-encoded antibody therapeutics in a sheep model. Front. Oncol. 2022, 12, 1017612. [Google Scholar] [CrossRef]
- Schommer, N.N.; Nguyen, J.; Yung, B.S.; Schultheis, K.; Muthumani, K.; Weiner, D.B.; Humeau, L.; Broderick, K.E.; Smith, T.R. Active Immunoprophylaxis and Vaccine Augmentations Mediated by a Novel Plasmid DNA Formulation. Hum. Gene Ther. 2019, 30, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Wise, M.C.; Xu, Z.; Tello-Ruiz, E.; Beck, C.; Trautz, A.; Patel, A.; Elliott, S.T.; Chokkalingam, N.; Kim, S.; Kerkau, M.G.; et al. In vivo delivery of synthetic DNA–encoded antibodies induces broad HIV-1–neutralizing activity. J. Clin. Investig. 2020, 130, 827–837. [Google Scholar] [CrossRef]
- Foster, K.R.; Schwan, H.P. Dielectric properties of tissues and biological materials: A critical review. Crit. Rev. Biomed. Eng. 1989, 17, 25–104. [Google Scholar]
- Gabriel, C.; Gabriel, S.; Corthout, E. The dielectric properties of biological tissues: I. Literature survey. Phys. Med. Biol. 1996, 41, 2231. [Google Scholar] [CrossRef]
- Suster, M.A.; Vitale, N.H.; Maji, D.; Mohseni, P. A Circuit Model of Human Whole Blood in a Microfluidic Dielectric Sensor. IEEE Trans. Circuits Syst. II Express Br. 2016, 63, 1156–1160. [Google Scholar]
- Maji, D.; De La Fuente, M.; Kucukal, E.; Sekhon, U.D.S.; Schmaier, A.H.; Gupta, A.S.; Gurkan, U.A.; Nieman, M.T.; Stavrou, E.X.; Mohseni, P.; et al. Assessment of whole blood coagulation with a microfluidic dielectric sensor. J. Thromb. Haemost. 2018, 16, 2050–2056. [Google Scholar] [CrossRef]
- Castizo-Olier, J.; Irurtia, A.; Jemni, M.; Carrasco-Marginet, M.; Fernández-García, R.; Rodríguez, F.A. Bioelectrical impedance vector analysis (BIVA) in sport and exercise: Systematic review and future perspectives. PLoS ONE 2018, 13, e0197957. [Google Scholar] [CrossRef] [PubMed]
- Kyle, U.G.; Bosaeus, I.; De Lorenzo, A.D.; Deurenberg, P.; Elia, M.; Gomez, J.M.; Heitmann, B.L.; Kent-Smith, L.; Melchior, J.-C.; Pirlich, M.; et al. Bioelectrical impedance analysis—Part I: Review of principles and methods. Clin. Nutr. 2004, 23, 1226–1243. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.; Weir, J.; Brown, L.; Weir, D. ASEP Procedures Recommendation I: Accurate Assessment of Muscular Strength and Power. J. Exerc. Physiol. Online 2001, 4, 1–21. [Google Scholar]
- Fu, B.; Freeborn, T.J. Cole-impedance parameters representing biceps tissue bioimpedance in healthy adults and their alterations following eccentric exercise. J. Adv. Res. 2020, 25, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Nakamura, T.; Kusuhara, T.; Kenichi, K.; Kuniyasu, K.; Kawashima, T.; Hanayama, K. Effectiveness of impedance parameters for muscle quality evaluation in healthy men. J. Physiol. Sci. JPS 2020, 70, 53. [Google Scholar] [CrossRef]
- Rutkove, S.B.; Callegari, S.; Concepcion, H.; Mourey, T.; Widrick, J.; Nagy, J.A.; Nath, A.K. Electrical impedance myography detects age-related skeletal muscle atrophy in adult zebrafish. Sci. Rep. 2023, 13, 7191. [Google Scholar] [CrossRef] [PubMed]
- Eikenes, L.; Tari, M.; Tufto, I.; Bruland, Ø.S.; Davies, C.D.L. Hyaluronidase induces a transcapillary pressure gradient and improves the distribution and uptake of liposomal doxorubicin (CaelyxTM) in human osteosarcoma xenografts. Br. J. Cancer 2005, 93, 81–88. [Google Scholar] [CrossRef]
- Hartikka, J.; Sukhu, L.; Buchner, C.; Hazard, D.; Bozoukova, V.; Margalith, M.; Nishioka, W.K.; Wheeler, C.J.; Manthorp, M.; Sawdey, M. Electroporation-Facilitated Delivery of Plasmid DNA in Skeletal Muscle: Plasmid Dependence of Muscle Damage and Effect of Poloxamer 188. Mol. Ther. 2001, 4, 407–415. [Google Scholar] [CrossRef]
- Li, F.; Yamaguchi, K.; Okada, K.; Matsushita, K.; Enatsu, N.; Chiba, K.; Yue, H.; Fujisawa, M. Efficient Transfection of DNA into Primarily Cultured Rat Sertoli Cells by Electroporation1. Biol. Reprod. 2013, 88, 61. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maji, D.; Miguela, V.; Cameron, A.D.; Campbell, D.A.; Sasset, L.; Yao, X.; Thompson, A.T.; Sussman, C.; Yang, D.; Miller, R.; et al. Enhancing In Vivo Electroporation Efficiency through Hyaluronidase: Insights into Plasmid Distribution and Optimization Strategies. Pharmaceutics 2024, 16, 547. https://doi.org/10.3390/pharmaceutics16040547
Maji D, Miguela V, Cameron AD, Campbell DA, Sasset L, Yao X, Thompson AT, Sussman C, Yang D, Miller R, et al. Enhancing In Vivo Electroporation Efficiency through Hyaluronidase: Insights into Plasmid Distribution and Optimization Strategies. Pharmaceutics. 2024; 16(4):547. https://doi.org/10.3390/pharmaceutics16040547
Chicago/Turabian StyleMaji, Debnath, Verónica Miguela, Andrew D. Cameron, Delcora A. Campbell, Linda Sasset, Xin Yao, Andy T. Thompson, Carleigh Sussman, David Yang, Robert Miller, and et al. 2024. "Enhancing In Vivo Electroporation Efficiency through Hyaluronidase: Insights into Plasmid Distribution and Optimization Strategies" Pharmaceutics 16, no. 4: 547. https://doi.org/10.3390/pharmaceutics16040547
APA StyleMaji, D., Miguela, V., Cameron, A. D., Campbell, D. A., Sasset, L., Yao, X., Thompson, A. T., Sussman, C., Yang, D., Miller, R., Drozdz, M. M., & Liberatore, R. A. (2024). Enhancing In Vivo Electroporation Efficiency through Hyaluronidase: Insights into Plasmid Distribution and Optimization Strategies. Pharmaceutics, 16(4), 547. https://doi.org/10.3390/pharmaceutics16040547