

Butyrate Producers in Very Low Birth Weight Infants with Neither Culture-Proven Sepsis nor Necrotizing Enterocolitis
 , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design and Pateint Domains
2.2. Feeding Care
2.3. Outcome and Definitions
2.4. Fecal Samples for DNA Extraction
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thatrimontrichai, A.; Chanvitan, P.; Janjindamai, W.; Dissaneevate, S.; Jefferies, A.; Shah, V. Trends in neonatal sepsis in a neonatal intensive care unit in Thailand before and after construction of a new facility. Asian Biomed 2014, 8, 771–778. [Google Scholar] [CrossRef]
- Masi, A.C.; Stewart, C.J. The role of the preterm intestinal microbiome in sepsis and necrotising enterocolitis. Early Hum. Dev. 2019, 138, 104854. [Google Scholar] [CrossRef] [PubMed]
- Thatrimontrichai, A.; Tonjit, P.; Janjindamai, W.; Dissaneevate, S.; Maneenil, G.; Phatigomet, M. Risk factors associated with 30-day mortality among neonates with A. baumannii sepsis. Pediatr. Infect. Dis. J. 2021, 40, 1111–1114. [Google Scholar] [CrossRef]
- Gerios, L.; Rodrigues, V.H.B.; Corrente, J.E.; Lyra, J.C.; Rugolo, L.; Bentlin, M.R. Late-onset sepsis in very low birth weight premature infants: A 10-year review of a Brazilian tertiary university hospital-the challenge remains. Am. J. Perinatol. 2024, 41, e1725–e1731. [Google Scholar] [CrossRef]
- Stoll, B.J.; Hansen, N.; Fanaroff, A.A.; Wright, L.L.; Carlo, W.A.; Ehrenkranz, R.A.; Lemons, J.A.; Donovan, E.F.; Stark, A.R.; Tyson, J.E.; et al. Late-onset sepsis in very low birth weight neonates: The experience of the NICHD Neonatal Research Network. Pediatrics 2002, 110, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.Y.; Chang, Y.H.; Lee, C.C. The impact of gut microbiota on morbidities in preterm infants. Kaohsiung J. Med. Sci. 2024, 40, 780–788. [Google Scholar] [CrossRef]
- Kim, C.S.; Claud, E.C. Necrotizing enterocolitis pathophysiology: How microbiome data alter our understanding. Clin. Perinatol. 2019, 46, 29–38. [Google Scholar] [CrossRef]
- Thänert, R.; Keen, E.C.; Dantas, G.; Warner, B.B.; Tarr, P.I. Necrotizing enterocolitis and the microbiome: Current status and future directions. J. Infect. Dis. 2021, 223, S257–S263. [Google Scholar] [CrossRef]
- Fu, X.; Li, S.; Jiang, Y.; Hu, X.; Wu, H. Necrotizing enterocolitis and intestinal microbiota: The timing of disease and combined effects of multiple species. Front. Pediatr. 2021, 9, 657349. [Google Scholar] [CrossRef]
- Liu, J.; Li, Y.; Feng, Y.; Pan, L.; Xie, Z.; Yan, Z.; Zhang, L.; Li, M.; Zhao, J.; Sun, J.; et al. Patterned progression of gut microbiota associated with necrotizing enterocolitis and late onset sepsis in preterm infants: A prospective study in a Chinese neonatal intensive care unit. PeerJ 2019, 7, e7310. [Google Scholar] [CrossRef]
- Stewart, C.J.; Embleton, N.D.; Marrs, E.C.L.; Smith, D.P.; Fofanova, T.; Nelson, A.; Skeath, T.; Perry, J.D.; Petrosino, J.F.; Berrington, J.E.; et al. Longitudinal development of the gut microbiome and metabolome in preterm neonates with late onset sepsis and healthy controls. Microbiome 2017, 5, 75. [Google Scholar] [CrossRef] [PubMed]
- Heston, S.M.; Lim, C.S.E.; Ong, C.; Chua, M.C.; Kelly, M.S.; Yeo, K.T. Strain-resolved metagenomic analysis of the gut as a reservoir for bloodstream infection pathogens among premature infants in Singapore. Gut Pathog. 2023, 15, 55. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, D.J.; Shalon, N.; Wardenburg, K.; DeVeaux, A.; Wallace, M.A.; Hall-Moore, C.; Ndao, I.M.; Sullivan, J.E.; Radmacher, P.; Escobedo, M.; et al. Gut pathogen colonization precedes bloodstream infection in the neonatal intensive care unit. Sci. Transl. Med. 2023, 15, 5562. [Google Scholar] [CrossRef]
- Phatigomet, M.; Thatrimontrichai, A.; Maneenil, G.; Dissaneevate, S.; Janjindamai, W. Risk factors for 30-day mortality in neonates with carbapenem-resistant A. baumannii sepsis. Pediatr. Infect. Dis. J. 2022, 41, 1012–1016. [Google Scholar] [CrossRef]
- Thatrimontrichai, A.; Surachat, K.; Singkhamanan, K.; Thongsuksai, P. Differential abundances of Bdellovibrio and Rheinheimera in the oral microbiota of neonates with and without clinical sepsis. Pediatr. Infect. Dis. J. 2024, 43, e195–e200. [Google Scholar] [CrossRef] [PubMed]
- Thatrimontrichai, A.; Surachat, K.; Singkhamanan, K.; Thongsuksai, P. Long duration of oral care using mother’s own milk influences oral microbiota and clinical outcomes in very-low-birthweight infants: Randomized controlled trial. Pediatr. Infect. Dis. J. 2023, 42, 804–810. [Google Scholar] [CrossRef]
- Bell, M.J.; Ternberg, J.L.; Feigin, R.D.; Keating, J.P.; Marshall, R.; Barton, L.; Brotherton, T. Neonatal necrotizing enterocolitis. Therapeutic decisions based upon clinical staging. Ann. Surg. 1978, 187, 1–7. [Google Scholar] [CrossRef]
- Levy, M.M.; Fink, M.P.; Marshall, J.C.; Abraham, E.; Angus, D.; Cook, D.; Cohen, J.; Opal, S.M.; Vincent, J.L.; Ramsay, G.; et al. 2001 SCCM/ESICM/ACCP/ATS/SIS international sepsis definitions conference. Intensive Care Med. 2003, 29, 530–538. [Google Scholar] [CrossRef]
- Lee, J.; Kim, H.S.; Jung, Y.H.; Choi, K.Y.; Shin, S.H.; Kim, E.K.; Choi, J.H. Oropharyngeal colostrum administration in extremely premature infants: An RCT. Pediatrics 2015, 135, e357–e366. [Google Scholar] [CrossRef]
- Jobe, A.H.; Bancalari, E. Bronchopulmonary dysplasia. Am. J. Respir. Crit. Care Med. 2001, 163, 1723–1729. [Google Scholar] [CrossRef]
- ZymoBIOMICS™ DNA Miniprep Kit. Available online: https://files.zymoresearch.com/protocols/_d4300t_d4300_d4304_zymobiomics_dna_miniprep_kit.pdf (accessed on 3 July 2017).
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.M.; Lin, C.P.; Jan, M.S. Early gut microbiota changes in preterm infants with bronchopulmonary dysplasia: A pilot case-control study. Am. J. Perinatol. 2021, 38, 1142–1149. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, R.; Tsuji, H.; Takahashi, T.; Nomoto, K.; Kawashima, K.; Nagata, S.; Yamashiro, Y. Ontogenesis of the gut microbiota composition in healthy, full-term, vaginally born and breast-fed infants over the first 3 years of life: A quantitative bird’s-eye view. Front. Microbiol. 2017, 8, 1388. [Google Scholar] [CrossRef] [PubMed]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef]
- Singh, V.; Lee, G.; Son, H.; Koh, H.; Kim, E.S.; Unno, T.; Shin, J.H. Butyrate producers, “The Sentinel of Gut”: Their intestinal significance with and beyond butyrate, and prospective use as microbial therapeutics. Front. Microbiol. 2022, 13, 1103836. [Google Scholar] [CrossRef]
- Round, J.L.; Lee, S.M.; Li, J.; Tran, G.; Jabri, B.; Chatila, T.A.; Mazmanian, S.K. The Toll-like receptor 2 pathway establishes colonization by a commensal of the human microbiota. Science 2011, 332, 974–977. [Google Scholar] [CrossRef]
- Seki, H.; Shiohara, M.; Matsumura, T.; Miyagawa, N.; Tanaka, M.; Komiyama, A.; Kurata, S. Prevention of antibiotic-associated diarrhea in children by Clostridium butyricum MIYAIRI. Pediatr. Int. 2003, 45, 86–90. [Google Scholar] [CrossRef]
- Fukushima, K.; Kudo, H.; Oka, K.; Hayashi, A.; Onizuka, M.; Kusakabe, S.; Hino, A.; Takahashi, M.; Takeda, K.; Mori, M.; et al. Clostridium butyricum MIYAIRI 588 contributes to the maintenance of intestinal microbiota diversity early after haematopoietic cell transplantation. Bone Marrow Transplant. 2024, 59, 795–802. [Google Scholar] [CrossRef]
- Sada, R.M.; Matsuo, H.; Motooka, D.; Kutsuna, S.; Hamaguchi, S.; Yamamoto, G.; Ueda, A. Clostridium butyricum bacteremia associated with probiotic use, Japan. Emerg. Infect. Dis. 2024, 30, 665–671. [Google Scholar] [CrossRef]
- Li, X.; Hu, S.; Yin, J.; Peng, X.; King, L.; Li, L.; Xu, Z.; Zhou, L.; Peng, Z.; Ze, X.; et al. Effect of synbiotic supplementation on immune parameters and gut microbiota in healthy adults: A double-blind randomized controlled trial. Gut Microbes 2023, 15, 2247025. [Google Scholar] [CrossRef]
- Yang, R.; Wang, H.; Chen, D.; Cai, Q.; Zhu, J.; Yuan, S.; Wang, F.; Xu, X. The effect of in-hospital breast milk intake on the gut microbiota of preterm infants. Clin. Nutr. ESPEN 2024, 60, 146–155. [Google Scholar] [CrossRef] [PubMed]


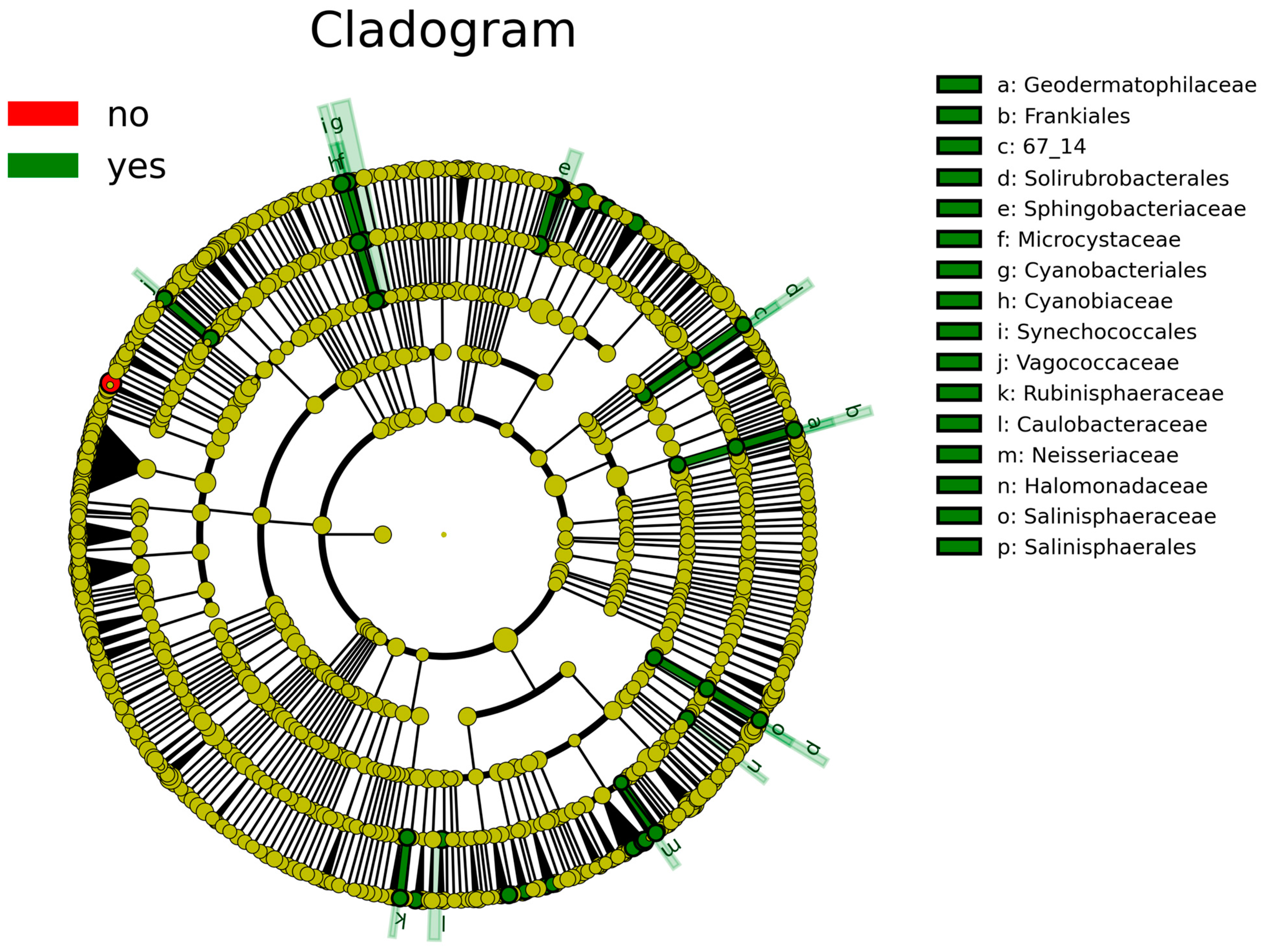

{kind=link}
{kind=link}
{kind=link}
| Baseline Characteristics | sInfection (n = 9) | Non-sInfection (n = 42) | p-Value |
|---|---|---|---|
| Gestational age, weeks * | 27.3 ± 1.9 | 28.9 ± 2.2 | 0.06 |
| Birth weight, g * | 882 ± 229 | 1069 ± 266 | 0.06 |
| Male, n (%) | 3 (33.3) | 25 (59.5) | 0.27 |
| Appropriate for gestational age, n (%) | 8 (88.9) | 32 (76.2) | 0.66 |
| Cesarean delivery, n (%) | 7 (77.8) | 29 (69.0) | 0.71 |
| Premature rupture of membranes, n (%) | 2 (22.2) | 7 (16.7) | 0.09 |
| Maternal antibiotics, n (%) | 2 (22.2) | 11 (26.2) | >0.99 |
| Antenatal steroid, n (%) | 7 (77.8) | 27 (64.3) | 0.70 |
| 1 min Apgar score † | 5 (4–6) | 6 (3–8) | 0.28 |
| 5 min Apgar score † | 8 (5–8) | 8 (7–9) | 0.27 |
| Number of endotracheal intubations, n (%) | 8 (88.9) | 30 (71.4) | 0.42 |
| Duration of invasive ventilation, days † | 5 (2–11) | 1 (0–3) | 0.01 |
| Duration of non-invasive ventilation, days † | 38 (28–51) | 11 (7–36) | 0.02 |
| Respiratory distress syndrome, n (%) | 6 (66.7) | 36 (85.7) | 0.19 |
| Surfactant administration, n (%) | 5 (55.6) | 17 (40.5) | 0.47 |
| Patent ductus arteriosus, n (%) | 4 (44.4) | 13 (31.0) | 0.46 |
| Moderate to severe bronchopulmonary dysplasia, n (%) | 4 (44.4) | 5 (11.9) | 0.04 |
| Duration of antibiotic use, days † | 23 (18–26) | 6 (3–12) | 0.001 |
| Date of full enteral feeding, days † | 27 (23–32) | 12 (7–17) | <0.001 |
| Date of regained birth weight, days * | 9.0 ± 4.1 | 8.3 ± 3.6 | 0.60 |
| Oral care by mother’s own milk, n (%) | 5 (55.6) | 20 (47.6) | 0.73 |
| Percentage volume of enteral breast milk † | 30 (4–94) | 93 (68–99) | 0.08 |
| Alpha Diversity | sInfection (n = 9) | Non-sInfection (n = 42) | p-Value | p-Value * |
|---|---|---|---|---|
| Shannon diversity a | 3.2 (2.4–3.7) | 3.4 (3.1–4.2) | 0.19 | 0.28 |
| Observed using operational taxonomic units a | 34 (31–42) | 49 (39–161) | 0.06 | 0.74 |
| Faith’s phylogenetic diversity a | 9.0 (7.5–12.7) | 8.4 (7.6–30.6) | 0.49 | 0.59 |
| Pielou’s evenness b | 0.6 ± 0.1 | 0.6 ± 0.1 | 0.50 | 0.33 |
| Beta diversity | Permutations | pseudo-F | p-Value | q-Value ** |
| Unweighted UniFrac distance | 999 | 1.06 | 0.29 | 0.29 |
| Weighted UniFrac distance | 999 | 1.81 | 0.08 | 0.08 |
| Taxa | Bacteria | sInfection (n = 9) | Non-sInfection (n = 42) |
|---|---|---|---|
| Phyla | Proteobacteria | 52.9% | 59.2% |
| Firmicutes | 30.2% | 30.9% | |
| Bacteroidota | 15.5% | 6.8% | |
| Actinobacteriota | 1.4% | 2.6% | |
| Classes | Gammaproteobacteria | 52.8% | 58.8% |
| Bacilli | 25.6% | 10.5% | |
| Bacteroidia | 15.5% | 6.8% | |
| Clostridia | 3.1% | 14.9% | |
| Negativicutes | 1.6% | 5.1% | |
| Actinobacteria | 1.4% | 2.6% | |
| Genera | Klebsiella | 15.8% | 23.0% |
| Escherichia-Shigella | 8.3% | 13.7% | |
| unclassified Enterobacteriaceae | 14.1% | 11.5% | |
| Enterococcus | 22.5% | 4.1% | |
| Faecalibacterium | 0% | 5.8% | |
| Bacteroides | 0% | 5.8% | |
| Acinetobacter | 0.8% | 5.1% | |
| Clostridium_sensu_stricto_1 | 2.7% | 4.5% | |
| Veillonella | 1.6% | 4.8% | |
| Serratia | 9.0% | 2.2% | |
| Elizabethkingia | 15.3% | 0% | |
| Streptococcus | 0.3% | 3.1% | |
| Bifidobacterium | 0.3% | 2.4% | |
| Enterobacter | 2.1% | 1.8% | |
| Staphylococcus | 2.7% | 1.2% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thatrimontrichai, A.; Praditaukrit, M.; Maneenil, G.; Dissaneevate, S.; Singkhamanan, K.; Surachat, K. Butyrate Producers in Very Low Birth Weight Infants with Neither Culture-Proven Sepsis nor Necrotizing Enterocolitis. Nutrients 2025, 17, 1329. https://doi.org/10.3390/nu17081329
Thatrimontrichai A, Praditaukrit M, Maneenil G, Dissaneevate S, Singkhamanan K, Surachat K. Butyrate Producers in Very Low Birth Weight Infants with Neither Culture-Proven Sepsis nor Necrotizing Enterocolitis. Nutrients. 2025; 17(8):1329. https://doi.org/10.3390/nu17081329
Chicago/Turabian StyleThatrimontrichai, Anucha, Manapat Praditaukrit, Gunlawadee Maneenil, Supaporn Dissaneevate, Kamonnut Singkhamanan, and Komwit Surachat. 2025. "Butyrate Producers in Very Low Birth Weight Infants with Neither Culture-Proven Sepsis nor Necrotizing Enterocolitis" Nutrients 17, no. 8: 1329. https://doi.org/10.3390/nu17081329
APA StyleThatrimontrichai, A., Praditaukrit, M., Maneenil, G., Dissaneevate, S., Singkhamanan, K., & Surachat, K. (2025). Butyrate Producers in Very Low Birth Weight Infants with Neither Culture-Proven Sepsis nor Necrotizing Enterocolitis. Nutrients, 17(8), 1329. https://doi.org/10.3390/nu17081329