
Assessment of the Efficacy of the Antihistamine Drug Rupatadine Used Alone or in Combination against Mycobacteria
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria and Culture Conditions
2.2. Antimicrobials
2.3. Evaluating Activity of RTD against Actively Growing Mycobacteria
2.4. Evaluating the Activity of RTD against Nonreplicating M. tuberculosis in Diverse Media
2.5. Evaluating the Activities of RTD in Combination with Anti-TB Drugs
2.6. Evaluating the In Vivo Anti-M. tuberculosis Activity of RTD
2.7. The Transcriptomic Profiling of RTD-Treated and Untreated M. tuberculosis
2.8. Statistical Analysis
3. Results
3.1. Antimycobacterial Activity of RTD
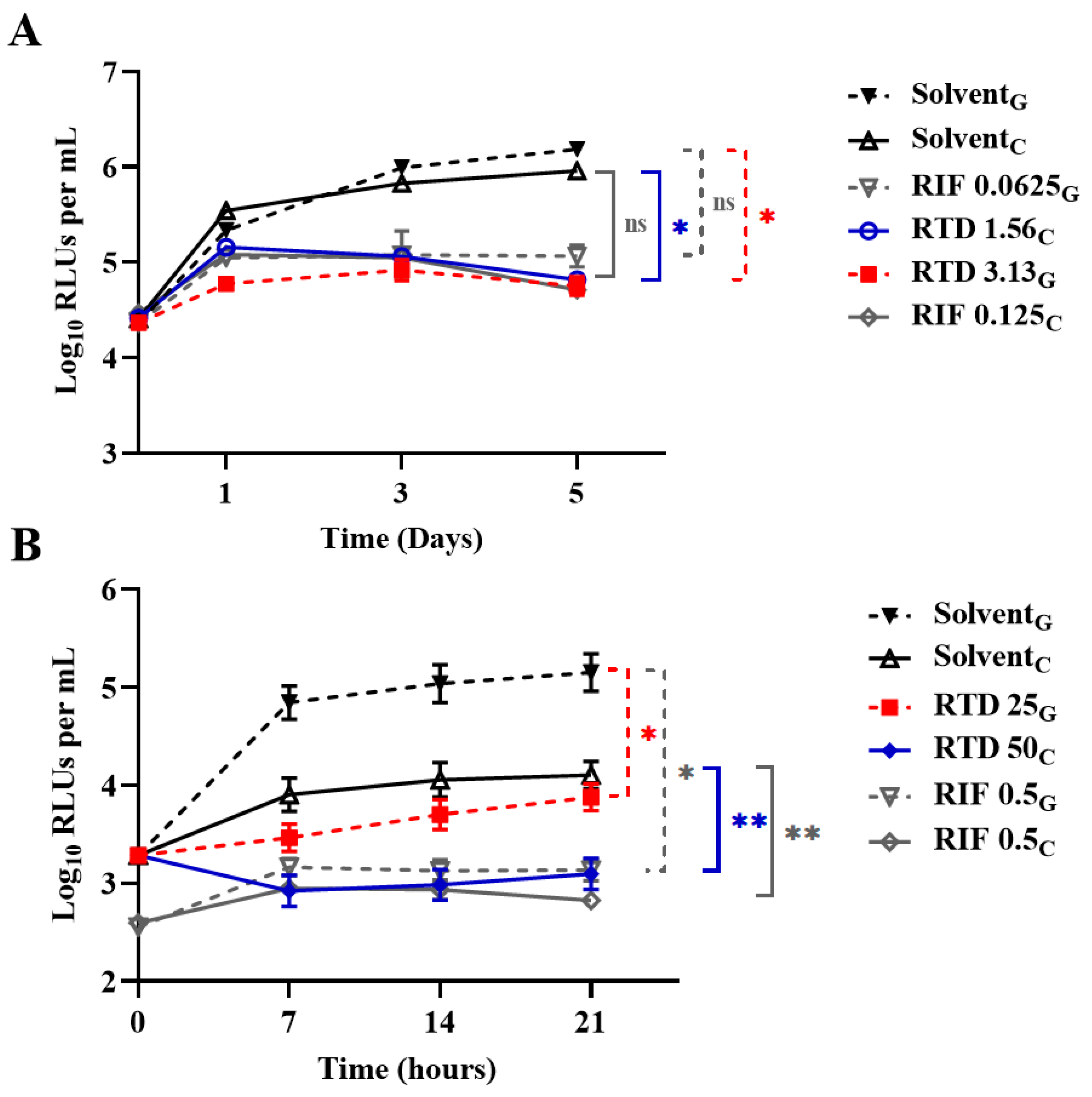
3.2. Activity of RTD against Nonreplicating AlRa Under Diverse Conditions
3.3. Partial Synergistic Effect of RTD in Combination with Several Antimycobacterial Drugs against M. tuberculosis
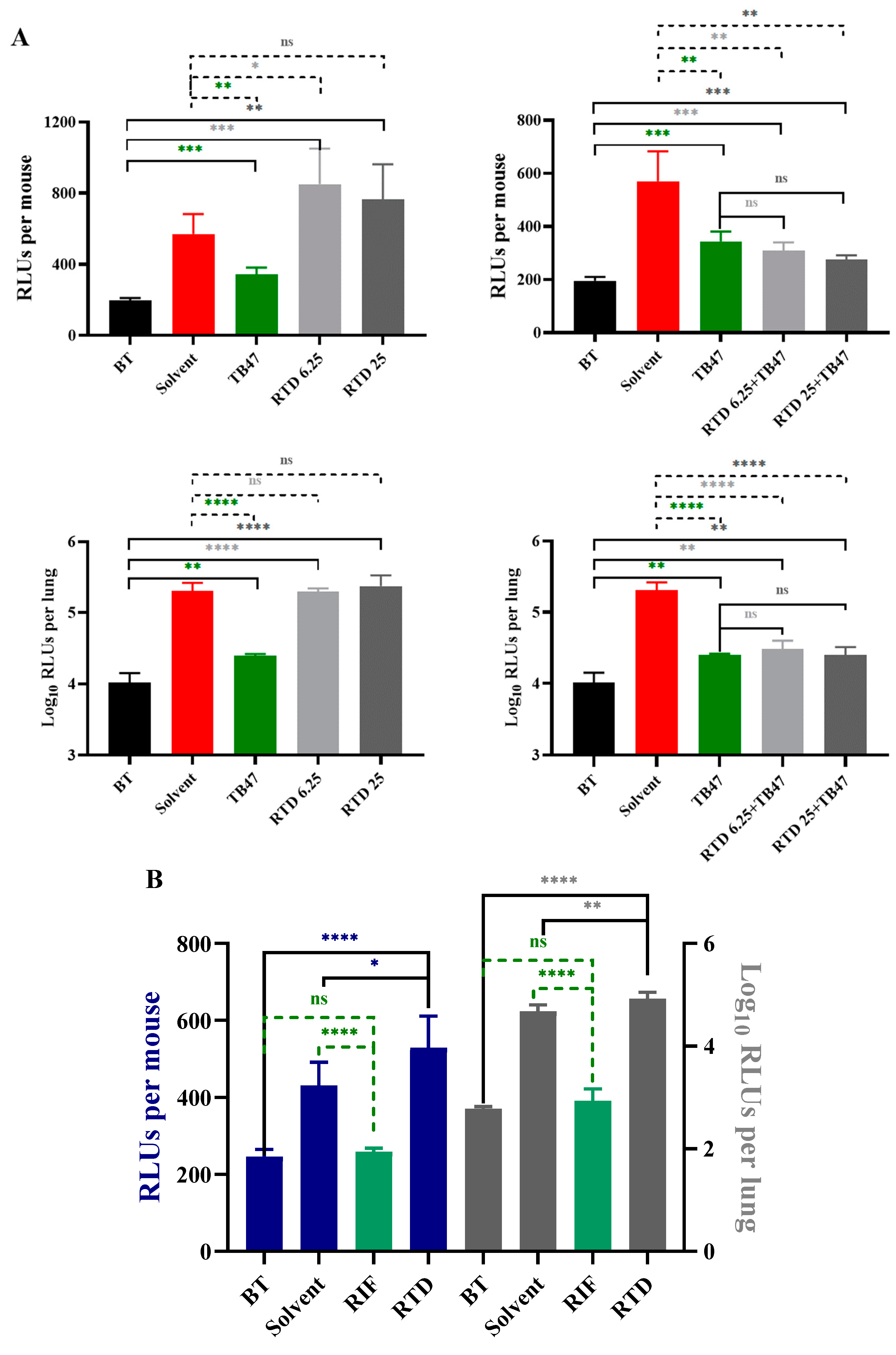
3.4. The In Vivo Anti-M. tuberculosis Activity of RTD
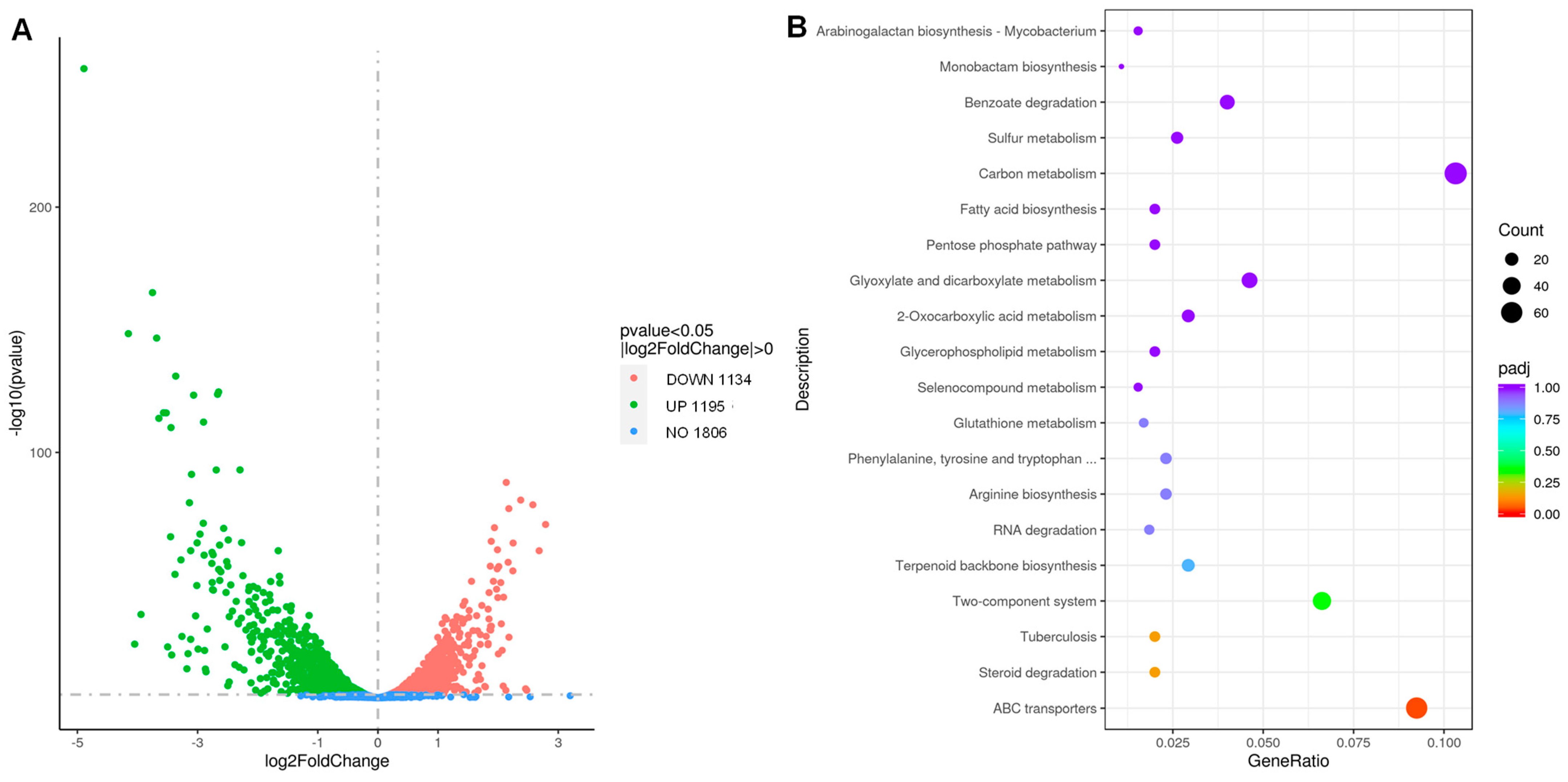
3.5. Transcriptome Profile of RTD-Treated and Untreated M. tuberculosis
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sia, J.K.; Rengarajan, J. Immunology of Mycobacterium tuberculosis infections. Microbiol. Spectr. 2019, 7, 1–37. [Google Scholar] [CrossRef]
- Urban, M.; Šlachtová, V.; Brulíková, L. Small organic molecules targeting the energy metabolism of Mycobacterium tuberculosis. Eur. J. Med. Chem. 2021, 212, 113139. [Google Scholar] [CrossRef] [PubMed]
- Getahun, H.; Matteelli, A.; Abubakar, I.; Aziz, M.A.; Baddeley, A.; Barreira, D.; den Boon, S.; Borroto Gutierrez, S.M.; Bruchfeld, J.; Burhan, E.; et al. Management of latent Mycobacterium tuberculosis infection: WHO guidelines for low tuberculosis burden countries. Eur. Respir. J. 2015, 46, 1563–1576. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Surolia, A. Mycobacterium tuberculosis: Surviving and indulging in an unwelcoming host. IUBMB Life 2018, 70, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.K.; Upadhyay, V. Epidemiology, diagnosis & treatment of non-tuberculous mycobacterial diseases. Indian J. Med. Res. 2020, 152, 185–226. [Google Scholar] [CrossRef] [PubMed]
- Shaku, M.T.; Ocius, K.L.; Apostolos, A.J.; Pires, M.M.; VanNieuwenhze, M.S.; Dhar, N.; Kana, B.D. Amidation of glutamate residues in mycobacterial peptidoglycan is essential for cell wall cross-linking. Front. Cell. Infect. Microbiol. 2023, 13, 1205829. [Google Scholar] [CrossRef]
- Falkinham, J.O. Challenges of NTM drug development. Front. Microbiol. 2018, 9, 1613. [Google Scholar] [CrossRef]
- Story-Roller, E.; Maggioncalda, E.C.; Cohen, K.A.; Lamichhane, G. Mycobacterium abscessus and β-Lactams: Emerging insights and potential opportunities. Front. Microbiol. 2018, 9, 2273. [Google Scholar] [CrossRef]
- Das, S.; Garg, T.; Chopra, S.; Dasgupta, A. Repurposing disulfiram to target infections caused by non-tuberculous mycobacteria. J. Antimicrob. Chemother. 2019, 74, 1317–1322. [Google Scholar] [CrossRef]
- Won, T.-B.; Kim, H.G.; Kim, J.-W.; Kim, J.K.; Kim, Y.H.; Kim, S.W.; Kim, H.Y.; Kim, D.W.; Kim, S.W.; Kim, C.-H.; et al. Efficacy and safety of rupatadine fumarate in the treatment of perennial allergic rhinitis: A multicenter, double-blinded, randomized, placebo-controlled, bridging study in Koreans. Asian Pac. J. Allergy Immunol. 2021. [Google Scholar] [CrossRef]
- Bachert, C.; van Cauwenberge, P.; Khaltaev, N. Allergic rhinitis and its impact on asthma. In collaboration with the World Health Organization. Executive summary of the workshop report. 7–10 December 1999, Geneva, Switzerland. Allergy 2002, 57, 841–855. [Google Scholar] [CrossRef] [PubMed]
- Molyva, D.; Kalokasidis, K.; Poulios, C.; Dedi, H.; Karkavelas, G.; Mirtsou, V.; Goulas, A. Rupatadine effectively prevents the histamine-induced up regulation of histamine H1R and bradykinin B2R receptor gene expression in the rat paw. Pharmacol. Rep. 2014, 66, 952–955. [Google Scholar] [CrossRef] [PubMed]
- Mullol, J.; Bousquet, J.; Bachert, C.; Canonica, G.W.; Giménez-Arnau, A.; Kowalski, M.L.; Simons, F.E.R.; Maurer, M.; Ryan, D.; Scadding, G. Update on rupatadine in the management of allergic disorders. Allergy 2015, 70 (Suppl. S100), 1–24. [Google Scholar] [CrossRef] [PubMed]
- Keam, S.J.; Plosker, G.L. Rupatadine: A review of its use in the management of allergic disorders. Drugs 2007, 67, 457–474. [Google Scholar] [CrossRef] [PubMed]
- Nettis, E.; Delle Donne, P.; Di Leo, E.; Calogiuri, G.F.; Ferrannini, A.; Vacca, A. Rupatadine for the treatment of urticaria. Expert Opin. Pharmacother. 2013, 14, 1807–1813. [Google Scholar] [CrossRef]
- Zhang, T.; Li, S.-Y.; Nuermberger, E.L. Autoluminescent Mycobacterium tuberculosis for rapid, real-time, non-invasive assessment of drug and vaccine efficacy. PLoS ONE 2012, 7, e29774. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Njire, M.M.; Liu, J.; Wu, T.; Wang, B.; Liu, T.; Cao, Y.; Liu, Z.; Wan, J.; Tu, Z.; et al. Engineering more stable, selectable marker-free autoluminescent mycobacteria by one step. PLoS ONE 2015, 10, e0119341. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tan, Y.; Islam, M.M.; Cao, Y.; Lu, X.; Zeng, S.; Hameed, H.M.A.; Zhou, P.; Cai, X.; Wang, S.; et al. Assessment of clofazimine and TB47 combination activity against Mycobacterium abscessus using a bioluminescent approach. Antimicrob. Agents Chemother. 2020, 64, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Chiwala, G.; Gao, Y.; Liu, Z.; Sapkota, S.; Lu, Z.; Guo, L.; Khan, S.A.; Zhong, N.; Zhang, T. TB47 and clofazimine form a highly synergistic sterilizing block in a second-line regimen for tuberculosis in mice. Biomed. Pharmacother. 2020, 131, 110782. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, Y.; Liu, J.; Tan, Y.; Liu, Z.; Chhotaray, C.; Jiang, H.; Lu, Z.; Chiwala, G.; Wang, S.; et al. The compound TB47 is highly bactericidal against Mycobacterium ulcerans in a Buruli ulcer mouse model. Nat. Commun. 2019, 10, 524. [Google Scholar] [CrossRef]
- Cho, S.; Lee, H.S.; Franzblau, S. Microplate Alamar Blue assay (MABA) and low oxygen recovery assay (LORA) for Mycobacterium tuberculosis. Methods Mol. Biol. 2015, 1285, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Ma, W.; Yusuf, B.; Li, C.; Hameed, H.M.A.; Wang, X.; Zhong, N.; Hu, J.; Zhang, T. High-throughput screening of compounds against autoluminescent nonreplicating Mycobacterium tuberculosis under diverse conditions. BioRxiv 2024. [Google Scholar] [CrossRef]
- Tyagi, S.; Ammerman, N.C.; Li, S.-Y.; Adamson, J.; Converse, P.J.; Swanson, R.V.; Almeida, D.V.; Grosset, J.H. Clofazimine shortens the duration of the first-line treatment regimen for experimental chemotherapy of tuberculosis. Proc. Natl. Acad. Sci. USA 2015, 112, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Yang, Y.; Tang, Y.; Yang, T.; Sang, Z.; Liu, Z.; Zhang, T.; Luo, Y. Design and synthesis of novel pyrimidine derivatives as potent antitubercular agents. Eur. J. Med. Chem. 2019, 163, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Baindara, P.; Singh, N.; Ranjan, M.; Nallabelli, N.; Chaudhry, V.; Pathania, G.L.; Sharma, N.; Kumar, A.; Patil, P.B.; Korpole, S. Laterosporulin10: A novel defensin like Class IId bacteriocin from Brevibacillus sp. strain SKDU10 with inhibitory activity against microbial pathogens. Microbiology 2016, 162, 1286–1299. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Gao, Y.; Ma, W.; Zhang, J.; Ju, Y.; Ding, J.; Zeng, S.; Hameed, H.M.A.; Zhong, N.; Cook, G.M.; et al. Establishment of an inhalation administration non-invasive murine model for rapidly testing drug activity against Mycobacterium tuberculosis. BioRxiv 2024. [Google Scholar] [CrossRef]
- Makafe, G.G.; Hussain, M.; Surineni, G.; Tan, Y.; Wong, N.-K.; Julius, M.; Liu, L.; Gift, C.; Jiang, H.; Tang, Y.; et al. Quinoline derivatives kill Mycobacterium tuberculosis by activating glutamate kinase. Cell Chem. Biol. 2019, 26, 1187–1194.e5. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, K.A.; Stewart, G.R.; Newton, S.M.; Vordermeier, H.M.; Wain, J.R.; Murphy, H.N.; Horner, K.; Young, D.B.; Wilkinson, R.J. Infection biology of a novel alpha-crystallin of Mycobacterium tuberculosis: Acr2. J. Immunol. 2005, 174, 4237–4243. [Google Scholar] [CrossRef] [PubMed]
- Healy, C.; Golby, P.; MacHugh, D.E.; Gordon, S.V. The MarR family transcription factor Rv1404 coordinates adaptation of Mycobacterium tuberculosis to acid stress via controlled expression of Rv1405c, a virulence-associated methyltransferase. Tuberculosis 2016, 97, 154–162. [Google Scholar] [CrossRef]
- Tükenmez, H.; Sarkar, S.; Anoosheh, S.; Kruchanova, A.; Edström, I.; Harrison, G.A.; Stallings, C.L.; Almqvist, F.; Larsson, C. Mycobacterium tuberculosis Rv3160c is a TetR-like transcriptional repressor that regulates expression of the putative oxygenase Rv3161c. Sci. Rep. 2021, 11, 1523. [Google Scholar] [CrossRef]
- Larsen, M.H.; Lacourciere, K.; Parker, T.M.; Kraigsley, A.; Achkar, J.M.; Adams, L.B.; Dupnik, K.M.; Hall-Stoodley, L.; Hartman, T.; Kanipe, C.; et al. The many hosts of mycobacteria 8 (MHM8): A conference report. Tuberculosis 2020, 121, 101914. [Google Scholar] [CrossRef]
- Togre, N.S.; Vargas, A.M.; Bhargavi, G.; Mallakuntla, M.K.; Tiwari, S. Fragment-based drug discovery against mycobacteria: The success and challenges. Int. J. Mol. Sci. 2022, 23, 10669. [Google Scholar] [CrossRef]
- Kim, W.S.; Kim, J.-S.; Kim, H.M.; Kwon, K.W.; Eum, S.-Y.; Shin, S.J. Comparison of immunogenicity and vaccine efficacy between heat-shock proteins, HSP70 and GrpE, in the DnaK operon of Mycobacterium tuberculosis. Sci. Rep. 2018, 8, 14411. [Google Scholar] [CrossRef]
- Trutneva, K.A.; Shleeva, M.O.; Demina, G.R.; Vostroknutova, G.N.; Kaprelyans, A.S. One-year old dormant, “Non-culturable” Mycobacterium tuberculosis preserves significantly diverse protein profile. Front. Cell Infect. Microbiol. 2020, 10, 26. [Google Scholar] [CrossRef]
- Tripathi, P.; Singh, L.K.; Kumari, S.; Hakiem, O.R.; Batra, J.K. ClpB is an essential stress regulator of Mycobacterium tuberculosis and endows survival advantage to dormant bacilli. Int. J. Med. Microbiol. 2020, 310, 151402. [Google Scholar] [CrossRef]
- Chatterjee, C.; Majumdar, S.; Deshpande, S.; Pant, D.; Matheshwaran, S. Real-time kinetic studies of Mycobacterium tuberculosis LexA-DNA interaction. Biosci. Rep. 2021, 41, BSR20211419. [Google Scholar] [CrossRef]
- Chandran, A.V.; Srikalaivani, R.; Paul, A.; Vijayan, M. Biochemical characterization of Mycobacterium tuberculosis LexA and structural studies of its C-terminal segment. Acta Crystallogr. Sect. D Struct. Biol. 2019, 75, 41–55. [Google Scholar] [CrossRef]
- Beck, I.N.; Usher, B.; Hampton, H.G.; Fineran, P.C.; Blower, T.R. Antitoxin autoregulation of M. tuberculosis toxin-antitoxin expression through negative cooperativity arising from multiple inverted repeat sequences. Biochem. J. 2020, 477, 2401–2419. [Google Scholar] [CrossRef]
- Janowski, R.; Panjikar, S.; Eddine, A.N.; Kaufmann, S.H.E.; Weiss, M.S. Structural analysis reveals DNA binding properties of Rv2827c, a hypothetical protein from Mycobacterium tuberculosis. J. Struct. Funct. Genom. 2009, 10, 137–150. [Google Scholar] [CrossRef]
- Konar, M.; Alam, M.S.; Arora, C.; Agrawal, P. WhiB2/Rv3260c, a cell division-associated protein of Mycobacterium tuberculosis H37Rv, has properties of a chaperone. FEBS J. 2012, 279, 2781–2792. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains # | MICs | |
|---|---|---|
| μg/mL | μM | |
| AlRa | 3.13 | 7.52 |
| AlRv | 3.13 | 7.52 |
| AlMm | 3.13 | 7.52 |
| AlMab | 50 | 120.20 |
| AlMs | 12.5 | 30.05 |
| Strains # | MIC | ||
|---|---|---|---|
| μg/mL | μM | ||
| Replicating | AlRa G | 3.13 | 7.52 |
| AlRa C | 1.56 | 3.75 | |
| Nonreplicating | AlRa G | 25 | 60.10 |
| AlRa C | 50 | 120.20 | |
| Drug * | MIC # (µg/mL) | FICI ※ | Effects | |||
|---|---|---|---|---|---|---|
| MIC RTD C | MIC RTD A | MIC C | MIC A | |||
| AMK | 0.20 | 3.13 | 0.5 | 0.5 | 1.0625 | Indifferent |
| CLO | 0.78 | 3.13 | 0.1 | 0.2 | 0.75 | Partial synergistic |
| INH | 3.13 | 3.13 | 0.003125 | 0.05 | 1.0625 | Indifferent |
| LEV | 0.20 | 3.13 | 0.125 | 0.125 | 1.0625 | Indifferent |
| LZD | 3.13 | 3.13 | 0.5 | 1 | 1.5 | Indifferent |
| PTM | 0.20 | 3.13 | 0.0625 | 0.125 | 0.5625 | Partial synergistic |
| RIF | 3.13 | 3.13 | 0.0015625 | 0.0125 | 1.125 | Indifferent |
| STR | 1.56 | 3.13 | 2 | 4 | 1 | Additive |
| TB47 | 0.78 | 3.13 | 0.00075 | 0.0015 | 0.75 | Partial synergistic |
| Gene_Id | log2FoldChange * | Gene_Name | Product/Function |
|---|---|---|---|
| Rv0350 | 3.68124885 | dnaK | Probable chaperone protein DnaK (heat shock protein 70); acts as a chaperone. |
| Rv0351 | 3.750319008 | grpE | Probable GrpE protein (HSP-70 cofactor); stimulates, jointly with Rv0352, the ATPase activity of Rv0350. |
| Rv0352 | 3.066383388 | dnaJ1 | Chaperone protein DnaJ (HSP-70 cofactor); stimulates, jointly with Rv0351, the ATPase activity of Rv0350. |
| Rv0384c | 2.667760091 | clpB | Probable endopeptidase ATP binding protein ClpB (heat shock protein F84.1); thought to be an ATPase subunit of an intracellular ATP-dependent protease. |
| Rv0440 | 2.448531361 | groEL2 | Molecular chaperone GroEL; prevents misfolding and promotes the refolding and proper assembly of unfolded polypeptides generated under stress conditions. |
| Rv2720 | 2.27206078 | lexA | Repressor LexA; involved in regulation of nucleotide excision repair and sos response. |
| Rv2827c | 2.142454829 | Rv2827c | Hypothetical protein; unknown. |
| Rv3260c | -2.134883679 | whiB2 | Probable transcriptional regulatory protein WhiB-like WhiB2; involved in transcriptional mechanism. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, X.; Ma, W.; Yusuf, B.; Su, B.; Hu, J.; Zhang, T. Assessment of the Efficacy of the Antihistamine Drug Rupatadine Used Alone or in Combination against Mycobacteria. Pharmaceutics 2024, 16, 1049. https://doi.org/10.3390/pharmaceutics16081049
Tian X, Ma W, Yusuf B, Su B, Hu J, Zhang T. Assessment of the Efficacy of the Antihistamine Drug Rupatadine Used Alone or in Combination against Mycobacteria. Pharmaceutics. 2024; 16(8):1049. https://doi.org/10.3390/pharmaceutics16081049
Chicago/Turabian StyleTian, Xirong, Wanli Ma, Buhari Yusuf, Biyi Su, Jinxing Hu, and Tianyu Zhang. 2024. "Assessment of the Efficacy of the Antihistamine Drug Rupatadine Used Alone or in Combination against Mycobacteria" Pharmaceutics 16, no. 8: 1049. https://doi.org/10.3390/pharmaceutics16081049