

Immune-Related Adverse Events Due to Cancer Immunotherapy: Immune Mechanisms and Clinical Manifestations
 , , ,
, , ,  , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Mechanisms and Targets of Immune Checkpoint Inhibitors
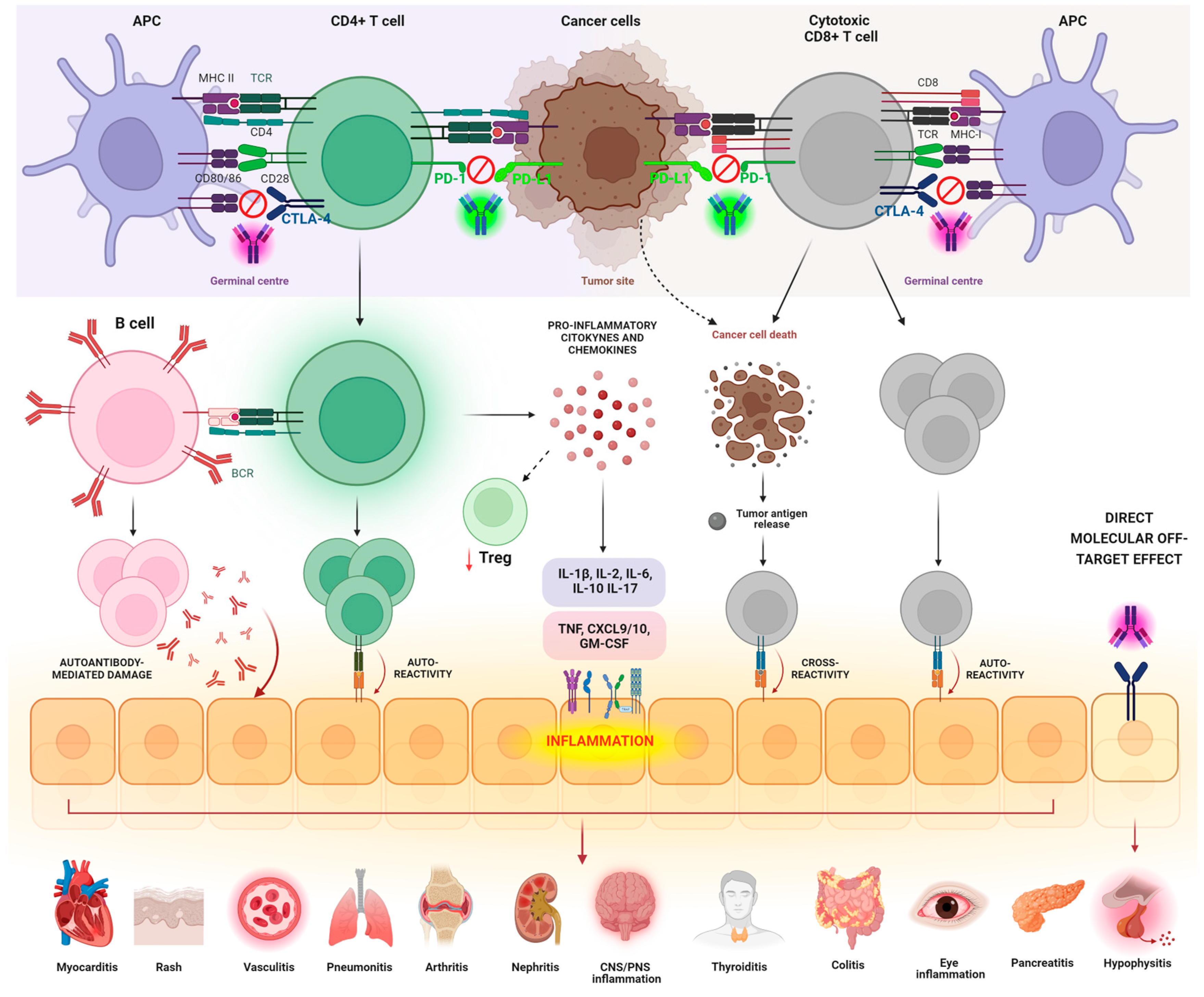
3. Pathogenic Mechanisms Underlying irAEs
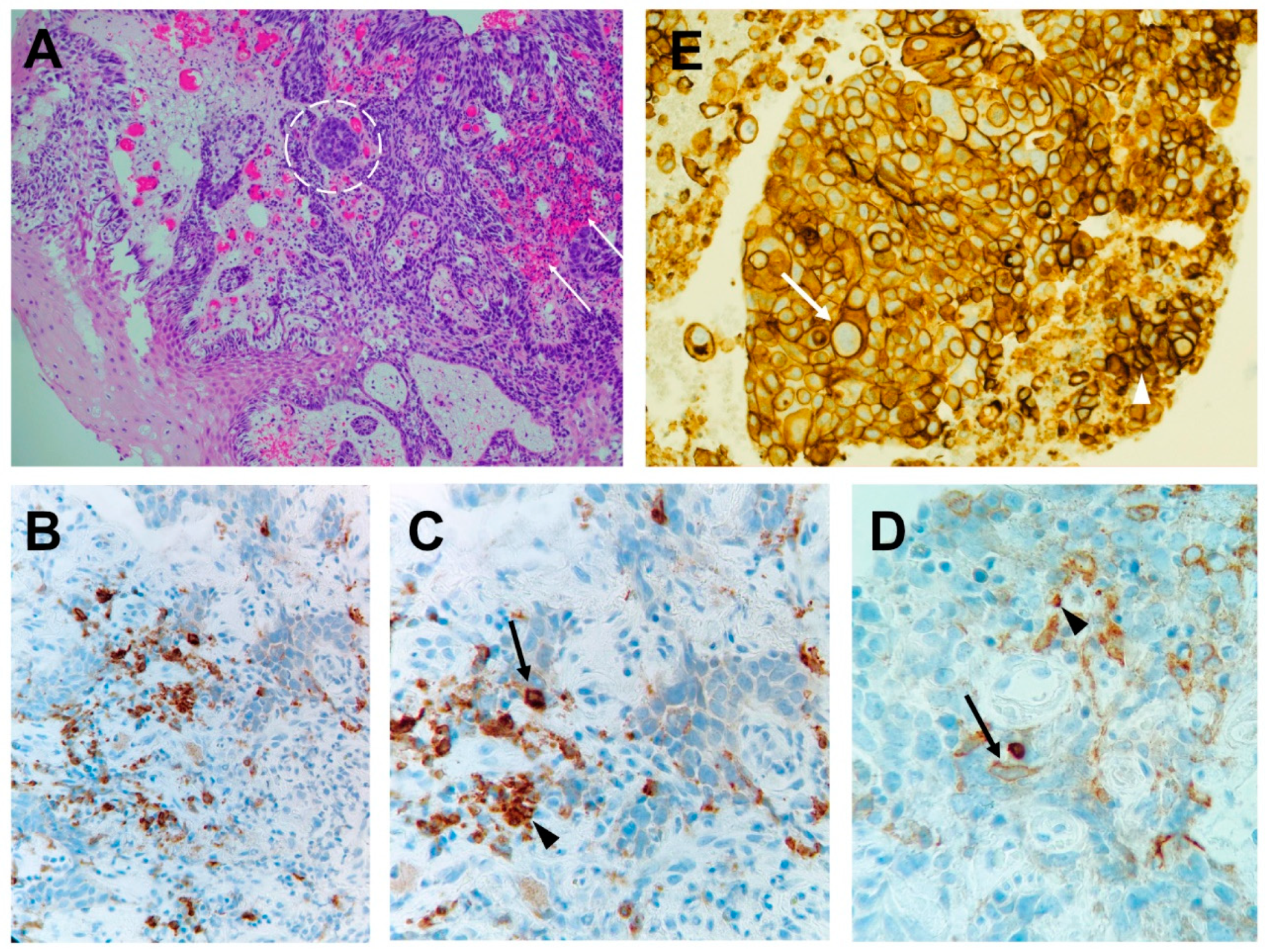
3.1. T-Cell-Mediated Mechanisms
3.1.1. Breach of Self-Tolerance of T Cells
3.1.2. Shared Antigens/Cross-Reactivity
3.1.3. Imbalance between Tregs and T-Effector Cells
3.2. B-Cell-Mediated Mechanisms
3.3. AutoAbs’ Action
3.4. Direct Effect of Monoclonal Abs
3.5. Inflammation and Cytokine-Mediated Mechanisms
4. Host-Specific and Environmental Factors Predisposed to irAEs
4.1. Role of the Host Microbiota
4.2. Genetic Background
4.3. Pre-Existing Autoimmunity
4.4. Patient-Specific Demographic Factors (Age, Sex, and Body Mass Index)
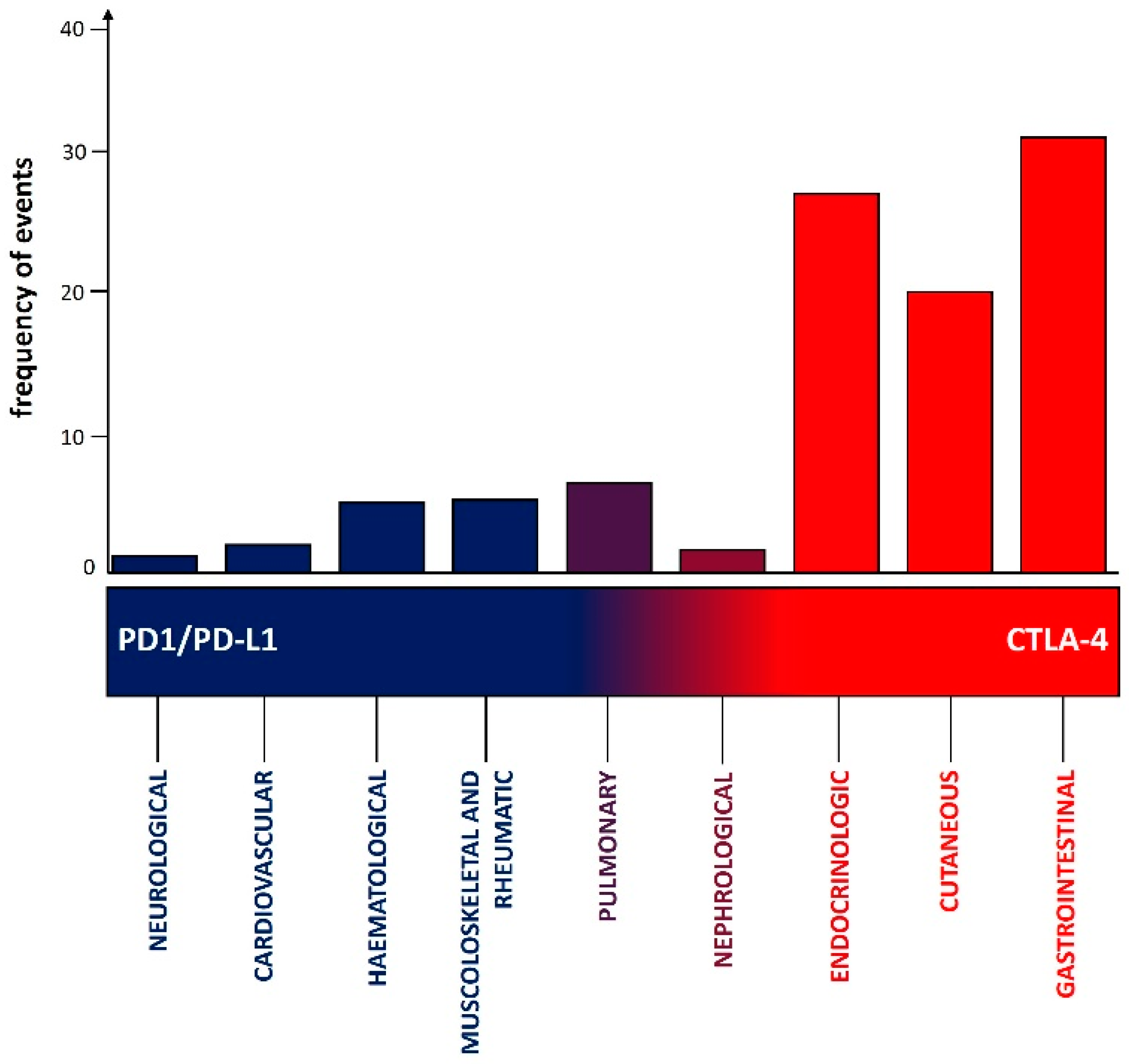
5. Main Clinical Manifestations Due to CTLA4 and PD1 Pathway Inhibitors
5.1. Gastrointestinal irAEs
5.2. Endocrinologic irAEs
5.3. Cutaneous irAEs
5.4. Pulmonary irAEs
5.5. Musculoskeletal irAEs
5.6. Hematological irAEs
5.7. Cardiovascular irAEs
5.8. Nephrological irAEs
5.9. Neurological irAEs
6. Current Limitations and Future Perspectives
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lebbé, C.; Weber, J.S.; Maio, M.; Neyns, B.; Harmankaya, K.; Hamid, O.; O’Day, S.J.; Konto, C.; Cykowski, L.; McHenry, M.B.; et al. Survival Follow-up and Ipilimumab Retreatment of Patients with Advanced Melanoma Who Received Ipilimumab in Prior Phase II Studies. Ann. Oncol. 2014, 25, 2277–2284. [Google Scholar] [CrossRef] [PubMed]
- Wolchok, J.D.; Chiarion-Sileni, V.; Gonzalez, R.; Rutkowski, P.; Grob, J.-J.; Cowey, C.L.; Lao, C.D.; Wagstaff, J.; Schadendorf, D.; Ferrucci, P.F.; et al. Overall Survival with Combined Nivolumab and Ipilimumab in Advanced Melanoma. N. Engl. J. Med. 2017, 377, 1345–1356. [Google Scholar] [CrossRef] [PubMed]
- Topalian, S.L.; Hodi, F.S.; Brahmer, J.R.; Gettinger, S.N.; Smith, D.C.; McDermott, D.F.; Powderly, J.D.; Sosman, J.A.; Atkins, M.B.; Leming, P.D.; et al. Five-Year Survival and Correlates among Patients with Advanced Melanoma, Renal Cell Carcinoma, or Non–Small Cell Lung Cancer Treated with Nivolumab. JAMA Oncol. 2019, 5, 1411–1420. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.C.; Duffy, C.R.; Allison, J.P. Fundamental Mechanisms of Immune Checkpoint Blockade Therapy. Cancer Discov. 2018, 8, 1069–1086. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Allison, J.P. Immune Checkpoint Targeting in Cancer Therapy: Toward Combination Strategies with Curative Potential. Cell 2015, 161, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Puzanov, I.; Diab, A.; Abdallah, K.; Bingham, C.O.; Brogdon, C.; Dadu, R.; Hamad, L.; Kim, S.; Lacouture, M.E.; LeBoeuf, N.R.; et al. Managing Toxicities Associated with Immune Checkpoint Inhibitors: Consensus Recommendations from the Society for Immunotherapy of Cancer (SITC) Toxicity Management Working Group. J. Immunother. Cancer 2017, 5, 95. [Google Scholar] [CrossRef] [PubMed]
- Haanen, J.; Obeid, M.; Spain, L.; Carbonnel, F.; Wang, Y.; Robert, C.; Lyon, A.R.; Wick, W.; Kostine, M.; Peters, S.; et al. Management of Toxicities from Immunotherapy: ESMO Clinical Practice Guideline for Diagnosis, Treatment and Follow-up. Ann. Oncol. 2022, 33, 1217–1238. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.A.; Schneider, B.J.; Brahmer, J.; Andrews, S.; Armand, P.; Bhatia, S.; Budde, L.E.; Costa, L.; Davies, M.; Dunnington, D.; et al. Management of Immunotherapy-Related Toxicities, Version 1.2019, NCCN Clinical Practice Guidelines in Oncology. J. Natl. Compr. Cancer Netw. 2019, 17, 255–289. [Google Scholar] [CrossRef]
- Ramos-Casals, M.; Brahmer, J.R.; Callahan, M.K.; Flores-Chávez, A.; Keegan, N.; Khamashta, M.A.; Lambotte, O.; Mariette, X.; Prat, A.; Suárez-Almazor, M.E. Immune-Related Adverse Events of Checkpoint Inhibitors. Nat. Rev. Dis. Primers 2020, 6, 38. [Google Scholar] [CrossRef]
- Ribas, A.; Wolchok, J.D. Cancer Immunotherapy Using Checkpoint Blockade. Science 2018, 359, 1350–1355. [Google Scholar] [CrossRef]
- Fritz, J.M.; Lenardo, M.J. Development of Immune Checkpoint Therapy for Cancer. J. Exp. Med. 2019, 216, 1244–1254. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Tagami, T.; Yamazaki, S.; Uede, T.; Shimizu, J.; Sakaguchi, N.; Mak, T.W.; Sakaguchi, S. Immunologic Self-Tolerance Maintained by Cd25+Cd4+Regulatory T Cells Constitutively Expressing Cytotoxic T Lymphocyte–Associated Antigen 4. J. Exp. Med. 2000, 192, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Willsmore, Z.N.; Coumbe, B.G.T.; Crescioli, S.; Reci, S.; Gupta, A.; Harris, R.J.; Chenoweth, A.; Chauhan, J.; Bax, H.J.; McCraw, A.; et al. Combined Anti-PD-1 and Anti-CTLA-4 Checkpoint Blockade: Treatment of Melanoma and Immune Mechanisms of Action. Eur. J. Immunol. 2021, 51, 544–556. [Google Scholar] [CrossRef] [PubMed]
- Sansom, D.M. CD28, CTLA-4 and Their Ligands: Who Does What and to Whom? Immunology 2000, 101, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, O.S.; Zheng, Y.; Nakamura, K.; Attridge, K.; Manzotti, C.; Schmidt, E.M.; Baker, J.; Jeffery, L.E.; Kaur, S.; Briggs, Z.; et al. Trans-Endocytosis of CD80 and CD86: A Molecular Basis for the Cell Extrinsic Function of CTLA-4. Science 2011, 332, 600. [Google Scholar] [CrossRef] [PubMed]
- Friedline, R.H.; Brown, D.S.; Nguyen, H.; Kornfeld, H.; Lee, J.; Zhang, Y.; Appleby, M.; Der, S.D.; Kang, J.; Chambers, C.A. CD4+ Regulatory T Cells Require CTLA-4 for the Maintenance of Systemic Tolerance. J. Exp. Med. 2009, 206, 421. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.S.K.; Sansom, D.M. Confusing Signals: Recent Progress in CTLA-4 Biology. Trends Immunol. 2015, 36, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Read, S.; Greenwald, R.; Izcue, A.; Robinson, N.; Mandelbrot, D.; Francisco, L.; Sharpe, A.H.; Powrie, F. Blockade of CTLA-4 on CD4+CD25+ Regulatory T Cells Abrogates Their Function In Vivo. J. Immunol. 2006, 177, 4376–4383. [Google Scholar] [CrossRef] [PubMed]
- Peggs, K.S.; Quezada, S.A.; Chambers, C.A.; Korman, A.J.; Allison, J.P. Blockade of CTLA-4 on Both Effector and Regulatory T Cell Compartments Contributes to the Antitumor Activity of Anti–CTLA-4 Antibodies. J. Exp. Med. 2009, 206, 1717–1725. [Google Scholar] [CrossRef]
- Wing, K.; Onishi, Y.; Prieto-Martin, P.; Yamaguchi, T.; Miyara, M.; Fehervari, Z.; Nomura, T.; Sakaguchi, S. CTLA-4 Control over Foxp3+ Regulatory T Cell Function. Science 2008, 322, 271–275. [Google Scholar] [CrossRef]
- Fife, B.T.; Bluestone, J.A. Control of Peripheral T-Cell Tolerance and Autoimmunity via the CTLA-4 and PD-1 Pathways. Immunol. Rev. 2008, 224, 166–182. [Google Scholar] [CrossRef]
- Francisco, L.M.; Sage, P.T.; Sharpe, A.H. The PD-1 Pathway in Tolerance and Autoimmunity. Immunol. Rev. 2010, 236, 219–242. [Google Scholar] [CrossRef] [PubMed]
- Fife, B.T.; Pauken, K.E. The Role of the PD-1 Pathway in Autoimmunity and Peripheral Tolerance. Ann. N. Y. Acad. Sci. 2011, 1217, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Pardoll, D.M. The Blockade of Immune Checkpoints in Cancer Immunotherapy. Nat. Rev. Cancer 2012, 12, 252–264. [Google Scholar] [CrossRef] [PubMed]
- Yearley, J.H.; Gibson, C.; Yu, N.; Moon, C.; Murphy, E.; Juco, J.; Lunceford, J.; Cheng, J.; Chow, L.Q.M.; Seiwert, T.Y.; et al. PD-L2 Expression in Human Tumors: Relevance to Anti-PD-1 Therapy in Cancer. Clin. Cancer Res. 2017, 23, 3158–3167. [Google Scholar] [CrossRef]
- Agata, Y.; Kawasaki, A.; Nishimura, H.; Ishida, Y.; Tsubata, T.; Yagita, H.; Honjo, T. Expression of the PD-1 Antigen on the Surface of Stimulated Mouse T and B Lymphocytes. Int. Immunol. 1996, 8, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Yokosuka, T.; Takamatsu, M.; Kobayashi-Imanishi, W.; Hashimoto-Tane, A.; Azuma, M.; Saito, T. Programmed Cell Death 1 Forms Negative Costimulatory Microclusters That Directly Inhibit T Cell Receptor Signaling by Recruiting Phosphatase SHP2. J. Exp. Med. 2012, 209, 1201–1217. [Google Scholar] [CrossRef] [PubMed]
- Keir, M.E.; Butte, M.J.; Freeman, G.J.; Sharpe, A.H. PD-1 and Its Ligands in Tolerance and Immunity. Annu. Rev. Immunol. 2008, 26, 677–704. [Google Scholar] [CrossRef] [PubMed]
- Doroshow, D.B.; Bhalla, S.; Beasley, M.B.; Sholl, L.M.; Kerr, K.M.; Gnjatic, S.; Wistuba, I.I.; Rimm, D.L.; Tsao, M.S.; Hirsch, F.R. PD-L1 as a Biomarker of Response to Immune-Checkpoint Inhibitors. Nat. Rev. Clin. Oncol. 2021, 18, 345–362. [Google Scholar] [CrossRef] [PubMed]
- Berti, A.; Bortolotti, R.; Dipasquale, M.; Kinspergher, S.; Prokop, L.; Grandi, G.; Inchiostro, S.; Paolazzi, G.; Caffo, O.; Veccia, A. Meta-Analysis of Immune-Related Adverse Events in Phase 3 Clinical Trials Assessing Immune Checkpoint Inhibitors for Lung Cancer. Crit. Rev. Oncol. Hematol. 2021, 162, 103351. [Google Scholar] [CrossRef]
- Wei, S.C.; Levine, J.H.; Cogdill, A.P.; Zhao, Y.; Anang, N.A.A.S.; Andrews, M.C.; Sharma, P.; Wang, J.; Wargo, J.A.; Pe’er, D.; et al. Distinct Cellular Mechanisms Underlie Anti-CTLA-4 and Anti-PD-1 Checkpoint Blockade. Cell 2017, 170, 1120–1133.e17. [Google Scholar] [CrossRef] [PubMed]
- Veccia, A.; Kostine, M.; Tison, A.; Dipasquale, M.; Kinspergher, S.; Prokop, L.; Grandi, G.; Inchiostro, S.; Caffo, O.; Paolazzi, G.; et al. Rheumatic Immune-and Nonimmune-Related Adverse Events in Phase 3 Clinical Trials Assessing PD-(L)1 Checkpoint Inhibitors for Lung Cancer: A Systematic Review and Meta-Analysis. Jt. Bone Spine 2022, 89, 105403. [Google Scholar] [CrossRef] [PubMed]
- Poto, R.; Troiani, T.; Criscuolo, G.; Marone, G.; Ciardiello, F.; Tocchetti, C.G.; Varricchi, G. Holistic Approach to Immune Checkpoint Inhibitor-Related Adverse Events. Front. Immunol. 2022, 13, 804597. [Google Scholar] [CrossRef]
- Esfahani, K.; Elkrief, A.; Calabrese, C.; Lapointe, R.; Hudson, M.; Routy, B.; Miller, W.H.; Calabrese, L. Moving towards Personalized Treatments of Immune-Related Adverse Events. Nat. Rev. Clin. Oncol. 2020, 17, 504–515. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, A.H.; Pauken, K.E. The Diverse Functions of the PD1 Inhibitory Pathway. Nat. Rev. Immunol. 2017, 18, 153–167. [Google Scholar] [CrossRef]
- Vétizou, M.; Pitt, J.M.; Daillère, R.; Lepage, P.; Waldschmitt, N.; Flament, C.; Rusakiewicz, S.; Routy, B.; Roberti, M.P.; Duong, C.P.M.; et al. Anticancer Immunotherapy by CTLA-4 Blockade Relies on the Gut Microbiota. Science 2015, 350, 1079–1084. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T.W. Role of the Microbiota in Immunity and Inflammation. Cell 2014, 157, 121. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wiesnoski, D.H.; Helmink, B.A.; Gopalakrishnan, V.; Choi, K.; DuPont, H.L.; Jiang, Z.D.; Abu-Sbeih, H.; Sanchez, C.A.; Chang, C.C.; et al. Fecal Microbiota Transplantation for Refractory Immune Checkpoint Inhibitor-Associated Colitis. Nat. Med. 2018, 24, 1804–1808. [Google Scholar] [CrossRef]
- Abdel-Wahab, N.; Diab, A.; Yu, R.K.; Futreal, A.; Criswell, L.A.; Tayar, J.H.; Dadu, R.; Shannon, V.; Shete, S.S.; Suarez-Almazor, M.E. Genetic Determinants of Immune-Related Adverse Events in Patients with Melanoma Receiving Immune Checkpoint Inhibitors. Cancer Immunol. Immunother. 2021, 70, 1939–1949. [Google Scholar] [CrossRef]
- Jiang, N.; Yu, Y.; Zhang, M.; Tang, Y.; Wu, D.; Wang, S.; Fang, Y.; Zhang, Y.; Meng, L.; Li, Y.; et al. Association between Germ-Line HLA and Immune-Related Adverse Events. Front. Immunol. 2022, 13, 952099. [Google Scholar] [CrossRef]
- Tivol, E.A.; Borriello, F.; Schweitzer, A.N.; Lynch, W.P.; Bluestone, J.A.; Sharpe, A.H. Loss of CTLA-4 Leads to Massive Lymphoproliferation and Fatal Multiorgan Tissue Destruction, Revealing a Critical Negative Regulatory Role of CTLA-4. Immunity 1995, 3, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, H.; Okazaki, T.; Tanaka, Y.; Nakatani, K.; Hara, M.; Matsumori, A.; Sasayama, S.; Mizoguchi, A.; Hiai, H.; Minato, N.; et al. Autoimmune Dilated Cardiomyopathy in PD-1 Receptor-Deficient Mice. Science 2001, 291, 319–322. [Google Scholar] [CrossRef]
- Lyubchenko, T.; Leung, D.Y.M.; Goleva, E. Why Are Immune Adverse Events so Common with Checkpoint Inhibitor Therapy? Ann. Allergy Asthma Immunol. 2021, 126, 608–610. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.Y.; Cham, J.; Zhang, L.; Fong, G.; Kwek, S.S.; Klinger, M.; Faham, M.; Fong, L. Immune Toxicities Elicted by CTLA-4 Blockade in Cancer Patients Are Associated with Early Diversification of the T-Cell Repertoire. Cancer Res. 2017, 77, 1322–1330. [Google Scholar] [CrossRef] [PubMed]
- Robert, L.; Tsoi, J.; Wang, X.; Emerson, R.; Homet, B.; Chodon, T.; Mok, S.; Huang, R.R.; Cochran, A.J.; Comin-Anduix, B.; et al. CTLA4 Blockade Broadens the Peripheral T-Cell Receptor Repertoire. Clin. Cancer Res. 2014, 20, 2424–2432. [Google Scholar] [CrossRef] [PubMed]
- Byrne, E.H.; Fisher, D.E. Immune and Molecular Correlates in Melanoma Treated with Immune Checkpoint Blockade. Cancer 2017, 123, 2143–2153. [Google Scholar] [CrossRef] [PubMed]
- König, D.; Läubli, H. Mechanisms of Immune-Related Complications in Cancer Patients Treated with Immune Checkpoint Inhibitors. Pharmacology 2021, 106, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.B.; Balko, J.M.; Compton, M.L.; Chalkias, S.; Gorham, J.; Xu, Y.; Hicks, M.; Puzanov, I.; Alexander, M.R.; Bloomer, T.L.; et al. Fulminant Myocarditis with Combination Immune Checkpoint Blockade. N. Engl. J. Med. 2016, 375, 1749–1755. [Google Scholar] [CrossRef] [PubMed]
- Flatz, L.; Berner, F.; Bomze, D.; Diem, S.; Ali, O.H.; Fässler, M.; Ring, S.; Niederer, R.; Ackermann, C.J.; Baumgaertner, P.; et al. Association of Checkpoint Inhibitor–Induced Toxic Effects with Shared Cancer and Tissue Antigens in Non–Small Cell Lung Cancer. JAMA Oncol. 2019, 5, 1043–1047. [Google Scholar] [CrossRef]
- Tanaka, A.; Sakaguchi, S. Regulatory T Cells in Cancer Immunotherapy. Cell Res. 2016, 27, 109–118. [Google Scholar] [CrossRef]
- Bennett, C.L.; Christie, J.; Ramsdell, F.; Brunkow, M.E.; Ferguson, P.J.; Whitesell, L.; Kelly, T.E.; Saulsbury, F.T.; Chance, P.F.; Ochs, H.D. The Immune Dysregulation, Polyendocrinopathy, Enteropathy, X-Linked Syndrome (IPEX) Is Caused by Mutations of FOXP3. Nat. Genet. 2001, 27, 20–21. [Google Scholar] [CrossRef] [PubMed]
- June, C.H.; Warshauer, J.T.; Bluestone, J.A. Is Autoimmunity the Achilles’ Heel of Cancer Immunotherapy? Nat. Med. 2017, 23, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Najafi, M.; Farhood, B.; Mortezaee, K. Contribution of Regulatory T Cells to Cancer: A Review. J. Cell Physiol. 2019, 234, 7983–7993. [Google Scholar] [CrossRef] [PubMed]
- Helmink, B.A.; Reddy, S.M.; Gao, J.; Zhang, S.; Basar, R.; Thakur, R.; Yizhak, K.; Sade-Feldman, M.; Blando, J.; Han, G.; et al. B Cells and Tertiary Lymphoid Structures Promote Immunotherapy Response. Nature 2020, 577, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Cabrita, R.; Lauss, M.; Sanna, A.; Donia, M.; Skaarup Larsen, M.; Mitra, S.; Johansson, I.; Phung, B.; Harbst, K.; Vallon-Christersson, J.; et al. Tertiary Lymphoid Structures Improve Immunotherapy and Survival in Melanoma. Nature 2020, 577, 561–565. [Google Scholar] [CrossRef]
- Sage, P.T.; Paterson, A.M.; Lovitch, S.B.; Sharpe, A.H. The Coinhibitory Receptor CTLA-4 Controls B Cell Responses by Modulating T Follicular Helper, T Follicular Regulatory, and T Regulatory Cells. Immunity 2014, 41, 1026–1039. [Google Scholar] [CrossRef] [PubMed]
- Thibult, M.L.; Mamessier, E.; Gertner-Dardenne, J.; Pastor, S.; Just-Landi, S.; Xerri, L.; Chetaille, B.; Olive, D. PD-1 Is a Novel Regulator of Human B-Cell Activation. Int. Immunol. 2013, 25, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Kuehn, H.S.; Ouyang, W.; Lo, B.; Deenick, E.K.; Niemela, J.E.; Avery, D.T.; Schickel, J.N.; Tran, D.Q.; Stoddard, J.; Zhang, Y.; et al. Immune Dysregulation in Human Subjects with Heterozygous Germline Mutations in CTLA4. Science 2014, 345, 1623–1627. [Google Scholar] [CrossRef]
- Das, R.; Bar, N.; Ferreira, M.; Newman, A.M.; Zhang, L.; Bailur, J.K.; Bacchiocchi, A.; Kluger, H.; Wei, W.; Halaban, R.; et al. Early B Cell Changes Predict Autoimmunity Following Combination Immune Checkpoint Blockade. J. Clin. Investig. 2018, 128, 715–720. [Google Scholar] [CrossRef]
- Nishimura, K.; Konishi, T.; Ochi, T.; Watanabe, R.; Noda, T.; Fukumoto, T.; Miura, N.; Miyauchi, Y.; Kikugawa, T.; Takenaka, K.; et al. CD21lo B Cells Could Be a Potential Predictor of Immune-Related Adverse Events in Renal Cell Carcinoma. J. Pers. Med. 2022, 12, 888. [Google Scholar] [CrossRef]
- Griss, J.; Bauer, W.; Wagner, C.; Simon, M.; Chen, M.; Grabmeier-Pfistershammer, K.; Maurer-Granofszky, M.; Roka, F.; Penz, T.; Bock, C.; et al. B Cells Sustain Inflammation and Predict Response to Immune Checkpoint Blockade in Human Melanoma. Nat. Commun. 2019, 10, 4186. [Google Scholar] [CrossRef] [PubMed]
- Willsmore, Z.N.; Harris, R.J.; Crescioli, S.; Hussein, K.; Kakkassery, H.; Thapa, D.; Cheung, A.; Chauhan, J.; Bax, H.J.; Chenoweth, A.; et al. B Cells in Patients with Melanoma: Implications for Treatment with Checkpoint Inhibitor Antibodies. Front. Immunol. 2021, 11, 622442. [Google Scholar] [CrossRef]
- Ludwig, R.J.; Vanhoorelbeke, K.; Leypoldt, F.; Kaya, Z.; Bieber, K.; McLachlan, S.M.; Komorowski, L.; Luo, J.; Cabral-Marques, O.; Hammers, C.M.; et al. Mechanisms of Autoantibody-Induced Pathology. Front. Immunol. 2017, 8, 260497. [Google Scholar] [CrossRef] [PubMed]
- Osorio, J.C.; Ni, A.; Chaft, J.E.; Pollina, R.; Kasler, M.K.; Stephens, D.; Rodriguez, C.; Cambridge, L.; Rizvi, H.; Wolchok, J.D.; et al. Antibody-Mediated Thyroid Dysfunction during T-Cell Checkpoint Blockade in Patients with Non-Small-Cell Lung Cancer. Ann. Oncol. 2017, 28, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Zaenker, P.; Gray, E.S.; Ziman, M.R. Autoantibody Production in Cancer—The Humoral Immune Response toward Autologous Antigens in Cancer Patients. Autoimmun. Rev. 2016, 15, 477–483. [Google Scholar] [CrossRef]
- Zaenker, P.; Lo, J.; Pearce, R.; Cantwell, P.; Cowell, L.; Lee, M.; Quirk, C.; Law, H.; Gray, E.; Ziman, M.; et al. A Diagnostic Autoantibody Signature for Primary Cutaneous Melanoma. Oncotarget 2018, 9, 30539–30551. [Google Scholar] [CrossRef][Green Version]
- Sexauer, D.; Gray, E.; Zaenker, P. Tumour-Associated Autoantibodies as Prognostic Cancer Biomarkers—A Review. Autoimmun. Rev. 2022, 21, 103041. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, G.C.; Gainor, J.F.; Altan, M.; Kravets, S.; Dahlberg, S.E.; Gedmintas, L.; Azimi, R.; Rizvi, H.; Riess, J.W.; Hellmann, M.D.; et al. Safety of Programmed Death-1 Pathway Inhibitors among Patients with Non-Small-Cell Lung Cancer and Preexisting Autoimmune Disorders. J. Clin. Oncol. 2018, 36, 1905–1912. [Google Scholar] [CrossRef]
- Abdel-Wahab, N.; Shah, M.; Lopez-Olivo, M.A.; Suarez-Almazor, M.E. Use of Immune Checkpoint Inhibitors in the Treatment of Patients with Cancer and Preexisting Autoimmune Disease: A Systematic Review. Ann. Intern. Med. 2018, 168, 121–130. [Google Scholar] [CrossRef]
- De Filette, J.; Jansen, Y.; Schreuer, M.; Everaert, H.; Velkeniers, B.; Neyns, B.; Bravenboer, B. Incidence of Thyroid-Related Adverse Events in Melanoma Patients Treated with Pembrolizumab. J. Clin. Endocrinol. Metab. 2016, 101, 4431–4439. [Google Scholar] [CrossRef]
- Mazarico, I.; Capel, I.; Giménez-Palop, O.; Albert, L.; Berges, I.; Luchtenberg, F.; García, Y.; Fernández-Morales, L.A.; De Pedro, V.J.; Caixàs, A.; et al. Low Frequency of Positive Antithyroid Antibodies Is Observed in Patients with Thyroid Dysfunction Related to Immune Check Point Inhibitors. J. Endocrinol. Investig. 2019, 42, 1443–1450. [Google Scholar] [CrossRef]
- Yamauchi, I.; Yasoda, A.; Matsumoto, S.; Sakamori, Y.; Kim, Y.H.; Nomura, M.; Otsuka, A.; Yamasaki, T.; Saito, R.; Kitamura, M.; et al. Incidence, Features, and Prognosis of Immune-Related Adverse Events Involving the Thyroid Gland Induced by Nivolumab. PLoS ONE 2019, 14, e0216954. [Google Scholar] [CrossRef]
- Luo, J.; Martucci, V.L.; Quandt, Z.; Groha, S.; Murray, M.H.; Lovly, C.M.; Rizvi, H.; Egger, J.V.; Plodkowski, A.J.; Abu-Akeel, M.; et al. Immunotherapy-Mediated Thyroid Dysfunction: Genetic Risk and Impact on Outcomes with PD-1 Blockade in Non–Small Cell Lung Cancer. Clin. Cancer Res. 2021, 27, 5131–5140. [Google Scholar] [CrossRef]
- Bluestone, J.A.; Anderson, M.; Herold, K.C.; Stamatouli, A.M.; Quandt, Z.; Perdigoto, A.L.; Clark, P.L.; Kluger, H.; Weiss, S.A.; Gettinger, S.; et al. Collateral Damage: Insulin-Dependent Diabetes Induced with Checkpoint Inhibitors. Diabetes 2018, 67, 1471–1480. [Google Scholar] [CrossRef]
- Johansen, A.; Christensen, S.J.; Scheie, D.; Højgaard, J.L.S.; Kondziella, D. Neuromuscular Adverse Events Associated with Anti-PD-1 Monoclonal Antibodies: Systematic Review. Neurology 2019, 92, 663–674. [Google Scholar] [CrossRef]
- Siegel, J.; Totonchy, M.; Damsky, W.; Berk-Krauss, J.; Castiglione, F.; Sznol, M.; Petrylak, D.P.; Fischbach, N.; Goldberg, S.B.; Decker, R.H.; et al. Bullous Disorders Associated with Anti–PD-1 and Anti–PD-L1 Therapy: A Retrospective Analysis Evaluating the Clinical and Histopathologic Features, Frequency, and Impact on Cancer Therapy. J. Am. Acad. Dermatol. 2018, 79, 1081–1088. [Google Scholar] [CrossRef]
- Cappelli, L.C.; Dorak, M.T.; Bettinotti, M.P.; Bingham, C.O.; Shah, A.A. Association of HLA-DRB1 Shared Epitope Alleles and Immune Checkpoint Inhibitor-Induced Inflammatory Arthritis. Rheumatology 2019, 58, 476–480. [Google Scholar] [CrossRef]
- Iwama, S.; De Remigis, A.; Callahan, M.K.; Slovin, S.F.; Wolchok, J.D.; Caturegli, P. Pituitary Expression of CTLA-4 Mediates Hypophysitis Secondary to Administration of CTLA-4 Blocking Antibody. Sci. Transl. Med. 2014, 6. [Google Scholar] [CrossRef]
- Tahir, S.A.; Gao, J.; Miura, Y.; Blando, J.; Tidwell, R.S.S.; Zhao, H.; Subudhi, S.K.; Tawbi, H.; Keung, E.; Wargo, J.; et al. Autoimmune Antibodies Correlate with Immune Checkpoint Therapy-Induced Toxicities. Proc. Natl. Acad. Sci. USA 2019, 116, 22246–22251. [Google Scholar] [CrossRef]
- Frasca, F.; Piticchio, T.; Le Moli, R.; Malaguarnera, R.; Campennì, A.; Cannavò, S.; Ruggeri, R.M. Recent insights into the pathogenesis of autoimmune hypophysitis. Expert Rev. Clin. Immunol. 2021, 17, 1175–1185. [Google Scholar] [CrossRef]
- Chatenoud, L.; Ferran, C.; Reuter, A.; Legendre, C.; Gevaert, Y.; Kreis, H.; Franchimont, P.; Bach, J.F. Systemic Reaction to the Anti–T-Cell Monoclonal Antibody OKT3 in Relation to Serum Levels of Tumor Necrosis Factor and Interferon-α. N. Engl. J. Med. 1989, 320, 1420–1421. [Google Scholar] [CrossRef]
- Slota, A.; Khan, R.; Rahman, A.; Warner, E.A. Cytokine Release Syndrome As a Rare Complication of Nivolumab: A Case Report. Blood 2019, 134, 5630. [Google Scholar] [CrossRef]
- Ceschi, A.; Noseda, R.; Palin, K.; Verhamme, K. Immune Checkpoint Inhibitor-Related Cytokine Release Syndrome: Analysis of WHO Global Pharmacovigilance Database. Front. Pharmacol. 2020, 11, 529822. [Google Scholar] [CrossRef]
- Tarhini, A.A.; Zahoor, H.; Lin, Y.; Malhotra, U.; Sander, C.; Butterfield, L.H.; Kirkwood, J.M. Baseline Circulating IL-17 Predicts Toxicity While TGF-Β1 and IL-10 Are Prognostic of Relapse in Ipilimumab Neoadjuvant Therapy of Melanoma. J. Immunother. Cancer 2015, 3, 39. [Google Scholar] [CrossRef]
- Phillips, G.S.; Wu, J.; Hellmann, M.D.; Postow, M.A.; Rizvi, N.A.; Freites-Martinez, A.; Chan, D.; Dusza, S.; Motzer, R.J.; Rosenberg, J.E.; et al. Treatment Outcomes of Immune-Related Cutaneous Adverse Events. J. Clin. Oncol. 2019, 37, 2746–2758. [Google Scholar] [CrossRef]
- Kurimoto, C.; Inaba, H.; Ariyasu, H.; Iwakura, H.; Ueda, Y.; Uraki, S.; Takeshima, K.; Furukawa, Y.; Morita, S.; Yamamoto, Y.; et al. Predictive and Sensitive Biomarkers for Thyroid Dysfunctions during Treatment with Immune-Checkpoint Inhibitors. Cancer Sci. 2020, 111, 1468–1477. [Google Scholar] [CrossRef]
- Lim, S.Y.; Lee, J.H.; Gide, T.N.; Menzies, A.M.; Guminski, A.; Carlino, M.S.; Breen, E.J.; Yang, J.Y.H.; Ghazanfar, S.; Kefford, R.F.; et al. Circulating Cytokines Predict Immune-Related Toxicity in Melanoma Patients Receiving Anti-PD-1–Based Immunotherapy. Clin. Cancer Res. 2019, 25, 1557–1563. [Google Scholar] [CrossRef]
- Khan, S.; Khan, S.A.; Luo, X.; Fattah, F.J.; Saltarski, J.; Gloria-McCutchen, Y.; Lu, R.; Xie, Y.; Li, Q.; Wakeland, E.; et al. Immune Dysregulation in Cancer Patients Developing Immune-Related Adverse Events. Br. J. Cancer 2018, 120, 63–68. [Google Scholar] [CrossRef]
- Khan, S.; Gerber, D.E. Autoimmunity, Checkpoint Inhibitor Therapy and Immune-Related Adverse Events: A Review. Semin. Cancer Biol. 2020, 64, 93–101. [Google Scholar] [CrossRef]
- Lee, D.J.; Lee, H.J.; Farmer, J.R.; Reynolds, K.L. Mechanisms Driving Immune-Related Adverse Events in Cancer Patients Treated with Immune Checkpoint Inhibitors. Curr. Cardiol. Rep. 2021, 23, 98. [Google Scholar] [CrossRef]
- Nishijima, S.; Suda, W.; Oshima, K.; Kim, S.W.; Hirose, Y.; Morita, H.; Hattori, M. The Gut Microbiome of Healthy Japanese and Its Microbial and Functional Uniqueness. DNA Res. 2016, 23, 125–133. [Google Scholar] [CrossRef]
- Hamada, K.; Isobe, J.; Hattori, K.; Hosonuma, M.; Baba, Y.; Murayama, M.; Narikawa, Y.; Toyoda, H.; Funayama, E.; Tajima, K.; et al. Turicibacter and Acidaminococcus Predict Immune-Related Adverse Events and Efficacy of Immune Checkpoint Inhibitor. Front. Immunol. 2023, 14, 1164724. [Google Scholar] [CrossRef]
- Lasagna, A.; Mascaro, F.; Figini, S.; Basile, S.; Gambini, G.; Klersy, C.; Lenti, M.V.; Di Sabatino, A.; Di Benedetto, A.; Calvi, M.; et al. Impact of Proton Pump Inhibitors on the Onset of Gastrointestinal Immune-Related Adverse Events during Immunotherapy. Cancer Med. 2023, 12, 19530–19536. [Google Scholar] [CrossRef]
- Hu, M.; Lin, X.; Sun, T.; Shao, X.; Huang, X.; Du, W.; Guo, M.; Zhu, X.; Zhou, Y.; Tong, T.; et al. Gut Microbiome for Predicting Immune Checkpoint Blockade-Associated Adverse Events. Genome Med. 2024, 16, 16. [Google Scholar] [CrossRef]
- Hoefsmit, E.P.; Rozeman, E.A.; Haanen, J.B.A.G.; Blank, C.U. Susceptible Loci Associated with Autoimmune Disease as Potential Biomarkers for Checkpoint Inhibitor-Induced Immune-Related Adverse Events. ESMO Open 2019, 4, e000472. [Google Scholar] [CrossRef]
- Akturk, H.K.; Kahramangil, D.; Sarwal, A.; Hoffecker, L.; Murad, M.H.; Michels, A.W. Immune Checkpoint Inhibitor-Induced Type 1 Diabetes: A Systematic Review and Meta-Analysis. Diabet. Med. 2019, 36, 1075–1081. [Google Scholar] [CrossRef]
- Hasan Ali, O.; Berner, F.; Bomze, D.; Fässler, M.; Diem, S.; Cozzio, A.; Jörger, M.; Früh, M.; Driessen, C.; Lenz, T.L.; et al. Human Leukocyte Antigen Variation Is Associated with Adverse Events of Checkpoint Inhibitors. Eur. J. Cancer 2019, 107, 8–14. [Google Scholar] [CrossRef]
- Johnson, D.B.; Sullivan, R.J.; Ott, P.A.; Carlino, M.S.; Khushalani, N.I.; Ye, F.; Guminski, A.; Puzanov, I.; Lawrence, D.P.; Buchbinder, E.I.; et al. Ipilimumab Therapy in Patients with Advanced Melanoma and Preexisting Autoimmune Disorders. JAMA Oncol. 2016, 2, 234–240. [Google Scholar] [CrossRef]
- Ibis, B.; Aliazis, K.; Cao, C.; Yenyuwadee, S.; Boussiotis, V.A. Immune-Related Adverse Effects of Checkpoint Immunotherapy and Implications for the Treatment of Patients with Cancer and Autoimmune Diseases. Front. Immunol. 2023, 14, 1197364. [Google Scholar] [CrossRef]
- Gulati, N.; Celen, A.; Johannet, P.; Mehnert, J.M.; Weber, J.; Krogsgaard, M.; Osman, I.; Zhong, J. Preexisting Immune-Mediated Inflammatory Disease Is Associated with Improved Survival and Increased Toxicity in Melanoma Patients Who Receive Immune Checkpoint Inhibitors. Cancer Med. 2021, 10, 7457–7465. [Google Scholar] [CrossRef]
- Lee, C.; Drobni, Z.D.; Zafar, A.; Gongora, C.A.; Zlotoff, D.A.; Alvi, R.M.; Taron, J.; Rambarat, P.K.; Schoenfeld, S.; Mosarla, R.C.; et al. Pre-Existing Autoimmune Disease Increases the Risk of Cardiovascular and Noncardiovascular Events after Immunotherapy. Cardio Oncol. 2022, 4, 660–669. [Google Scholar] [CrossRef]
- Wang, Z.; Aguilar, E.G.; Luna, J.I.; Dunai, C.; Khuat, L.T.; Le, C.T.; Mirsoian, A.; Minnar, C.M.; Stoffel, K.M.; Sturgill, I.R.; et al. Paradoxical Effects of Obesity on T Cell Function during Tumor Progression and PD-1 Checkpoint Blockade. Nat. Med. 2018, 25, 141–151. [Google Scholar] [CrossRef]
- Cortellini, A.; Bersanelli, M.; Buti, S.; Cannita, K.; Santini, D.; Perrone, F.; Giusti, R.; Tiseo, M.; Michiara, M.; Di Marino, P.; et al. A Multicenter Study of Body Mass Index in Cancer Patients Treated with Anti-PD-1/PD-L1 Immune Checkpoint Inhibitors: When Overweight Becomes Favorable. J. Immunother. Cancer 2019, 7, 57. [Google Scholar] [CrossRef]
- Naik, G.S.; Waikar, S.S.; Johnson, A.E.W.; Buchbinder, E.I.; Haq, R.; Hodi, F.S.; Schoenfeld, J.D.; Ott, P.A. Complex Inter-Relationship of Body Mass Index, Gender and Serum Creatinine on Survival: Exploring the Obesity Paradox in Melanoma Patients Treated with Checkpoint Inhibition. J. Immunother. Cancer 2019, 7, 89. [Google Scholar] [CrossRef]
- Guzman-Prado, Y.; Ben Shimol, J.; Samson, O. Body Mass Index and Immune-Related Adverse Events in Patients on Immune Checkpoint Inhibitor Therapies: A Systematic Review and Meta-Analysis. Cancer Immunol. Immunother. 2021, 70, 89–100. [Google Scholar] [CrossRef]
- Young, A.C.; Quach, H.T.; Song, H.; Davis, E.J.; Moslehi, J.J.; Ye, F.; Williams, G.R.; Johnson, D.B. Impact of Body Composition on Outcomes from Anti-PD1 +/− Anti-CTLA-4 Treatment in Melanoma. J. Immunother. Cancer 2020, 8, e000821. [Google Scholar] [CrossRef]
- Shah, K.P.; Song, H.; Ye, F.; Moslehi, J.J.; Balko, J.M.; Salem, J.E.; Johnson, D.B. Demographic Factors Associated with Toxicity in Patients Treated with Anti-Programmed Cell Death-1 Therapy. Cancer Immunol. Res. 2020, 8, 851–855. [Google Scholar] [CrossRef]
- Valpione, S.; Pasquali, S.; Campana, L.G.; Piccin, L.; Mocellin, S.; Pigozzo, J.; Chiarion-Sileni, V. Sex and Interleukin-6 Are Prognostic Factors for Autoimmune Toxicity Following Treatment with Anti-CTLA4 Blockade. J. Transl. Med. 2018, 16, 94. [Google Scholar] [CrossRef]
- Arnaud-Coffin, P.; Maillet, D.; Gan, H.K.; Stelmes, J.J.; You, B.; Dalle, S.; Péron, J. A Systematic Review of Adverse Events in Randomized Trials Assessing Immune Checkpoint Inhibitors. Int. J. Cancer 2019, 145, 639–648. [Google Scholar] [CrossRef]
- Balducci, D.; Quatraccioni, C.; Benedetti, A.; Marzioni, M.; Maroni, L. Gastrointestinal Disorders as Immune-Related Adverse Events. Explor. Target Antitumor Ther. 2021, 2, 174. [Google Scholar] [CrossRef]
- Wang, D.Y.; Ye, F.; Zhao, S.; Johnson, D.B. Incidence of Immune Checkpoint Inhibitor-Related Colitis in Solid Tumor Patients: A Systematic Review and Meta-Analysis. Oncoimmunology 2017, 6, e1344805. [Google Scholar] [CrossRef]
- Yanai, S.; Toya, Y.; Sugai, T.; Matsumoto, T. Gastrointestinal Adverse Events Induced by Immune-Checkpoint Inhibitors. Digestion 2021, 102, 965–973. [Google Scholar] [CrossRef]
- Nicolaides, S.; Boussioutas, A. Immune-Related Adverse Events of the Gastrointestinal System. Cancers 2023, 15, 691. [Google Scholar] [CrossRef]
- Barroso-Sousa, R.; Barry, W.T.; Garrido-Castro, A.C.; Hodi, F.S.; Min, L.; Krop, I.E.; Tolaney, S.M. Incidence of Endocrine Dysfunction Following the Use of Different Immune Checkpoint Inhibitor Regimens: A Systematic Review and Meta-Analysis. JAMA Oncol. 2018, 4, 173–182. [Google Scholar] [CrossRef]
- Cardona, Z.; Sosman, J.A.; Chandra, S.; Huang, W. Endocrine Side Effects of Immune Checkpoint Inhibitors. Front. Endocrinol. 2023, 14, 1157805. [Google Scholar] [CrossRef]
- Min, L.; Hodi, F.S.; Giobbie-Hurder, A.; Ott, P.A.; Luke, J.J.; Donahue, H.; Davis, M.; Carroll, R.S.; Kaiser, U.B. Systemic High-Dose Corticosteroid Treatment Does Not Improve the Outcome of Ipilimumab-Related Hypophysitis: A Retrospective Cohort Study. Clin. Cancer Res. 2015, 21, 749–755. [Google Scholar] [CrossRef]
- Ma, C.; Hodi, F.S.; Giobbie-Hurder, A.; Wang, X.; Zhou, J.; Zhang, A.; Zhou, Y.; Mao, F.; Angell, T.E.; Andrews, C.P.; et al. The Impact of High-Dose Glucocorticoids on the Outcome of Immune-Checkpoint Inhibitor–Related Thyroid Disorders. Cancer Immunol. Res. 2019, 7, 1214–1220. [Google Scholar] [CrossRef]
- Toi, Y.; Sugawara, S.; Sugisaka, J.; Ono, H.; Kawashima, Y.; Aiba, T.; Kawana, S.; Saito, R.; Aso, M.; Tsurumi, K.; et al. Profiling Preexisting Antibodies in Patients Treated with Anti–PD-1 Therapy for Advanced Non–Small Cell Lung Cancer. JAMA Oncol. 2019, 5, 376–383. [Google Scholar] [CrossRef]
- Okada, N.; Iwama, S.; Okuji, T.; Kobayashi, T.; Yasuda, Y.; Wada, E.; Onoue, T.; Goto, M.; Sugiyama, M.; Tsunekawa, T.; et al. Anti-Thyroid Antibodies and Thyroid Echo Pattern at Baseline as Risk Factors for Thyroid Dysfunction Induced by Anti-Programmed Cell Death-1 Antibodies: A Prospective Study. Br. J. Cancer 2020, 122, 771–777. [Google Scholar] [CrossRef]
- Al Ashi, S.I.; Thapa, B.; Flores, M.; Ahmed, R.; Rahim, S.E.G.; Amir, M.; Alomari, M.; Chadalavada, P.; Morrison, S.L.; Bena, J.F.; et al. Endocrine Toxicity and Outcomes in Patients with Metastatic Malignancies Treated with Immune Checkpoint Inhibitors. J. Endocr. Soc. 2021, 5, bvab100. [Google Scholar] [CrossRef]
- Jessel, S.; Weiss, S.A.; Austin, M.; Mahajan, A.; Etts, K.; Zhang, L.; Aizenbud, L.; Perdigoto, A.L.; Hurwitz, M.; Sznol, M.; et al. Immune Checkpoint Inhibitor-Induced Hypophysitis and Patterns of Loss of Pituitary Function. Front. Oncol. 2022, 12, 836859. [Google Scholar] [CrossRef]
- Amereller, F.; Deutschbein, T.; Joshi, M.; Schopohl, J.; Schilbach, K.; Detomas, M.; Duffy, L.; Carroll, P.; Papa, S.; Störmann, S. Differences between Immunotherapy-Induced and Primary Hypophysitis—A Multicenter Retrospective Study. Pituitary 2022, 25, 152–158. [Google Scholar] [CrossRef]
- Lu, D.; Yao, J.; Yuan, G.; Gao, Y.; Zhang, J.; Guo, X. Immune Checkpoint Inhibitor-Associated New-Onset Primary Adrenal Insufficiency: A Retrospective Analysis Using the FAERS. J. Endocrinol. Investig. 2022, 45, 2131–2137. [Google Scholar] [CrossRef]
- Quandt, Z.; Young, A.; Anderson, M. Immune Checkpoint Inhibitor Diabetes Mellitus: A Novel Form of Autoimmune Diabetes. Clin. Exp. Immunol. 2020, 200, 131–140. [Google Scholar] [CrossRef]
- Collins, L.K.; Chapman, M.S.; Carter, J.B.; Samie, F.H. Cutaneous Adverse Effects of the Immune Checkpoint Inhibitors. Curr. Probl. Cancer 2017, 41, 125–128. [Google Scholar] [CrossRef]
- Sibaud, V. Dermatologic Reactions to Immune Checkpoint Inhibitors. Am. J. Clin. Dermatol. 2017, 19, 345–361. [Google Scholar] [CrossRef]
- Quach, H.T.; Johnson, D.B.; LeBoeuf, N.R.; Zwerner, J.P.; Dewan, A.K. Cutaneous Adverse Events Caused by Immune Checkpoint Inhibitors. J. Am. Acad. Dermatol. 2021, 85, 956–966. [Google Scholar] [CrossRef]
- Watanabe, T.; Yamaguchi, Y. Cutaneous Manifestations Associated with Immune Checkpoint Inhibitors. Front. Immunol. 2023, 14, 1071983. [Google Scholar] [CrossRef]
- Nishino, M.; Giobbie-Hurder, A.; Hatabu, H.; Ramaiya, N.H.; Hodi, F.S. Incidence of Programmed Cell Death 1 Inhibitor–Related Pneumonitis in Patients with Advanced Cancer: A Systematic Review and Meta-Analysis. JAMA Oncol. 2016, 2, 1607–1616. [Google Scholar] [CrossRef]
- Georgakopoulou, V.E.; Garmpis, N.; Mermigkis, D.; Damaskos, C.; Chlapoutakis, S.; Mantzouranis, K.; Gkoufa, A.; Papageorgiou, C.V.; Garmpi, A.; Makrodimitri, S.; et al. Pulmonary Adverse Events Due to Immune Checkpoint Inhibitors: A Literature Review. Monaldi Arch. Chest Dis. 2021, 92. [Google Scholar] [CrossRef]
- Kostine, M.; Finckh, A.; Bingham, C.O.; Visser, K.; Leipe, J.; Schulze-Koops, H.; Choy, E.H.; Benesova, K.; Radstake, T.R.D.J.; Cope, A.P.; et al. EULAR Points to Consider for the Diagnosis and Management of Rheumatic Immune-Related Adverse Events Due to Cancer Immunotherapy with Checkpoint Inhibitors. Ann. Rheum. Dis. 2021, 80, 36–48. [Google Scholar] [CrossRef]
- Le Burel, S.; Champiat, S.; Mateus, C.; Marabelle, A.; Michot, J.M.; Robert, C.; Belkhir, R.; Soria, J.C.; Laghouati, S.; Voisin, A.L.; et al. Prevalence of Immune-Related Systemic Adverse Events in Patients Treated with Anti-Programmed Cell Death 1/Anti-Programmed Cell Death-Ligand 1 Agents: A Single-Centre Pharmacovigilance Database Analysis. Eur. J. Cancer 2017, 82, 34–44. [Google Scholar] [CrossRef]
- Kostine, M.; Rouxel, L.; Barnetche, T.; Veillon, R.; Martin, F.; Dutriaux, C.; Dousset, L.; Pham-Ledard, A.; Prey, S.; Beylot-Barry, M.; et al. Rheumatic Disorders Associated with Immune Checkpoint Inhibitors in Patients with Cancer—Clinical Aspects and Relationship with Tumour Response: A Single-Centre Prospective Cohort Study. Ann. Rheum. Dis. 2018, 77, 393–398. [Google Scholar] [CrossRef]
- Belkhir, R.; Le Burel, S.; Dunogeant, L.; Marabelle, A.; Hollebecque, A.; Besse, B.; Leary, A.; Voisin, A.L.; Pontoizeau, C.; Coutte, L.; et al. Rheumatoid Arthritis and Polymyalgia Rheumatica Occurring after Immune Checkpoint Inhibitor Treatment. Ann. Rheum. Dis. 2017, 76, 1747–1750. [Google Scholar] [CrossRef]
- Calabrese, C.; Cappelli, L.C.; Kostine, M.; Kirchner, E.; Braaten, T.; Calabrese, L. Polymyalgia Rheumatica-like Syndrome from Checkpoint Inhibitor Therapy: Case Series and Systematic Review of the Literature. RMD Open 2019, 5, e000906. [Google Scholar] [CrossRef]
- Ramos-Casals, M.; Maria, A.; Suárez-Almazor, M.E.; Lambotte, O.; Fisher, B.A.; Hernández-Molina, G.; Guilpain, P.; Pundole, X.; Flores-Chávez, A.; Baldini, C.; et al. Sicca/Sjögren’s Syndrome Triggered by PD-1/PD-L1 Checkpoint Inhibitors. Data from the International Immunocancer Registry (ICIR). Clin. Exp. Rheumatol. 2019, 37, S114–S122. [Google Scholar]
- Tison, A.; Garaud, S.; Chiche, L.; Cornec, D.; Kostine, M. Immune-Checkpoint Inhibitor Use in Patients with Cancer and Pre-Existing Autoimmune Diseases. Nat. Rev. Rheumatol. 2022, 18, 641–656. [Google Scholar] [CrossRef]
- Tison, A.; Quéré, G.; Misery, L.; Funck-Brentano, E.; Danlos, F.X.; Routier, E.; Robert, C.; Loriot, Y.; Lambotte, O.; Bonniaud, B.; et al. Safety and Efficacy of Immune Checkpoint Inhibitors in Patients with Cancer and Preexisting Autoimmune Disease: A Nationwide, Multicenter Cohort Study. Arthritis Rheumatol. 2019, 71, 2100–2111. [Google Scholar] [CrossRef]
- Michot, J.M.; Lazarovici, J.; Tieu, A.; Champiat, S.; Voisin, A.L.; Ebbo, M.; Godeau, B.; Michel, M.; Ribrag, V.; Lambotte, O. Haematological Immune-Related Adverse Events with Immune Checkpoint Inhibitors, How to Manage? Eur. J. Cancer 2019, 122, 72–90. [Google Scholar] [CrossRef] [PubMed]
- Delanoy, N.; Michot, J.M.; Comont, T.; Kramkimel, N.; Lazarovici, J.; Dupont, R.; Champiat, S.; Chahine, C.; Robert, C.; Herbaux, C.; et al. Haematological Immune-Related Adverse Events Induced by Anti-PD-1 or Anti-PD-L1 Immunotherapy: A Descriptive Observational Study. Lancet Haematol. 2019, 6, e48–e57. [Google Scholar] [CrossRef] [PubMed]
- Omar, N.E.; El-Fass, K.A.; Abushouk, A.I.; Elbaghdady, N.; Barakat, A.E.M.; Noreldin, A.E.; Johar, D.; Yassin, M.; Hamad, A.; Elazzazy, S.; et al. Diagnosis and Management of Hematological Adverse Events Induced by Immune Checkpoint Inhibitors: A Systematic Review. Front. Immunol. 2020, 11, 531929. [Google Scholar] [CrossRef]
- Davis, E.J.; Salem, J.; Young, A.; Green, J.R.; Ferrell, P.B.; Ancell, K.K.; Lebrun-Vignes, B.; Moslehi, J.J.; Johnson, D.B. Hematologic Complications of Immune Checkpoint Inhibitors. Oncologist 2019, 24, 584–588. [Google Scholar] [CrossRef]
- Saliba, A.N.; Xie, Z.; Higgins, A.S.; Andrade-Gonzalez, X.A.; Fuentes-Bayne, H.E.; Hampel, P.J.; Kankeu Fonkoua, L.A.; Childs, D.S.; Rakshit, S.; Bezerra, E.D.; et al. Immune-Related Hematologic Adverse Events in the Context of Immune Checkpoint Inhibitor Therapy. Am. J. Hematol. 2021, 96, E362–E367. [Google Scholar] [CrossRef]
- Calvo, R. Hematological Side Effects of Immune Checkpoint Inhibitors: The Example of Immune-Related Thrombocytopenia. Front. Pharmacol. 2019, 10, 438076. [Google Scholar] [CrossRef]
- Diaz, L.; Jauzelon, B.; Dillies, A.C.; Le Souder, C.; Faillie, J.L.; Maria, A.T.J.; Palassin, P. Hemophagocytic Lymphohistiocytosis Associated with Immunological Checkpoint Inhibitors: A Pharmacovigilance Study. J. Clin. Med. 2023, 12, 1985. [Google Scholar] [CrossRef]
- Lyon, A.R.; Yousaf, N.; Battisti, N.M.L.; Moslehi, J.; Larkin, J. Immune Checkpoint Inhibitors and Cardiovascular Toxicity. Lancet Oncol. 2018, 19, e447–e458. [Google Scholar] [CrossRef]
- Raschi, E.; Rossi, S.; De Giglio, A.; Fusaroli, M.; Burgazzi, F.; Rinaldi, R.; Potena, L. Cardiovascular Toxicity of Immune Checkpoint Inhibitors: A Guide for Clinicians. Drug Saf. 2023, 46, 819–833. [Google Scholar] [CrossRef]
- Cozma, A.; Sporis, N.D.; Lazar, A.L.; Buruiana, A.; Ganea, A.M.; Malinescu, T.V.; Berechet, B.M.; Fodor, A.; Sitar-Taut, A.V.; Vlad, V.C.; et al. Cardiac Toxicity Associated with Immune Checkpoint Inhibitors: A Systematic Review. Int. J. Mol. Sci. 2022, 23, 10948. [Google Scholar] [CrossRef]
- Patel, R.P.; Parikh, R.; Gunturu, K.S.; Tariq, R.Z.; Dani, S.S.; Ganatra, S.; Nohria, A. Cardiotoxicity of Immune Checkpoint Inhibitors. Curr. Oncol. Rep. 2021, 23, 79. [Google Scholar] [CrossRef]
- Longhitano, E.; Muscolino, P.; Lo Re, C.; Ferrara, S.A.; Cernaro, V.; Gembillo, G.; Tessitore, D.; Speranza, D.; Figura, F.; Santarpia, M.; et al. Immune Checkpoint Inhibitors and the Kidney: A Focus on Diagnosis and Management for Personalised Medicine. Cancers 2023, 15, 1891. [Google Scholar] [CrossRef]
- Linee Guida Gestione Della Tossicita’ Da Immunoterapia|AIOM. Available online: https://www.aiom.it/linee-guida-aiom-2021-gestione-della-tossicita-da-immunoterapia (accessed on 2 March 2024).
- Gupta, S.; Cortazar, F.B.; Riella, L.V.; Leaf, D.E. Immune Checkpoint Inhibitor Nephrotoxicity: Update 2020. Kidney360 2020, 1, 130–140. [Google Scholar] [CrossRef]
- Miao, J.; Sise, M.E.; Herrmann, S.M. Immune Checkpoint Inhibitor Related Nephrotoxicity: Advances in Clinicopathologic Features, Noninvasive Approaches, and Therapeutic Strategy and Rechallenge. Front. Nephrol. 2022, 2, 1017921. [Google Scholar] [CrossRef]
- Cuzzubbo, S.; Javeri, F.; Tissier, M.; Roumi, A.; Barlog, C.; Doridam, J.; Lebbe, C.; Belin, C.; Ursu, R.; Carpentier, A.F. Neurological Adverse Events Associated with Immune Checkpoint Inhibitors: Review of the Literature. Eur. J. Cancer 2017, 73, 1–8. [Google Scholar] [CrossRef]
- Bruna, J.; Argyriou, A.A.; Anastopoulou, G.G.; Alemany, M.; Nadal, E.; Kalofonou, F.; Piulats, J.M.; Simó, M.; Velasco, R.; Kalofonos, H.P. Incidence and Characteristics of Neurotoxicity in Immune Checkpoint Inhibitors with Focus on Neuromuscular Events: Experience beyond the Clinical Trials. J. Peripher. Nerv. Syst. 2020, 25, 171–177. [Google Scholar] [CrossRef]
- Dubey, D.; David, W.S.; Reynolds, K.L.; Chute, D.F.; Clement, N.F.; Cohen, J.V.; Lawrence, D.P.; Mooradian, M.J.; Sullivan, R.J.; Guidon, A.C. Severe Neurological Toxicity of Immune Checkpoint Inhibitors: Growing Spectrum. Ann. Neurol. 2020, 87, 659–669. [Google Scholar] [CrossRef]
- Johnson, D.B.; Manouchehri, A.; Haugh, A.M.; Quach, H.T.; Balko, J.M.; Lebrun-Vignes, B.; Mammen, A.; Moslehi, J.J.; Salem, J.E. Neurologic Toxicity Associated with Immune Checkpoint Inhibitors: A Pharmacovigilance Study. J. Immunother. Cancer 2019, 7, 134. [Google Scholar] [CrossRef]
- Farina, A.; Birzu, C.; Elsensohn, M.-H.; Picca, A.; Muñiz-Castrillo, S.; Vogrig, A.; Villagrán-García, M.; Lundahl Ciano-Petersen, N.; Massacesi, L.; Hervier, B.; et al. Neurological Outcomes in Immune Checkpoint Inhibitor-Related Neurotoxicity. Brain Commun. 2023, 5, 20. [Google Scholar] [CrossRef] [PubMed]
- Marini, A.; Bernardini, A.; Gigli, G.L.; Valente, M.; Muñiz-Castrillo, S.; Honnorat, J.; Vogrig, A. Neurologic Adverse Events of Immune Checkpoint Inhibitors: A Systematic Review. Neurology 2021, 96, 754–766. [Google Scholar] [CrossRef]
- Vogrig, A.; Muñiz-Castrillo, S.; Desestret, V.; Joubert, B.; Honnorat, J. Pathophysiology of Paraneoplastic and Autoimmune Encephalitis: Genes, Infections, and Checkpoint Inhibitors. Ther. Adv. Neurol. Disord. 2020, 13, 1756286420932797. [Google Scholar] [CrossRef]
- Graus, F.; Dalmau, J. Paraneoplastic Neurological Syndromes in the Era of Immune-Checkpoint Inhibitors. Nat. Rev. Clin. Oncol. 2019, 16, 535–548. [Google Scholar] [CrossRef]
- Farina, A.; Villagrán-García, M.; Vogrig, A.; Zekeridou, A.; Muñiz-Castrillo, S.; Velasco, R.; Guidon, A.C.; Joubert, B.; Honnorat, J. Neurological Adverse Events of Immune Checkpoint Inhibitors and the Development of Paraneoplastic Neurological Syndromes. Lancet Neurol. 2024, 23, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Touat, M.; Maisonobe, T.; Knauss, S.; Ben Hadj Salem, O.; Hervier, B.; Auré, K.; Szwebel, T.A.; Kramkimel, N.; Lethrosne, C.; Bruch, J.F.; et al. Immune Checkpoint Inhibitor-Related Myositis and Myocarditis in Patients with Cancer. Neurology 2018, 91, e985–e994. [Google Scholar] [CrossRef] [PubMed]
- Shelly, S.; Triplett, J.D.; Pinto, M.V.; Milone, M.; Diehn, F.E.; Zekeridou, A.; Liewluck, T. Immune Checkpoint Inhibitor-Associated Myopathy: A Clinicoseropathologically Distinct Myopathy. Brain Commun. 2020, 2, fcaa181. [Google Scholar] [CrossRef] [PubMed]
- Aldrich, J.; Pundole, X.; Tummala, S.; Palaskas, N.; Andersen, C.R.; Shoukier, M.; Abdel-Wahab, N.; Deswal, A.; Suarez-Almazor, M.E. Inflammatory Myositis in Cancer Patients Receiving Immune Checkpoint Inhibitors. Arthritis Rheumatol. 2021, 73, 866–874. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Ishikawa, N.; Konoeda, F.; Seki, N.; Fukushima, S.; Takahashi, K.; Uhara, H.; Hasegawa, Y.; Inomata, S.; Otani, Y.; et al. Nivolumab-Related Myasthenia Gravis with Myositis and Myocarditis in Japan. Neurology 2017, 89, 1127–1134. [Google Scholar] [CrossRef] [PubMed]
- Marco, C.; Simó, M.; Alemany, M.; Casasnovas, C.; Domínguez, R.; Vilariño, N.; Calvo, M.; Martín-Liberal, J.; Brenes, J.; Sabater-Riera, J.; et al. Myasthenia Gravis Induced by Immune Checkpoint Inhibitors: An Emerging Neurotoxicity in Neuro-Oncology Practice: Case Series. J. Clin. Med. 2022, 12, 130. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zeng, H.; Fang, W.; Song, L. Clinical characteristics, treatment and outcome of nivolumab-induced myasthenia gravis. Invest New Drugs 2023, 41, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Dubey, D.; David, W.S.; Amato, A.A.; Reynolds, K.L.; Clement, N.F.; Chute, D.F.; Cohen, J.V.; Lawrence, D.P.; Mooradian, M.J.; Sullivan, R.J.; et al. Varied Phenotypes and Management of Immune Checkpoint Inhibitor-Associated Neuropathies. Neurology 2019, 93, e1093–e1103. [Google Scholar] [CrossRef]
- Vogrig, A.; Muñiz-Castrillo, S.; Joubert, B.; Picard, G.; Rogemond, V.; Skowron, F.; Egri, M.; Desestret, V.; Tilikete, C.; Psimaras, D.; et al. Cranial Nerve Disorders Associated with Immune Checkpoint Inhibitors. Neurology 2021, 96, E866–E875. [Google Scholar] [CrossRef]
- Valencia-Sanchez, C.; Sechi, E.; Dubey, D.; Flanagan, E.P.; McKeon, A.; Pittock, S.J.; Zekeridou, A. Immune Checkpoint Inhibitor-Associated Central Nervous System Autoimmunity. Eur. J. Neurol. 2023, 30, 2418–2429. [Google Scholar] [CrossRef]
- Vogrig, A.; Muñiz-Castrillo, S.; Joubert, B.; Picard, G.; Rogemond, V.; Marchal, C.; Chiappa, A.M.; Chanson, E.; Skowron, F.; Leblanc, A.; et al. Central Nervous System Complications Associated with Immune Checkpoint Inhibitors. J. Neurol. Neurosurg. Psychiatry 2020, 91, 772–778. [Google Scholar] [CrossRef]
- Vogrig, A.; Fouret, M.; Joubert, B.; Picard, G.; Rogemond, V.; Pinto, A.L.; Muñiz-Castrillo, S.; Roger, M.; Raimbourg, J.; Dayen, C.; et al. Increased Frequency of Anti-Ma2 Encephalitis Associated with Immune Checkpoint Inhibitors. Neurol. Neuroimmunol. Neuroinflamm. 2019, 6, 604. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Checkpoint Targets | Drugs | Indications |
|---|---|---|
| CTLA-4 | Ipilimumab 1, 2, 3, 5 | NSCLC, advanced CRC, advanced MM, HL |
| Tremelimumab 1, 2 | NSCLC, HCC | |
| PD-1 | Nivolumab 1, 2, 3, 4, 5 | HNSCC, NSCLC, GC, ESGC, CRC, HCC, RCC, HL, MM, skin cancer |
| Pembrolizumab 1, 2, 3, 4, 5 | HNSCC, NSCLC, GC, ESGC, CRC, HCC, RCC, HL, MM, skin cancer, TNBC | |
| Cemiplimab 1, 2, 3 | skin cancer, NSCLC | |
| Sintilimab 1, 2, 5 | NSCLC, HCC, HL | |
| Camrelizumab 2, 5 | NSCLC, HCC, HL, ESCC, HNSCC | |
| Dostarlimab 1, 2, 3 | UCEC | |
| Tislelizumab 1, 5 | NSCLC, HL, BRCA | |
| Penpulimab 5 | HCC, GC, NSCLC, NPC, HL | |
| Toripalimab 1, 5 | Skin cancer, HNSCC, BRCA | |
| Zimberelimab 5 | HL | |
| Serplulimab 5 | GC, CRC, NSCLC | |
| Pucotenlimab 5 | CRC, MM | |
| PD-L1 | Durvalumab 1, 2, 3, 5 | NSCLC, SCLC, BRCA |
| Atezolizumab 1, 2, 3, 5 | NSCLC, SCLC, BRCA, skin cancer, HCC, TNBC | |
| Avelumab 1, 2, 3 | skin cancer, RCC, BRCA | |
| Envafolimab 5 | CRC | |
| Sugevalimab 5 | NSCLC |
| irToxicity | irAE | Clinical Presentation |
|---|---|---|
| Gastrointestinal | Colitis | Diarrhea, abdominal pain, rectal bleeding, fever, nausea, and vomiting |
| Gastritis and esophagitis | Dyspepsia, epigastric pain, heartburn, reflux, nausea | |
| Cholecystitis | Abdominal pain at right upper quadrant, vomiting, diarrhea, and fever | |
| Pancreatitis | Epigastric pain, nausea, vomiting, fever, and diarrhea | |
| Duodenitis | Abdominal pain and diarrhea | |
| Hepatitis | Asymptomatic vs. mild (fever and rash) or severe (jaundice and liver failure) symptoms | |
| Endocrinologic | Thyroiditis | Usually asymptomatic in the first phases (tachycardia due to possible hyperthyroidism) followed by signs and symptoms of hypothyroidism. Anti-thyroid Abs always positive |
| Hypophysitis | Various combination of symptoms related to adrenal insufficiency, central hypothyroidism, hypogonadism, and/or central diabetes insipidus, along with headache and visual loss, due to possible mass effect with consequent cerebral oedema | |
| Cutaneous | Maculopapular rash | Faint erythematous macules and papules that coalesce into plaques; rashes in the trunk and extremities |
| Xerosis and eczematous conditions | Itchy, poorly demarcated, and erythematous macules and papules that coalesce into plaques and patches on the trunk and extremities; seborrheic dermatitis-like lesions on the face | |
| Psoriasis/psoriasiform eruption | Scaly erythematous plaques with well-defined borders on the trunk and extremities | |
| Lichen planus/lichen planus-like eruption | Rashes associated with itchy, red to violaceous, flat-topped papules, or plaques on the extremities and trunk, rarely on the oral mucosa | |
| Bullous diseases | Tense bullae accompanied by or preceded by pruritus; in some cases, only urticarial and eczematous plaques with pruritus | |
| Vitiligo | Flecked macules that coalesce into patches | |
| Alopecia | Well-defined patches or diffuse hair loss on the scalp, alopecia universalis | |
| Pruritus | May involve every body part, particularly trunk and scalp | |
| Stevens-Johnson syndrome and toxic epidermal necrolysis | High fever, widespread detachment of the epidermis, erosions, and mucositis | |
| DIHS/DRESS | Renal and hepatic impairment | |
| Pulmonary | Pneumonitis or ILD | Breathlessness and/or other respiratory symptoms (fever, cough, chest pain) |
| Sarcoidosis | Dry cough, dyspnea, fever, chronic fatigue, weight loss, chest pain | |
| Musculoskeletal and rheumatic | Inflammatory arthritis | Joint inflammation leading to articular pain, swelling, warmth, and stiffness (particularly if morning stiffness lasts more than 30 min). Laboratory tests, such as C-reactive protein and erythrocyte sedimentation rate, are often elevated. It could exhibit different clinical phenotypes, ranging from oligoarticular to polyarticular involvement but more often presenting symmetrically and involving both large and small joints (rheumatoid arthritis-like phenotype) |
| Polymyalgia rheumatica | Pain and stiffness primarily affecting the shoulders and/or hips, due to inflammation of periarticular structures | |
| Vasculitis | Inflammations of blood vessels; particularly but not limited to large-size blood vessels (e.g., giant cell arteritis) | |
| Sicca Syndrome | Marked by dryness of mucous membranes, particularly but not limited to the eyes and mouth. | |
| Hematological | autoimmune hemolytic anemia | Severe anemia with antiglobulin (Direct Coombs) test is usually positive for complement 3d or for IgG |
| Immune thrombocytopenia | Mostly asymptomatic severe abnormal platelet counts with possible haemorrhagic manifestations | |
| Immune-mediated neutropenia | Usually severe with bacterial and fungal infections | |
| Cardiovascular | Myocarditis | Shortness of breath, tachycardia or palpitations, fatigue, chest pain, cough, and episodes of syncope. Possible overlap with myocarditis, myositis, and/or myasthenia gravis |
| Pericarditis | Shortness of breath but also precordial pain, jugular venous congestion, or cardiogenic shock | |
| Arrythmias | Palpitations, fatigue, dyspnea, dizziness, or syncope, | |
| Nephrological | Acute kidney injury | increased creatinine levels, need for renal replacement therapy, sterile pyuria (500 white blood cells/hpf), eosinophilia (500 cells per L), haematuria, proteinuria |
| Electrolyte disorders | Hyponatraemia, hypokalaemia, metabolic acidosis | |
| Neurological | Myositis | Limb-girdle weakness with frequent bulbar, ocular, and facial involvement. Respiratory dysfunction and myocarditis in 30% of patients. Possible overlap with Myasthenia |
| Myasthenia Gravis | Muscle weakness and fatigue with daily fluctuations, frequent bulbar and respiratory involvement, presence of anti-AchR Abs. Possible overlap with myositis and myocarditis | |
| Acute/subacute demyelinating polyradiculonevritis | Predominant motor involvement and concomitant affection of cranial nerves. Frequent pleocytosis in cerebrospinal fluid | |
| Sensory neuronopathy | Sensory deficits with asymmetric onset, predominant involvement of upper limbs, often associated with anti-onconeural Abs | |
| Cranial neuropathy | All cranial nerves may be affected. More often, facial palsy or painless neuritis | |
| Focal encephalitis | Limbic encephalitis or Brainstem Encephalitis (cerebellar syndromes are less common). Onconeural Abs are often detectable (++ anti-Ma2-Abs or anti-Hu Abs) | |
| Meningitis and meningoencephalitis | Fever, meningeal signs with or without encephalopathy. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casagrande, S.; Sopetto, G.B.; Bertalot, G.; Bortolotti, R.; Racanelli, V.; Caffo, O.; Giometto, B.; Berti, A.; Veccia, A. Immune-Related Adverse Events Due to Cancer Immunotherapy: Immune Mechanisms and Clinical Manifestations. Cancers 2024, 16, 1440. https://doi.org/10.3390/cancers16071440
Casagrande S, Sopetto GB, Bertalot G, Bortolotti R, Racanelli V, Caffo O, Giometto B, Berti A, Veccia A. Immune-Related Adverse Events Due to Cancer Immunotherapy: Immune Mechanisms and Clinical Manifestations. Cancers. 2024; 16(7):1440. https://doi.org/10.3390/cancers16071440
Chicago/Turabian StyleCasagrande, Silvia, Giulia Boscato Sopetto, Giovanni Bertalot, Roberto Bortolotti, Vito Racanelli, Orazio Caffo, Bruno Giometto, Alvise Berti, and Antonello Veccia. 2024. "Immune-Related Adverse Events Due to Cancer Immunotherapy: Immune Mechanisms and Clinical Manifestations" Cancers 16, no. 7: 1440. https://doi.org/10.3390/cancers16071440
APA StyleCasagrande, S., Sopetto, G. B., Bertalot, G., Bortolotti, R., Racanelli, V., Caffo, O., Giometto, B., Berti, A., & Veccia, A. (2024). Immune-Related Adverse Events Due to Cancer Immunotherapy: Immune Mechanisms and Clinical Manifestations. Cancers, 16(7), 1440. https://doi.org/10.3390/cancers16071440