


Derivatization of Hyaluronan to Target Neuroblastoma and Neuroglioma Expressing CD44
 ,
,  ,
,  ,
,  ,
,  and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Synthesis of HA-R
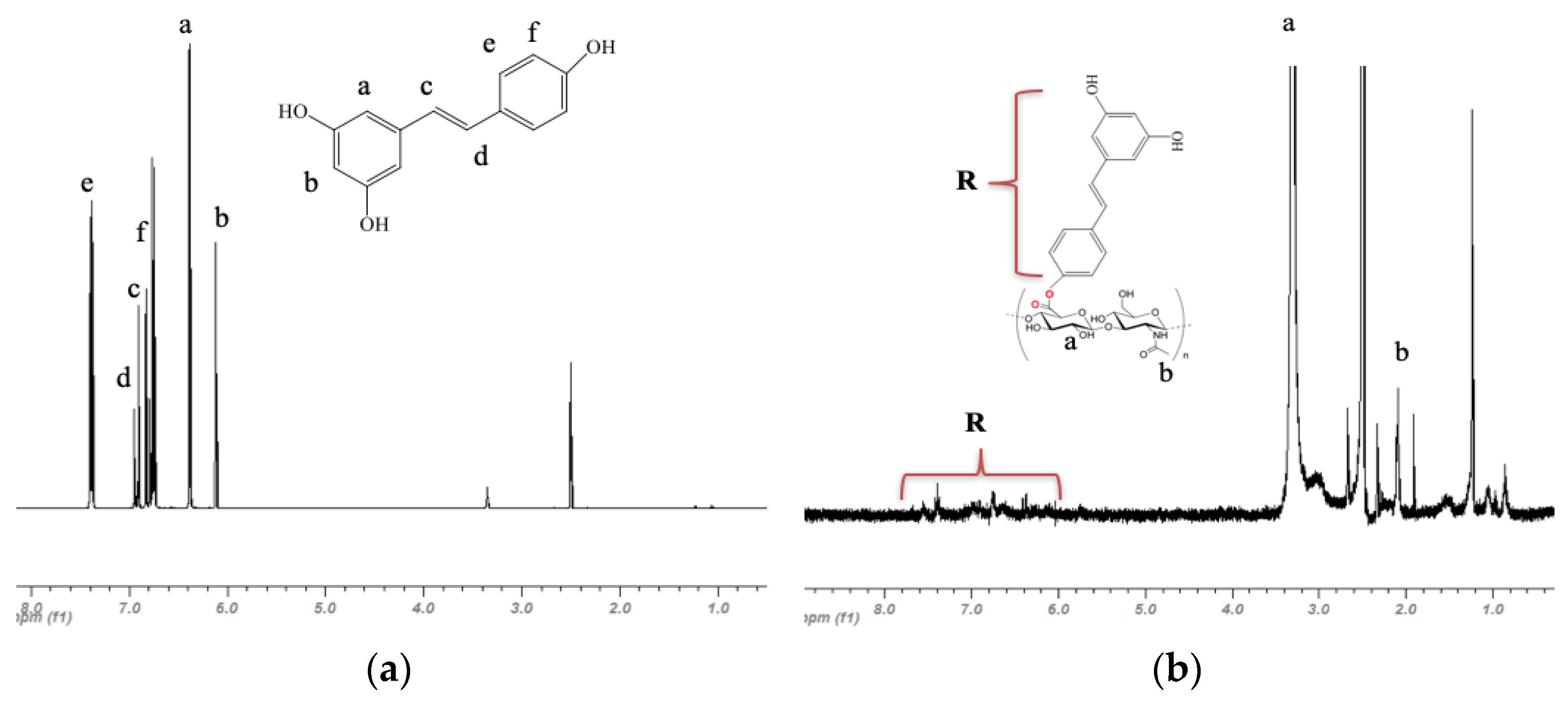
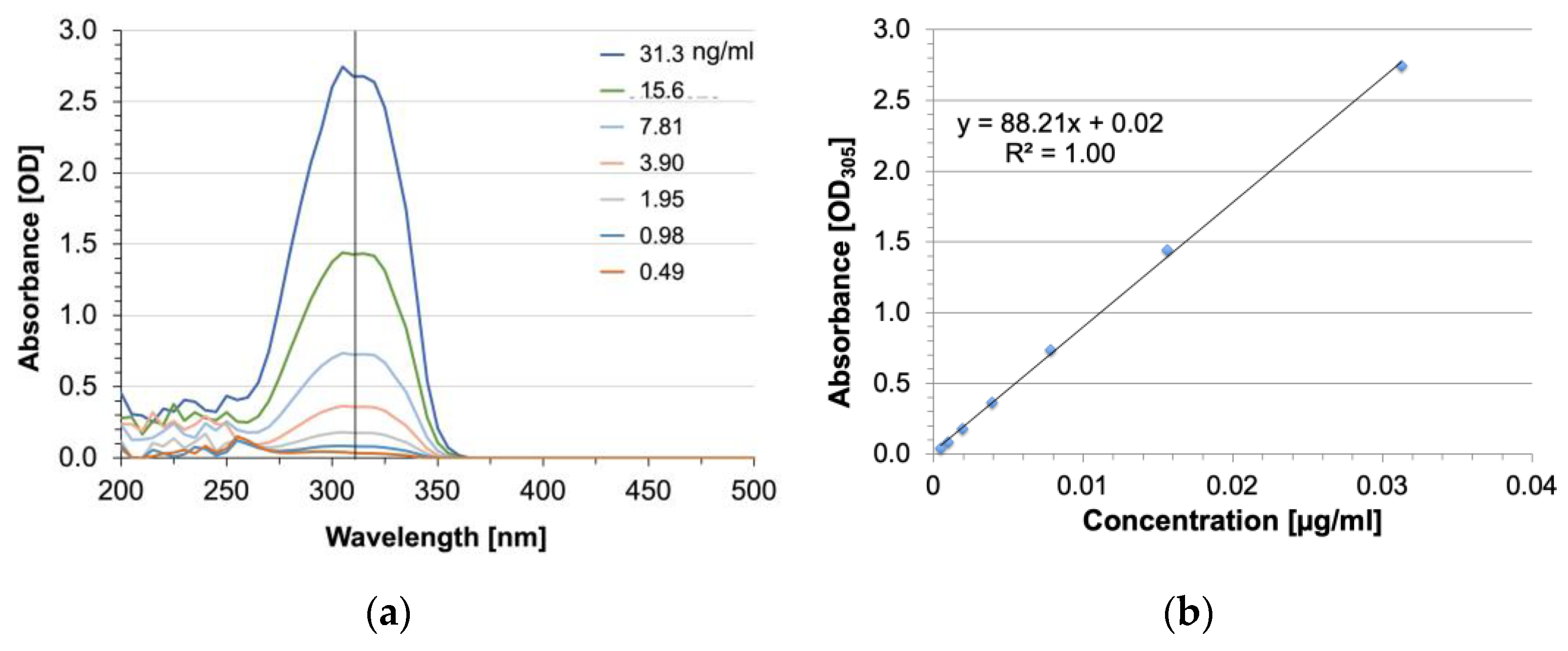
2.2. Characterization of HA-R
2.3. Cell Culture
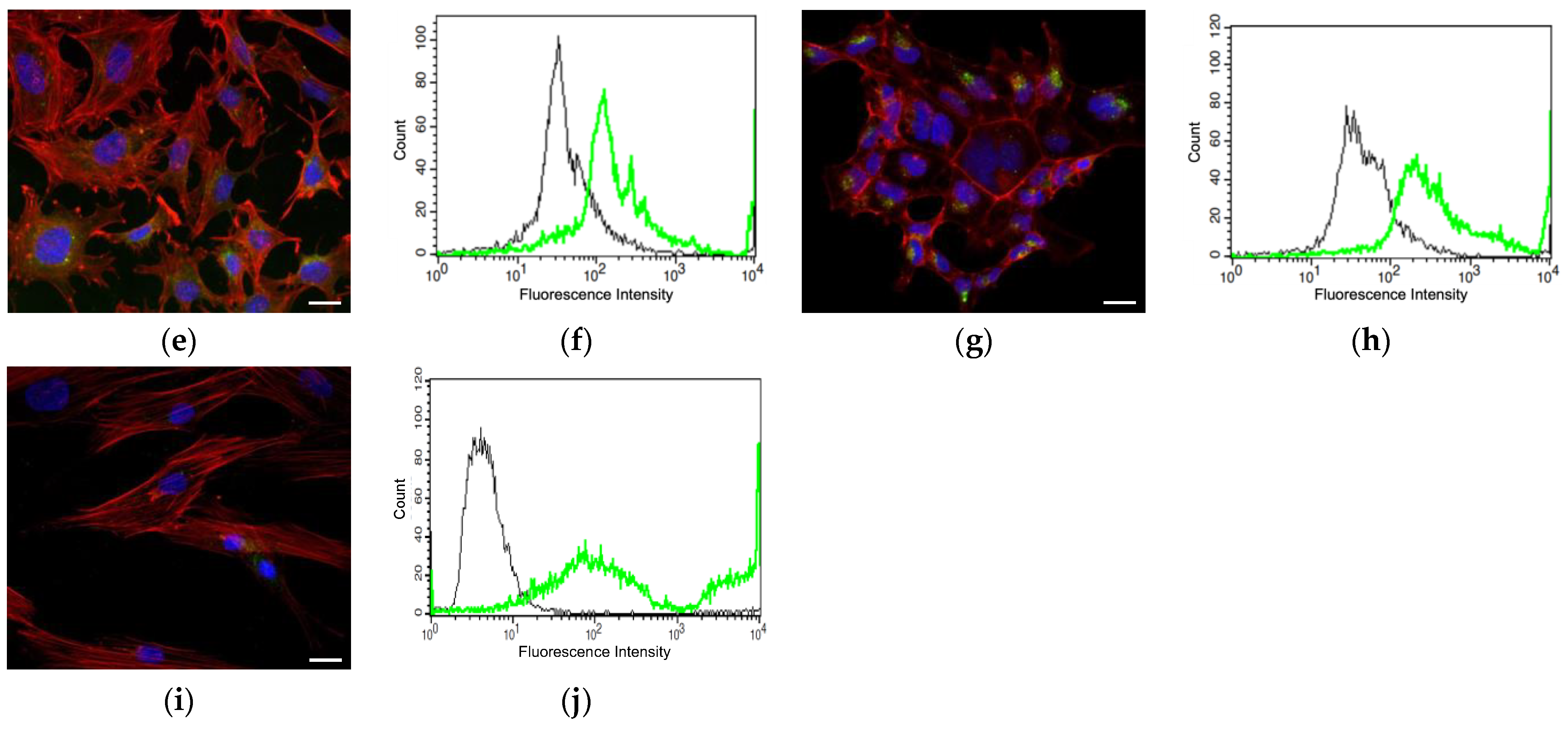
2.4. Cellular Uptake
2.5. CD44 Expression
2.6. Lysosomal Tracking
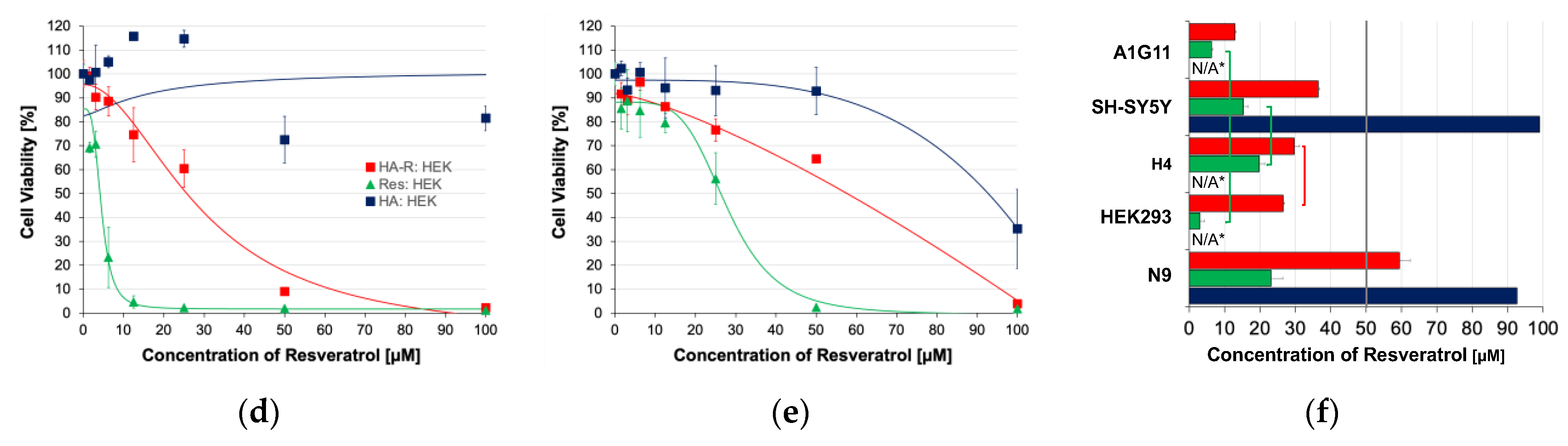
2.7. Cytotoxicity Study
2.8. Data Analysis
3. Results and Discussion
3.1. Characterization of HA-R
3.2. CD44 Expression
3.3. HA Uptake
3.4. In Vitro Toxicity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
References
- Margolis, R.U.; Margolis, R.K.; Chang, L.B.; Preti, C. Glycosaminoglycans of brain during development. Biochemistry 1975, 14, 85–88. [Google Scholar] [CrossRef]
- Morriss-Kay, G.M.; Tuckett, F.; Solursh, M. The effects of Streptomyces hyaluronidase on tissue organization and cell cycle time in rat embryos. J. Embryol. Exp. Morphol. 1986, 98, 59–70. [Google Scholar] [CrossRef]
- Bignami, A.; Asher, R. Some observations on the localization of hyaluronic acid in adult, newborn and embryonal rat brain. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 1992, 10, 45–57. [Google Scholar] [CrossRef]
- Preston, M.; Sherman, L.S. Neural stem cell niches: Roles for the hyaluronan-based extracellular matrix. Front. Biosci. (Sch. Ed.) 2011, 3, 1165–1179. [Google Scholar] [CrossRef]
- Sherman, L.S.; Struve, J.N.; Rangwala, R.; Wallingford, N.M.; Tuohy, T.M.; Kuntz, C., IV. Hyaluronate-based extracellular matrix: Keeping glia in their place. Glia 2002, 38, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Bignami, A.; Asher, R.; Perides, G.; Rahemtulla, F. The extracellular matrix of cerebral gray matter: Golgi’s pericellular net and Nissl's nervösen grau revisited. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 1992, 10, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, C.; Syková, E. Extracellular space structure revealed by diffusion analysis. Trends Neurosci. 1998, 21, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Fowke, T.M.; Karunasinghe, R.N.; Bai, J.Z.; Jordan, S.; Gunn, A.J.; Dean, J.M. Hyaluronan synthesis by developing cortical neurons in vitro. Sci. Rep. 2017, 7, 44135. [Google Scholar] [CrossRef]
- Asher, R.; Perides, G.; Vanderhaeghen, J.J.; Bignami, A. Extracellular matrix of central nervous system white matter: Demonstration of an hyaluronate-protein complex. J. Neurosci. Res. 1991, 28, 410–421. [Google Scholar] [CrossRef]
- Toole, B.P. Hyaluronan: From extracellular glue to pericellular cue. Nat. Rev. Cancer 2004, 4, 528–539. [Google Scholar] [CrossRef]
- Boregowda, R.K.; Appaiah, H.N.; Siddaiah, M.; Kumarswamy, S.B.; Sunila, S.; Thimmaiah, K.N.; Mortha, K.; Toole, B.; d Banerjee, S. Expression of hyaluronan in human tumor progression. J. Carcinog. 2006, 5, 2. [Google Scholar] [CrossRef] [PubMed]
- Pusch, A.; Boeckenhoff, A.; Glaser, T.; Kaminski, T.; Kirfel, G.; Hans, M.; Steinfarz, B.; Swandulla, D.; Kubitscheck, U.; Gieselmann, V.; et al. CD44 and hyaluronan promote invasive growth of B35 neuroblastoma cells into the brain. Biochim. Et Biophys. Acta 2010, 1803, 261–274. [Google Scholar] [CrossRef] [PubMed]
- Aruffo, A.; Stamenkovic, I.; Melnick, M.; Underhill, C.B.; Seed, B. CD44 is the principal cell surface receptor for hyaluronate. Cell 1990, 61, 1303–1313. [Google Scholar] [CrossRef] [PubMed]
- Misra, S.; Hascall, V.C.; Markwald, R.R.; Ghatak, S. Interactions between Hyaluronan and Its Receptors (CD44, RHAMM) Regulate the Activities of Inflammation and Cancer. Front. Immunol. 2015, 6, 201. [Google Scholar] [CrossRef] [PubMed]
- Hassn Mesrati, M.; Syafruddin, S.E.; Mohtar, M.A.; Syahir, A. CD44: A Multifunctional Mediator of Cancer Progression. Biomolecules 2021, 11, 1850. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.T.; Berry, P.A.; Hyde, C.; Stower, M.J.; Maitland, N.J. Prospective identification of tumorigenic prostate cancer stem cells. Cancer Res. 2005, 65, 10946–10951. [Google Scholar] [CrossRef] [PubMed]
- Lau, W.M.; Teng, E.; Chong, H.S.; Lopez, K.A.P.; Tay, A.Y.L.; Salto-Tellez, M.; Shabbir, A.; So, J.B.Y.; Chan, S.L. CD44v8-10 is a cancer-specific marker for gastric cancer stem cells. Cancer Res. 2014, 74, 2630–2641. [Google Scholar] [CrossRef] [PubMed]
- Screaton, G.R.; Bell, M.V.; Jackson, D.G.; Cornelis, F.B.; Gerth, U.; Bell, J.I. Genomic structure of DNA encoding the lymphocyte homing receptor CD44 reveals at least 12 alternatively spliced exons. Proc. Natl. Acad. Sci. USA 1992, 89, 12160–12164. [Google Scholar] [CrossRef] [PubMed]
- Prochazka, L.; Tesarik, R.; Turanek, J. Regulation of alternative splicing of CD44 in cancer. Cell. Signal. 2014, 26, 2234–2239. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Cai, L.; Udeani, G.O.; Slowing, K.V.; Thomas, C.F.; Beecher, C.W.; Fong, H.H.; Farnsworth, N.R.; Kinghorn, A.D.; Mehta, R.G.; et al. Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science 1997, 275, 218–220. [Google Scholar] [CrossRef]
- Lin, B. A Novel Resveratrol Analog: Its Cell Cycle Inhibitory, Pro-Apooptotic and Anti-Inflammatroy Activities on Human Tumor Cells; Kent State University: Kent, UK, 2006. [Google Scholar]
- Shah, K.N.; Crowder, D.C.; Overmeyer, J.H.; Maltese, W.A.; Yun, Y.H. Hyaluronan drug delivery dystems are promising for cancer therapy because of their selective attachment, enhanced uptake, and superior efficacy. Biomed. Eng. Lett. 2015, 5, 109–123. [Google Scholar] [CrossRef]
- Shah, K.N.; Ditto, A.J.; Crowder, D.C.; Overmeyer, J.H.; Tavana, H.; Maltese, W.A.; Yun, Y.H. Receptor-Mediated Attachment and Uptake of Hyaluronan Conjugates by Breast Cancer Cells. Mol. Pharm. 2017, 14, 3968–3977. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.F.; Lin, Y.K.; Huang, Z.R.; Fang, J.Y. Delivery of resveratrol, a red wine polyphenol, from solutions and hydrogels via the skin. Biol. Pharm. Bull. 2008, 31, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Ng, K.Y. Resveratrol-loaded calcium-pectinate beads: Effects of formulation parameters on drug release and bead characteristics. J. Pharm. Sci. 2010, 99, 840–860. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Chaudhury, A.; Ng, K.Y. Preparation and evaluation of zinc-pectin-chitosan composite particles for drug delivery to the colon: Role of chitosan in modifying in vitro and in vivo drug release. Int. J. Pharm. 2011, 406, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Teskac, K.; Kristl, J. The evidence for solid lipid nanoparticles mediated cell uptake of resveratrol. Int. J. Pharm. 2010, 390, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Aillon, K.L.; Cai, S.; Christian, J.M.; Davies, N.M.; Berkland, C.J.; Forrest, M.L. Pulmonary delivery of cisplatin-hyaluronan conjugates via endotracheal instillation for the treatment of lung cancer. Int. J. Pharm. 2010, 392, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Bernshaw, N.J.; Lu, Z.R.; Kopecek, J.; Prestwich, G.D. Targeted delivery of doxorubicin by HPMA copolymer-hyaluronan bioconjugates. Pharm. Res. 2002, 19, 396–402. [Google Scholar] [CrossRef]
- Dwibedi, V.; Rath, S.K.; Prakash, R.; Saxena, S. Response surface statistical optimization of fermentation parameters for resveratrol production by the endophytic fungus Arcopilus aureus and its tyrosinase inhibitory activity. Biotechnol. Lett. 2021, 43, 627–644. [Google Scholar] [CrossRef]
- Vázquez, B.E.R.; Rodríguez-Beas, C.; Iñiguez-Palomares, R.A.; Santacruz-Ortega, H.; Mendoza-Cruz, R.; Bazán-Díaz, L.S.; Soberanes, Y.; Rodríguez-León, E.; Navarro, R.E. Spectroscopic analysis and nuclear magnetic resonance for silver nanoparticles synthesized with trans-resveratrol and cis-resveratrol. Colloid Polym. Sci. 2022, 300, 465–475. [Google Scholar] [CrossRef]
- Bonechi, C.; Martini, S.; Ciani, L.; Lamponi, S.; Rebmann, H.; Rossi, C.; Ristori, S. Using Liposomes as Carriers for Polyphenolic Compounds: The Case of Trans-Resveratrol. PLoS ONE 2012, 7, e41438. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.J.; Yoon, H.Y.; Koo, H.; Ko, S.H.; Shim, J.S.; Lee, J.H.; Kim, K.; Kwon, I.C.; Kim, D.D. Self-assembled nanoparticles based on hyaluronic acid-ceramide (HA-CE) and Pluronic® for tumor-targeted delivery of docetaxel. Biomaterials 2011, 32, 7181–7190. [Google Scholar] [CrossRef]
- Park, J.S.; Yi, S.W.; Kim, H.J.; Park, K.H. Receptor-mediated gene delivery into human mesenchymal stem cells using hyaluronic acid-shielded polyethylenimine/pDNA nanogels. Carbohydr. Polym. 2016, 136, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Owen, S.C. Hyaluronic acid binding to CD44S is indiscriminate of molecular weight. Biochim. Et Biophys. Acta Biomembr. 2020, 1862, 183348. [Google Scholar] [CrossRef] [PubMed]
- Gross, N.; Balmas, K.; Brognara, C.B. Absence of functional CD44 hyaluronan receptor on human NMYC-amplified neuroblastoma cells. Cancer Res. 1997, 57, 1387–1393. [Google Scholar] [PubMed]
- Ferlemann, F.C.; Menon, V.; Condurat, A.L.; Rößler, J.; Pruszak, J. Surface marker profiling of SH-SY5Y cells enables small molecule screens identifying BMP4 as a modulator of neuroblastoma differentiation. Sci. Rep. 2017, 7, 13612. [Google Scholar] [CrossRef] [PubMed]
- De Marzo, A.M.; Bradshaw, C.; Sauvageot, J.; Epstein, J.I.; Miller, G.J. CD44 and CD44v6 downregulation in clinical prostatic carcinoma: Relation to Gleason grade and cytoarchitecture. Prostate 1998, 34, 162–168. [Google Scholar] [CrossRef]
- Shtivelman, E.; Bishop, J.M. Expression of CD44 is repressed in neuroblastoma cells. Mol. Cell. Biol. 1991, 11, 5446–5453. [Google Scholar] [PubMed]
- Tomé-Carneiro, J.; Larrosa, M.; González-Sarrías, A.; Tomás-Barberán, F.A.; García-Conesa, M.T.; Espín, J.C. Resveratrol and clinical trials: The crossroad from in vitro studies to human evidence. Curr. Pharm. Des. 2013, 19, 6064–6093. [Google Scholar] [CrossRef]
- McKenzie, J.L.; Dalchau, R.; Fabre, J.W. Biochemical characterisation and localization in brain of a human brain-leucocyte membrane glycoprotein recognised by a monoclonal antibody. J. Neurochem. 1982, 39, 1461–1466. [Google Scholar] [CrossRef]
- Bignami, A.; Dahl, D. Brain-specific hyaluronate-binding protein. A product of white matter astrocytes? J. Neurocytol. 1986, 15, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Cruz, T.F.; Quackenbush, E.J.; Letarte, M.; Moscarello, M.A. Elevated levels of a glycoprotein antigen (P-80) in gray and white matter of brain from victims of multiple sclerosis. Neurochem. Res. 1986, 11, 877–889. [Google Scholar] [CrossRef] [PubMed]
- Vogel, H.; Butcher, E.C.; Picker, L.J. H-CAM expression in the human nervous system: Evidence for a role in diverse glial interactions. J. Neurocytol. 1992, 21, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, H.; Tooyama, I.; Kawamata, T.; Ikeda, K.; McGeer, P.L. Morphological diversities of CD44 positive astrocytes in the cerebral cortex of normal subjects and patients with Alzheimer's disease. Brain Res. 1993, 632, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.L.; Liu, Z.; Shen, J.; Werner, A.; Kreutzberg, G.W.; Raivich, G. Regulation of the cell adhesion molecule CD44 after nerve transection and direct trauma to the mouse brain. J. Comp. Neurol. 2000, 426, 468–492. [Google Scholar] [CrossRef] [PubMed]
- Naruse, M.; Shibasaki, K.; Yokoyama, S.; Kurachi, M.; Ishizaki, Y. Dynamic changes of CD44 expression from progenitors to subpopulations of astrocytes and neurons in developing cerebellum. PLoS ONE 2013, 8, e53109. [Google Scholar] [CrossRef]
- Dahl, D.; Crosby, C.J.; Bignami, A. Neurofilament proteins in fish: A study with monoclonal antibodies reacting with mammalian NF 150K and NF 200K. J. Comp. Neurol. 1986, 250, 399–402. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vo, G.V.; Rao, K.M.; Chung, I.; Ha, C.-S.; An, S.S.A.; Yun, Y.H. Derivatization of Hyaluronan to Target Neuroblastoma and Neuroglioma Expressing CD44. Pharmaceutics 2024, 16, 836. https://doi.org/10.3390/pharmaceutics16060836
Vo GV, Rao KM, Chung I, Ha C-S, An SSA, Yun YH. Derivatization of Hyaluronan to Target Neuroblastoma and Neuroglioma Expressing CD44. Pharmaceutics. 2024; 16(6):836. https://doi.org/10.3390/pharmaceutics16060836
Chicago/Turabian StyleVo, Giau Van, Kummara Madhusudana Rao, Ildoo Chung, Chang-Sik Ha, Seong Soo A. An, and Yang H. Yun. 2024. "Derivatization of Hyaluronan to Target Neuroblastoma and Neuroglioma Expressing CD44" Pharmaceutics 16, no. 6: 836. https://doi.org/10.3390/pharmaceutics16060836
APA StyleVo, G. V., Rao, K. M., Chung, I., Ha, C.-S., An, S. S. A., & Yun, Y. H. (2024). Derivatization of Hyaluronan to Target Neuroblastoma and Neuroglioma Expressing CD44. Pharmaceutics, 16(6), 836. https://doi.org/10.3390/pharmaceutics16060836