
Deciphering the Non-Coding RNA Landscape of Pediatric Acute Myeloid Leukemia

 ,
,  , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Controls
2.2. Cell Sorting
2.3. RNA Isolation
2.4. Micro-Array Analysis
2.5. Small RNA-Sequencing Analysis
2.6. miRNA-lncRNA Network Constuction
2.7. Gene Set Enrichment Analysis
3. Results
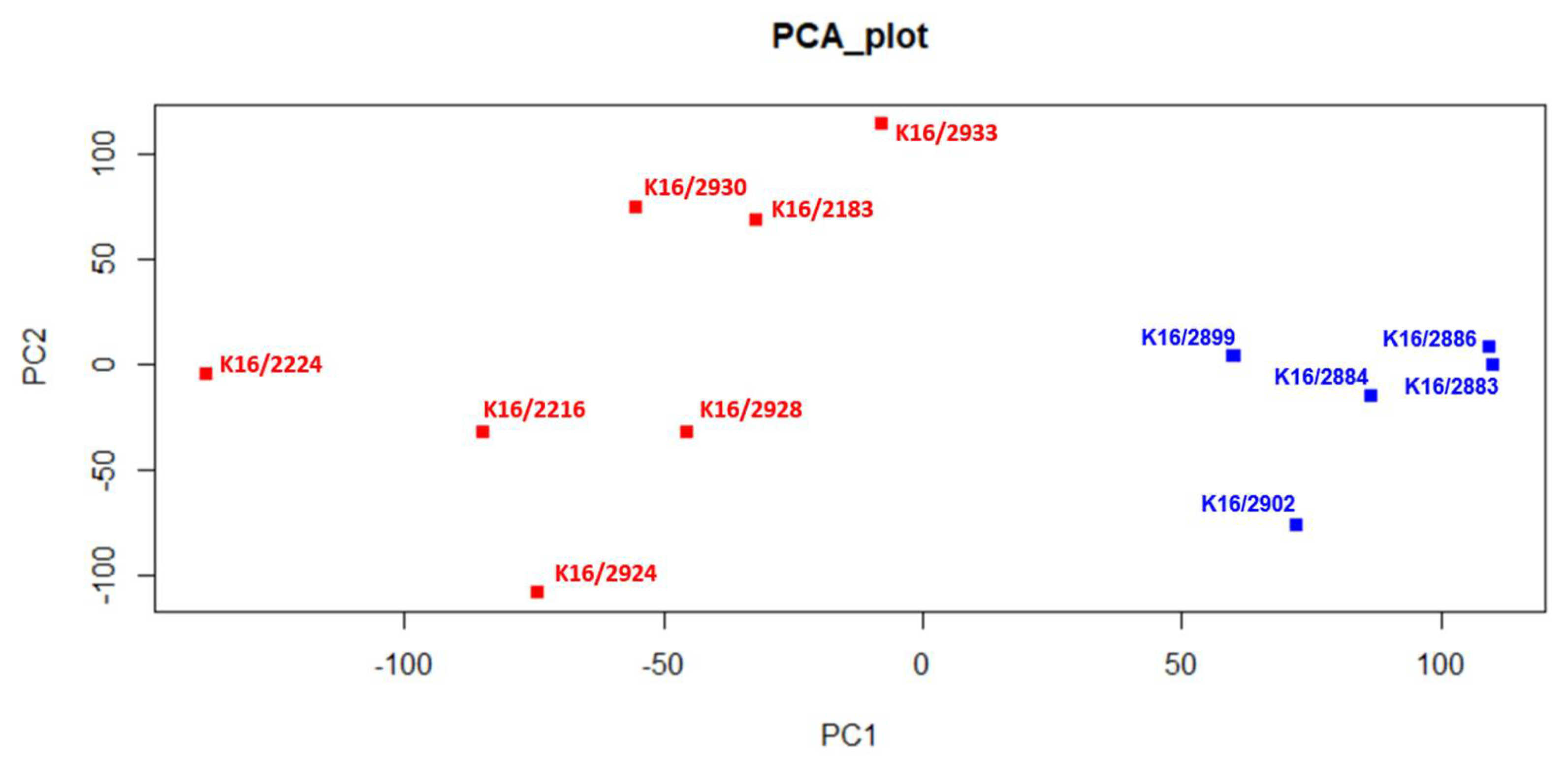
3.1. Identification of Differentially Expressed lncRNAs in pedAML Subpopulations
3.2. Identification of Potential Functional Networks
3.3. Identification of Potential Functional Relevance of Unique LSC DE-lncRNAs and Shared DE-lncRNAs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rasche, M.; Zimmermann, M.; Borschel, L.; Bourquin, J.-P.; Dworzak, M.; Klingebiel, T.; Lehrnbecher, T.; Creutzig, U.; Klusmann, J.-H.; Reinhardt, D. Successes and Challenges in the Treatment of Pediatric Acute Myeloid Leukemia: A Retrospective Analysis of the AML-BFM Trials from 1987 to 2012. Leukemia 2018, 32, 2167–2177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Moerloose, B.; Reedijk, A.; de Bock, G.H.; Lammens, T.; de Haas, V.; Denys, B.; Dedeken, L.; van den Heuvel-Eibrink, M.M.; Te Loo, M.; Uyttebroeck, A.; et al. Response-Guided Chemotherapy for Pediatric Acute Myeloid Leukemia without Hematopoietic Stem Cell Transplantation in First Complete Remission: Results from Protocol DB AML-01. Pediatr. Blood Cancer 2019, 66, e27605. [Google Scholar] [CrossRef]
- Zwaan, C.M.; Kolb, E.A.; Reinhardt, D.; Abrahamsson, J.; Adachi, S.; Aplenc, R.; De Bont, E.S.J.M.; De Moerloose, B.; Dworzak, M.; Gibson, B.E.S.; et al. Collaborative Efforts Driving Progress in Pediatric Acute Myeloid Leukemia. J. Clin. Oncol. 2015, 33, 2949–2962. [Google Scholar] [CrossRef] [Green Version]
- Depreter, B.; Weening, K.E.; Vandepoele, K.; Essand, M.; De Moerloose, B.; Themeli, M.; Cloos, J.; Hanekamp, D.; Moors, I.; D’hont, I.; et al. TARP Is an Immunotherapeutic Target in Acute Myeloid Leukemia Expressed in the Leukemic Stem Cell Compartment. Haematologica 2020, 105, 1306–1316. [Google Scholar] [CrossRef]
- Depreter, B.; De Moerloose, B.; Vandepoele, K.; Uyttebroeck, A.; Van Damme, A.; Denys, B.; Dedeken, L.; Dresse, M.-F.; Van der Werff Ten Bosch, J.; Hofmans, M.; et al. Clinical Significance of TARP Expression in Pediatric Acute Myeloid Leukemia. Hemasphere 2020, 4, e346. [Google Scholar] [CrossRef]
- Wang, J.; Chen, S.; Xiao, W.; Li, W.; Wang, L.; Yang, S.; Wang, W.; Xu, L.; Liao, S.; Liu, W.; et al. CAR-T Cells Targeting CLL-1 as an Approach to Treat Acute Myeloid Leukemia. J. Hematol. Oncol. 2018, 11, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapuis, A.G.; Egan, D.N.; Bar, M.; Schmitt, T.M.; McAfee, M.S.; Paulson, K.G.; Voillet, V.; Gottardo, R.; Ragnarsson, G.B.; Bleakley, M.; et al. T Cell Receptor Gene Therapy Targeting WT1 Prevents Acute Myeloid Leukemia Relapse Post-Transplant. Nat. Med. 2019, 25, 1064–1072. [Google Scholar] [CrossRef]
- Taussig, D.C.; Pearce, D.J.; Simpson, C.; Rohatiner, A.Z.; Lister, T.A.; Kelly, G.; Luongo, J.L.; Danet-Desnoyers, G.-A.H.; Bonnet, D. Hematopoietic Stem Cells Express Multiple Myeloid Markers: Implications for the Origin and Targeted Therapy of Acute Myeloid Leukemia. Blood 2005, 106, 4086–4092. [Google Scholar] [CrossRef] [Green Version]
- Cai, S.F.; Levine, R.L. Genetic and Epigenetic Determinants of AML Pathogenesis. Semin. Hematol. 2019, 56, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Bill, M.; Papaioannou, D.; Karunasiri, M.; Kohlschmidt, J.; Pepe, F.; Walker, C.J.; Walker, A.E.; Brannan, Z.; Pathmanathan, A.; Zhang, X.; et al. Expression and Functional Relevance of Long Non-Coding RNAs in Acute Myeloid Leukemia Stem Cells. Leukemia 2019, 33, 2169–2182. [Google Scholar] [CrossRef]
- Schwarzer, A.; Emmrich, S.; Schmidt, F.; Beck, D.; Ng, M.; Reimer, C.; Adams, F.F.; Grasedieck, S.; Witte, D.; Käbler, S.; et al. The Non-Coding RNA Landscape of Human Hematopoiesis and Leukemia. Nat. Commun. 2017, 8, 218. [Google Scholar] [CrossRef] [PubMed]
- Bhan, A.; Soleimani, M.; Mandal, S.S. Long Noncoding RNA and Cancer: A New Paradigm. Cancer Res. 2017, 77, 3965–3981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhari, R.; Sedano, M.J.; Harrison, A.L.; Subramani, R.; Lin, K.Y.; Ramos, E.I.; Lakshmanaswamy, R.; Gadad, S.S. Long Noncoding RNAs in Cancer: From Discovery to Therapeutic Targets. Adv. Clin. Chem. 2020, 95, 105–147. [Google Scholar] [CrossRef] [PubMed]
- Geisler, S.; Coller, J. RNA in Unexpected Places: Long Non-Coding RNA Functions in Diverse Cellular Contexts. Nat. Rev. Mol. Cell Biol. 2013, 14, 699–712. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wang, J. LncRNA TUG1 Regulates Cell Viability and Death by Regulating MiR-193a-5p/Rab10 Axis in Acute Myeloid Leukemia. OncoTargets Ther. 2020, 13, 1289–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, M.-F.; Li, L.-J.; Ma, J.-B. LncRNA HOTTIP Promotes Proliferation and Cell Cycle Progression of Acute Myeloid Leukemia Cells. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 2908–2915. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.; Lu, P.; Guan, J.; Zhou, Y.; Zou, L.; Yi, X.; Cheng, H. LncRNA KCNQ1OT1 Controls Cell Proliferation, Differentiation and Apoptosis by Sponging MiR-326 to Regulate c-Myc Expression in Acute Myeloid Leukemia. Neoplasma 2020, 67, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Guo, R.; Yuan, Z.; Shi, H.; Zhang, D. LncRNA HOTAIR Regulates CCND1 and CCND2 Expression by Sponging MiR-206 in Ovarian Cancer. Cell Physiol. Biochem. 2018, 49, 1289–1303. [Google Scholar] [CrossRef]
- Depreter, B.; De Moerloose, B.; Vandepoele, K.; Uyttebroeck, A.; Van Damme, A.; Terras, E.; Denys, B.; Dedeken, L.; Dresse, M.-F.; Van der Werff Ten Bosch, J.; et al. Deciphering Molecular Heterogeneity in Pediatric AML Using a Cancer vs. Normal Transcriptomic Approach. Pediatr. Res. 2021, 89, 1695–1705. [Google Scholar] [CrossRef] [PubMed]
- Volders, P.-J.; Anckaert, J.; Verheggen, K.; Nuytens, J.; Martens, L.; Mestdagh, P.; Vandesompele, J. LNCipedia 5: Towards a Reference Set of Human Long Non-Coding RNAs. Nucleic Acids Res. 2019, 47, D135–D139. [Google Scholar] [CrossRef] [Green Version]
- Dewaele, S.; Delhaye, L.; De Paepe, B.; de Bony, E.J.; De Wilde, J.; Vanderheyden, K.; Anckaert, J.; Yigit, N.; Nuytens, J.; Vanden Eynde, E.; et al. The Long Non-Coding RNA SAMMSON Is Essential for Uveal Melanoma Cell Survival. Oncogene 2021, 41, 15–25. [Google Scholar] [CrossRef]
- Hofmans, M.; Lammens, T.; Depreter, B.; Wu, Y.; Erlacher, M.; Caye, A.; Cavé, H.; Flotho, C.; de Haas, V.; Niemeyer, C.M.; et al. Long Non-Coding RNAs as Novel Therapeutic Targets in Juvenile Myelomonocytic Leukemia. Sci. Rep. 2021, 11, 2801. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Beck, D.; Thoms, J.A.I.; Liu, L.; Zhao, W.; Pimanda, J.E.; Zhou, X. Annotating Function to Differentially Expressed LincRNAs in Myelodysplastic Syndrome Using a Network-Based Method. Bioinformatics 2017, 33, 2622–2630. [Google Scholar] [CrossRef] [PubMed]
- Montalban-Bravo, G.; Kanagal-Shamanna, R.; Class, C.A.; Sasaki, K.; Ravandi, F.; Cortes, J.E.; Daver, N.; Takahashi, K.; Short, N.J.; DiNardo, C.D.; et al. Outcomes of Acute Myeloid Leukemia with Myelodysplasia Related Changes Depend on Diagnostic Criteria and Therapy. Am. J. Hematol. 2020, 95, 612–622. [Google Scholar] [CrossRef]
- Li, Y.; Kong, C.; Wu, C.; Wang, Y.; Xu, B.; Liang, S.; Ying, X. Knocking down of LINC01220 Inhibits Proliferation and Induces Apoptosis of Endometrial Carcinoma through Silencing MAPK11. Biosci. Rep. 2019, 39, BSR20181794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, C.; Gao, Y.-Y.; Ju, Q.-Q.; Zhang, C.-X.; Gong, M.; Li, Z.-L. LINC00649 Underexpression Is an Adverse Prognostic Marker in Acute Myeloid Leukemia. BMC Cancer 2020, 20, 841. [Google Scholar] [CrossRef]
- Chen, X.; Chen, S. LINC00649 Promotes Bladder Cancer Malignant Progression by Regulating the MiR-15a-5p/HMGA1 Axis. Oncol. Rep. 2021, 45, 8. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Di, X.; Bi, Y.; Sun, S.; Wang, T. Long Non-Coding RNA LINC00649 Regulates YES-Associated Protein 1 (YAP1)/Hippo Pathway to Accelerate Gastric Cancer (GC) Progression via Sequestering MiR-16-5p. Bioengineered 2021, 12, 1791–1802. [Google Scholar] [CrossRef] [PubMed]
- Ye, G.; Guo, L.; Xing, Y.; Sun, W.; Yuan, M. Identification of Prognostic Biomarkers of Prostate Cancer with Long Non-Coding RNA-Mediated Competitive Endogenous RNA Network. Exp. Ther. Med. 2019, 17, 3035–3040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Gu, Y.; Chen, S.; Tian, Y.; Yang, S. Hsa_circ_0079480 Promotes Tumor Progression in Acute Myeloid Leukemia via MiR-654-3p/HDGF Axis. Aging 2020, 13, 1120–1131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Li, Q.; Zhu, K.; Zhu, J.; Li, J.; Yuan, Y.; Zhang, P.; Zhou, L.; Liu, L. LncRNA LINC00265/MiR-485-5p/IRF2-Mediated Autophagy Suppresses Apoptosis in Acute Myeloid Leukemia Cells. Am. J. Transl. Res. 2020, 12, 2451–2462. [Google Scholar] [PubMed]
- Liu, W.; Cheng, F. Circular RNA CircCRKL Inhibits the Proliferation of Acute Myeloid Leukemia Cells via the MiR-196a-5p/MiR-196b-5p/P27 Axis. Bioengineered 2021, 12, 7704–7713. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Chen, Y.; Zhu, Z.; Tan, H.; Lu, J.; Qin, P.; Xu, L. Identification of the Key Genes and MicroRNAs in Adult Acute Myeloid Leukemia with FLT3 Mutation by Bioinformatics Analysis. Int. J. Med. Sci. 2020, 17, 1269–1280. [Google Scholar] [CrossRef] [PubMed]
- Havelange, V.; Ranganathan, P.; Geyer, S.; Nicolet, D.; Huang, X.; Yu, X.; Volinia, S.; Kornblau, S.M.; Andreeff, M.; Croce, C.M.; et al. Implications of the MiR-10 Family in Chemotherapy Response of NPM1-Mutated AML. Blood 2014, 123, 2412–2415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhi, F.; Cao, X.; Xie, X.; Wang, B.; Dong, W.; Gu, W.; Ling, Y.; Wang, R.; Yang, Y.; Liu, Y. Identification of Circulating MicroRNAs as Potential Biomarkers for Detecting Acute Myeloid Leukemia. PLoS ONE 2013, 8, e56718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Min, J.; Sui, X.; Qian, Y.; Liu, Y.; Liu, Z.; Zhou, H.; Li, X.; Gong, Y. MicroRNA-26a-5p and MicroRNA-23b-3p up-Regulate Peroxiredoxin III in Acute Myeloid Leukemia. Leuk. Lymphoma 2015, 56, 460–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.; Hur, E.-H.; Moon, J.H.; Goo, B.-K.; Choi, D.R.; Lee, J.-H. Expression and Prognostic Significance of MicroRNAs in Korean Patients with Myelodysplastic Syndrome. Korean J. Intern. Med. 2019, 34, 390–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.-Y.; Lai, H.-F.; Huang, T.-C.; Chen, Y.-G.; Ye, R.-H.; Chang, P.-Y.; Lai, S.-W.; Chen, Y.-C.; Lee, C.-H.; Liu, W.-N.; et al. Aberrantly Reduced Expression of MiR-342-5p Contributes to CCND1-Associated Chronic Myeloid Leukemia Progression and Imatinib Resistance. Cell Death Dis. 2021, 12, 908. [Google Scholar] [CrossRef]
- Zhou, H.; Jia, X.; Yang, F.; Shi, P. MiR-148a-3p Suppresses the Progression of Acute Myeloid Leukemia via Targeting Cyclin-Dependent Kinase 6 (CDK6). Bioengineered 2021, 12, 4508–4519. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Wang, X.; Qi, P.; Liu, C.; Wang, S.; Wan, Q.; Liu, Y.; Su, Y.; Jin, L.; Liu, Y.; et al. Tumor Microenvironmental Competitive Endogenous RNA Network and Immune Cells Act as Robust Prognostic Predictor of Acute Myeloid Leukemia. Front. Oncol. 2021, 11, 584884. [Google Scholar] [CrossRef] [PubMed]
- Ozdogan, H.; Gur Dedeoglu, B.; Oztemur Islakoglu, Y.; Aydos, A.; Kose, S.; Atalay, A.; Yegin, Z.A.; Avcu, F.; Uckan Cetinkaya, D.; Ilhan, O. DICER1 Gene and MiRNA Dysregulation in Mesenchymal Stem Cells of Patients with Myelodysplastic Syndrome and Acute Myeloblastic Leukemia. Leuk. Res. 2017, 63, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, L.; Chen, L.; Liu, H.; Ren, S.; Tao, Y. Long Noncoding RNA DANCR Confers Cytarabine Resistance in Acute Myeloid Leukemia by Activating Autophagy via the MiR-874-3P/ATG16L1 Axis. Mol. Oncol. 2021, 15, 1203–1216. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Chen, S.; Lu, J.; Yuan, D.; He, L.; Qin, P.; Tan, H.; Xu, L. MicroRNA-363-3p Promote the Development of Acute Myeloid Leukemia with RUNX1 Mutation by Targeting SPRYD4 and FNDC3B. Medicine 2021, 100, e25807. [Google Scholar] [CrossRef] [PubMed]
- Drobna, M.; Szarzyńska, B.; Jaksik, R.; Sędek, Ł.; Kuchmiy, A.; Taghon, T.; Van Vlierberghe, P.; Szczepański, T.; Witt, M.; Dawidowska, M. Hsa-MiR-20b-5p and Hsa-MiR-363-3p Affect Expression of PTEN and BIM Tumor Suppressor Genes and Modulate Survival of T-ALL Cells In Vitro. Cells 2020, 9, 1137. [Google Scholar] [CrossRef]
- Sachs, K.; Sarver, A.L.; Noble-Orcutt, K.E.; LaRue, R.S.; Antony, M.L.; Chang, D.; Lee, Y.; Navis, C.M.; Hillesheim, A.L.; Nykaza, I.R.; et al. Single-Cell Gene Expression Analyses Reveal Distinct Self-Renewing and Proliferating Subsets in the Leukemia Stem Cell Compartment in Acute Myeloid Leukemia. Cancer Res. 2020, 80, 458–470. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Y.; Xing, P.; Liu, Q.; Zhang, C.; Sui, Y.; Wu, C. Development and Validation of a Hypoxia-Related Prognostic Signature for Breast Cancer. Oncol. Lett. 2020, 20, 1906–1914. [Google Scholar] [CrossRef] [PubMed]
- Silva, G.; Cardoso, B.A.; Belo, H.; Almeida, A.M. Vorinostat Induces Apoptosis and Differentiation in Myeloid Malignancies: Genetic and Molecular Mechanisms. PLoS ONE 2013, 8, e53766. [Google Scholar] [CrossRef]
- Subedi, A.; Liu, Q.; Ayyathan, D.M.; Sharon, D.; Cathelin, S.; Hosseini, M.; Xu, C.; Voisin, V.; Bader, G.D.; D’Alessandro, A.; et al. Nicotinamide Phosphoribosyltransferase Inhibitors Selectively Induce Apoptosis of AML Stem Cells by Disrupting Lipid Homeostasis. Cell Stem Cell 2021, 28, 1851–1867.e8. [Google Scholar] [CrossRef]
- Kersten, B.; Valkering, M.; Wouters, R.; van Amerongen, R.; Hanekamp, D.; Kwidama, Z.; Valk, P.; Ossenkoppele, G.; Zeijlemaker, W.; Kaspers, G.; et al. CD45RA, a specific marker for leukaemia stem cell sub-populations in acute myeloid leukaemia. Br. J. Haematol. 2016, 173, 219–235. [Google Scholar] [CrossRef]
- Zeijlemaker, W.; Kelder, A.; Wouters, R.; Valk, P.J.; Witte, B.I.; Cloos, J.; Ossenkoppele, G.J.; Schuurhuis, G.J. Absence of leukaemic CD34 cells in acute myeloid leukaemia is of high prognostic value: A longstanding controversy deciphered. Br. J. Haematol. 2015, 171, 227–238. [Google Scholar] [CrossRef] [Green Version]
- Zeijlemaker, W.; Kelder, A.; Oussoren-Brockhoff, Y.J.; Scholten, W.J.; Snel, A.N.; Veldhuizen, D.; Cloos, J.; Ossenkoppele, G.J.; Schuurhuis, G.J. A simple one-tube assay for immunophenotypical quantification of leukemic stem cells in acute myeloid leukemia. Leukemia 2016, 30, 439–446. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variation | Median (Range) | ||
|---|---|---|---|
| Age, years | 14 (10–15) | ||
| WBC count, ×109 | 79 (58.1–118) | ||
| Morphological blast count | |||
| BM% | 88 (34–95) | ||
| PB% | 74 (38–78) | ||
| N | % | ||
| Gender | |||
| F | 3 | 75.0% | |
| M | 1 | 25.0% | |
| Study protocol | |||
| DB AML-01 | 4 | 100.0% | |
| CNS involvement | 0 | 0.0% | |
| WT1 overexpression | 2 | 50.0% | |
| Fusion transcript | 2 | 50.0% | |
| CBF leukemia | 2 | 50.0% | |
| AML1::ETO + C-KITWT | 1 | 25.0% | |
| AML1::ETO + C-KITMUT | 0 | 0.0% | |
| CBF::MYH11 | 1 | 25.0% | |
| Gene mutation | |||
| NPM1 | 0 | 0.0% | |
| CEBPA | 0 | 0.0% | |
| FLT3 | |||
| ITD | 2 | 50.0% | |
| WT | 2 | 50.0% | |
| Karyotype | |||
| Normal | 1 | 25.0% | |
| Abnormal | 3 | 75.0% | |
| FAB classification | |||
| M0 | 1 | 25.0% | |
| M2 | 1 | 25.0% | |
| M4 | 2 | 50.0% | |
| Transcript ID | Location | Strand | Class |
|---|---|---|---|
| Lnc-CHST2-2:1 | chr3:143123362-143131893 | + | intronic |
| Lnc-EPS15L1-3:1 | chr19:16283359-16324514 | − | bidirectional |
| Lnc-KLHL25-4:1 | chr15:85621264-85627689 | − | antisense |
| Lnc-GSG1-1:1 | chr12:13196786-13197774 | − | antisense |
| Lnc-RGMA-1:1 | chr15:92805770-92808567 | − | intronic |
| Lnc-KMT2E-1:10 | chr7:104941063-104962060 | + | intergenic |
| LINC01220:10 | chr14:75294441-75296638 | + | intergenic |
| LINC00649:11 | chr21:33931554-33977774 | + | antisense |
| Lnc-LYST-4:1 | chr1:235826323-235883708 | − | intronic |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vanhooren, J.; Van Camp, L.; Depreter, B.; de Jong, M.; Uyttebroeck, A.; Van Damme, A.; Dedeken, L.; Dresse, M.-F.; van der Werff ten Bosch, J.; Hofmans, M.; et al. Deciphering the Non-Coding RNA Landscape of Pediatric Acute Myeloid Leukemia. Cancers 2022, 14, 2098. https://doi.org/10.3390/cancers14092098
Vanhooren J, Van Camp L, Depreter B, de Jong M, Uyttebroeck A, Van Damme A, Dedeken L, Dresse M-F, van der Werff ten Bosch J, Hofmans M, et al. Deciphering the Non-Coding RNA Landscape of Pediatric Acute Myeloid Leukemia. Cancers. 2022; 14(9):2098. https://doi.org/10.3390/cancers14092098
Chicago/Turabian StyleVanhooren, Jolien, Laurens Van Camp, Barbara Depreter, Martijn de Jong, Anne Uyttebroeck, An Van Damme, Laurence Dedeken, Marie-Françoise Dresse, Jutte van der Werff ten Bosch, Mattias Hofmans, and et al. 2022. "Deciphering the Non-Coding RNA Landscape of Pediatric Acute Myeloid Leukemia" Cancers 14, no. 9: 2098. https://doi.org/10.3390/cancers14092098
APA StyleVanhooren, J., Van Camp, L., Depreter, B., de Jong, M., Uyttebroeck, A., Van Damme, A., Dedeken, L., Dresse, M.-F., van der Werff ten Bosch, J., Hofmans, M., Philippé, J., De Moerloose, B., & Lammens, T. (2022). Deciphering the Non-Coding RNA Landscape of Pediatric Acute Myeloid Leukemia. Cancers, 14(9), 2098. https://doi.org/10.3390/cancers14092098