Abstract
The exacerbation of climatic changes helped to increase the risk of plant diseases in the world. The novelty of this study lies in the manufacture of therapeutic nutrients using nanotechnology with strong effectiveness against plant disease. Based on this concept, we mycosynthesized bimetallic ZnO-CuO nanoparticles (NPs), alternatives to reduce the spread of Vicia faba Fusarium wilt disease, which is one of the world’s most imperative cultivated crops. The article’s uniqueness comes in the utilization of ZnO-CuO nanoparticles to carry out two crucial tasks: therapeutic nutrients and managing Fusarium disease. To evaluate the resistance of infected plants, disease index (DI), photosynthetic pigments, osmolytes, oxidative stress and yield parameters were assessed. NPs of ZnO, CuO, and ZnO-CuO were mycosynthesized using a biomass filtrate of Aspergillus fumigatus OQ519856. DI reached 87.5%, due to Fusarium infection, and, as a result, a severe decrease in growth characters, photosynthetic pigments, total soluble carbohydrates, and proteins as well as yield parameters was observed. Infected plants produced more of the studied metabolites and antioxidants. On the other hand, the treatment with CuO-ZnO NPs led to a great decline in the DI by 22.5% and increased the protection by 74.28%. A clear improvement in growth characters, photosynthetic pigments and a high content of carbohydrates and proteins was also observed in both healthy and infected plants as a result of CuO-ZnO NPs treatment. Remarkably, CuO-ZnO NPs significantly increased the yield parameters, i.e., pods/plant and pod weight, by 146.1% and 228.8%, respectively. It could be suggested that foliar application of NPs of ZnO, CuO, and ZnO-CuO could be commercially used as antifusarial agents and strong elicitors of induced systemic resistance.
1. Introduction
Faba bean is one of the essential nutritious popular food crops in the world and plays a main role in the Egyptian cuisine. However, it faces several biotic constrains including viral, bacterial, fungal infections, as well as various pests and insects [1]. Many fungal infections that impact faba bean production as well as other crops in various locations are classified as root system diseases [2,3]. Fusarium is one of these disorders which has a definite effect on the plant’s autoimmunity and consequently causes a decrease in crop productivity and quantity during the growing season. [4]. Given the difficulty and harm of completely controlling Fusarium wilt disease chemically, the most appropriate alternative was to adopt a highly efficient natural approach to combat wilt disease, which is also less expensive and eco-friendly. Furthermore, climatic change has lately intensified, resulting in the rapid spread of plant diseases [4,5].
In fact, the induction of systemic resistance in the plant makes the plant able to recover from the damage of pathogens. Systemic resistance or immunity can be induced in plants by biotic or abiotic inducers [6]. NPs improve structural and metabolic parameters and greatly increase yield by generating systemic resistance. [7,8]. By encouraging growth metrics and metabolic properties, ZnO-NPs have been suggested as a synthetic fungicide substitute for eliminating Fusarium wilt disease by inducing systemic resistance [9]. The possibility of nanoparticles to induce SAR as a practical approach of controlling numerous plant diseases is only now being realized [10]. In order to treat fungal infections, the use of nanobiotechnology to resist infections and boost plant physiological immunity has produced significant and effective results. This is because it might boost plant systemic immunity, disease resistance, and yield [11,12,13,14]. In this regard, biosynthesis of nanomaterials using fungal extracts, a plentiful source of natural compounds such as carbohydrates, phenols, flavonoids, tannins and alkaloids, is a promising approach that can operate as a safe reducing and stabilizing agent [15].
Zinc oxide nanoparticles play an important role in the process of sugar oxidation in plants [7]. Nano CuO applications increased plant health, growth, and production, and have the potential to have a significant impact on the management of plant diseases in ornamentals [16,17]. Attention turned to feeding the plant with compounds that contain copper, as scientific reports have proven the antimicrobial efficiency of copper, as it stimulates the activation of laccases, which is one of the enzymatic analogues of polyphenol oxidase that breaks down and oxidizes phenolic substances. It has been reported that in vitro application of copper on potato plants infected with brown mold bacteria inhibited the bacterial growth. It is worth noting that plants treated with copper showed a significant improvement in morphological characteristics and physiological immune responses. This article’s novelty stems from the use of bimetallic ZnO-CuO nanoparticles to perform two critical tasks: therapeutic nutrition delivery and Fusarium wilt management with high efficacy. The main goal of this research is to recover the plant’s fighting in contradiction of Fusarium wilt disease in faba beans plant caused by Fusarium oxysporum using nano-biotechnology represented by bimetallic ZnO-CuO NPs as a nano-fertilizer against the fungal plant pathogen.
2. Materials and Methods
2.1. Biosynthesis of NPs
NPs were myco-synthesized using a filtrate of A. fumigatus OQ519856 and the complete characterization of NPs of ZnO, CuO, and ZnO-CuO (such as UV-Vis., HR-TEM, DLS, and SEM analysis) was recorded in our previous study [18]. Moreover, the concentration used of ZnO-CuO NPs (125 µg/mL) ZnO NPs (1000 µg/mL) and CuO NPs (500 µg/mL) depends on the MIC against fusarium, as mentioned in previous published article [18].
2.2. Source of the Fusarium Pathogen
The pathogen was obtained from the Regional Center for Mycology and Biotechnology (RCMB) at Al-Azhar University. The pathogen was cultured on PDA media for 5 days before being stored at 4 °C, and the pathogenic fungus inoculum was ready.
2.3. In Vivo Experiment
2.3.1. In Vivo Study
The present study employed seeds of Vicia faba L, namely the CV Giza 3 variety.
The seeds were immersed in the nano solutions (125 μg/mL) for a duration of 2 h, with a ratio of 2 mL of solution per gram of seeds. These treatments were employed in the experimental procedure. A single seed/pot, measuring 30 cm in diameter, was planted in the Research Garden located at the Faculty of Science, Al-Azhar University in Cairo, Egypt. The pot included a mixture of sand and clay in a weight-to-weight ratio of 1:3, with a total mass of 4 kg. The distribution of the pots was carried out, ensuring that each treatment had 8 repetitions.
The experimental treatments were organized as follows: T1 represented the healthy control group, T2 represented the group of infected plants that were left untreated, T3 represented the group of healthy plants that were treated with zinc oxide nanoparticles (ZnO NPs), T4 represented the group of infected plants that were treated with ZnO NPs, T5 represented the group of healthy plants that were treated with copper oxide nanoparticles (CuO NPs), T6 represented the group of infected plants that were treated with CuO NPs, T7 represented the group of healthy plants that were treated with a combination of ZnO-CuO NPs, and T8 represented the group of infected plants that were treated with ZnO-CuO NPs.
The pathogenic fungus was artificially infected by injecting 10 mL of Fusarium oxysporum 107 spores/ mL) into the soil. After germination and the appearance of the cotyledons of the plants, the plants’ (healthy and infected) treatments with nanosolutions was started. Three doses (20 mL per plant, once per week) of nanosolutions at a concentration of (125 g/mL) were applied to the plants’ leaves using a one-handed pressure irrigator. Assessment of plant resistance, recording disease symptoms, as well as taking plant samples for biochemical tests were conducted 60 days after germination.
A duration of 60 days after germination, symptoms were recorded, and disease severity and protection were determined according to Attia et al. [19].
2.3.2. Metabolic Indicators for Resistance
The lengths of the shoots were measured at random from above the soil surface to the end of the plant’s growing tip and recorded in cm. Root lengths were measured from the soil surface to the end of the root tip and recorded in cm. Chlorophyll and carotenoids were measured using the technique of Abdelaziz et al. [20]. A solution consisting of 50 mL of acetone with an 80% concentration was employed to extract photosynthetic pigments from a sample of fresh leaves weighing 0.5 g. The filtrate was subsequently quantified by spectrophotometric analysis at wavelengths of 665 nm, 649 nm, and 470 nm. The estimation of the soluble protein was conducted by Lowry et al. [21]. A volume of 1 mL of the extract was mixed with 5 mL of an alkaline reagent consisting of 50 mL of a 2% Na2CO3 solution prepared in 0.1 N NaOH and 1 mL of a 0.5% CuSO4 solution prepared in 1% potassium sodium tartrate. Additionally, 0.5 mL of Folin’s reagent, diluted by a ratio of 1:3 v/v, was added to the mixture. After a duration of 30 min, an observable alteration in color was detected at a wavelength of 750 nanometers.
The quantification of soluble carbohydrates in the shoot was determined using the methodology outlined in the study conducted by Umbreit et al. [22]. The dried shoots from each treatment, weighing 0.5 g, were collected and mixed with 2.5 mL of a 2% phenol solution and 5 mL of a 30% trichloroacetic acid (TCA) solution. The resulting mixture was subsequently filtered using filter paper. Subsequently, a volume of 2 mL of anthrone reagent, prepared by dissolving 2 g of anthrone in 1 L of 95% sulfuric acid (H2SO4), was introduced. The color that was observed and measured had a wavelength of 620 nm, with a blue-green color.
The content of free proline was established by the method of Bates et al. [23]. The protocol mentioned in Dai et al. [24] was used to assess the total phenolics. The method of Hu, Richter [25] was used to determine the amount of malondialdehyde (MDA) in fresh faba bean leaves. Fresh faba bean leaves were tested for hydrogen peroxide H2O2 content [26]. The approved technique of Srivastava, [27] was used to determine peroxidase activity. The activity of polyphenol oxidase was measured by the method of Matta [28].
2.3.3. Statistical Analyses
The data were subjected to one-way variance analysis (ANOVA). The LSD test by CoStat was used to reveal statistically significant differences between treatments at p 0.05. The resulting data are shown as mean standard errors (n = 3).
3. Results and Discussion
3.1. V. faba Protection
The results in Table 1 and Figure 1 showed that the fungus infects the roots of the plant, causing wilting, yellowing, and eventually death. F. oxysporum was highly virulence and caused 87.5% DI. The results showed that both ZnO NPs and CuO NPs alone efficiently reduced the severity of the Fusarium wilt disease, with average DIs (50% and 37.5%) and protections (42.8% and 57.1%), respectively. However, the combined treatment of ZnO-CuO NPs was even more effective, with a DI (22.5%) and protection of 22.5% and 74.28%, respectively. This suggests that the combination of these two NPs could be a promising approach for controlling V. faba Fusarium wilt disease.

Table 1.
Effect NPs of ZnO, CuO, and ZnO-CuO on disease index of F. oxysporum wilt disease of V. faba.

Figure 1.
Symptoms of wilt disease: (A) untreated infected, (B) untreated healthy, (C) infected treated with ZnO NPs, (D) infected treated with CuO NPs and (E) infected treated with ZnO-CuO NPs.
ZnO NPs have been reported to possess strong direct fungicidal effects against numerous plant pathogenic fungi, including F oxysporum, through their high surface-area-to-volume ratio that enables them to penetrate the fungal cell wall and disrupt its metabolic processes, leading to the inhibition of fungal growth.
These results are explained indirectly by the fact that ZnO NPs are some of the important elements that stimulate plants to synthesize growth hormones and repair hormone imbalances [29]. The researchers used CuO NPs in laboratories to fight fusarium, and the results showed that the use of CuO NPs inhibited fungal growth [30]. The present study also found that the combination treatment of ZnO and CuO NPs was more effective than either NP alone, indicating a potential synergistic effect between the two NPs (Zn and Cu). This is consistent with previous studies that have also demonstrated that the combination of metal NPs can have a synergistic effect on their antifungal properties against phytopathogenic fungi [31].
Growth Biomarkers
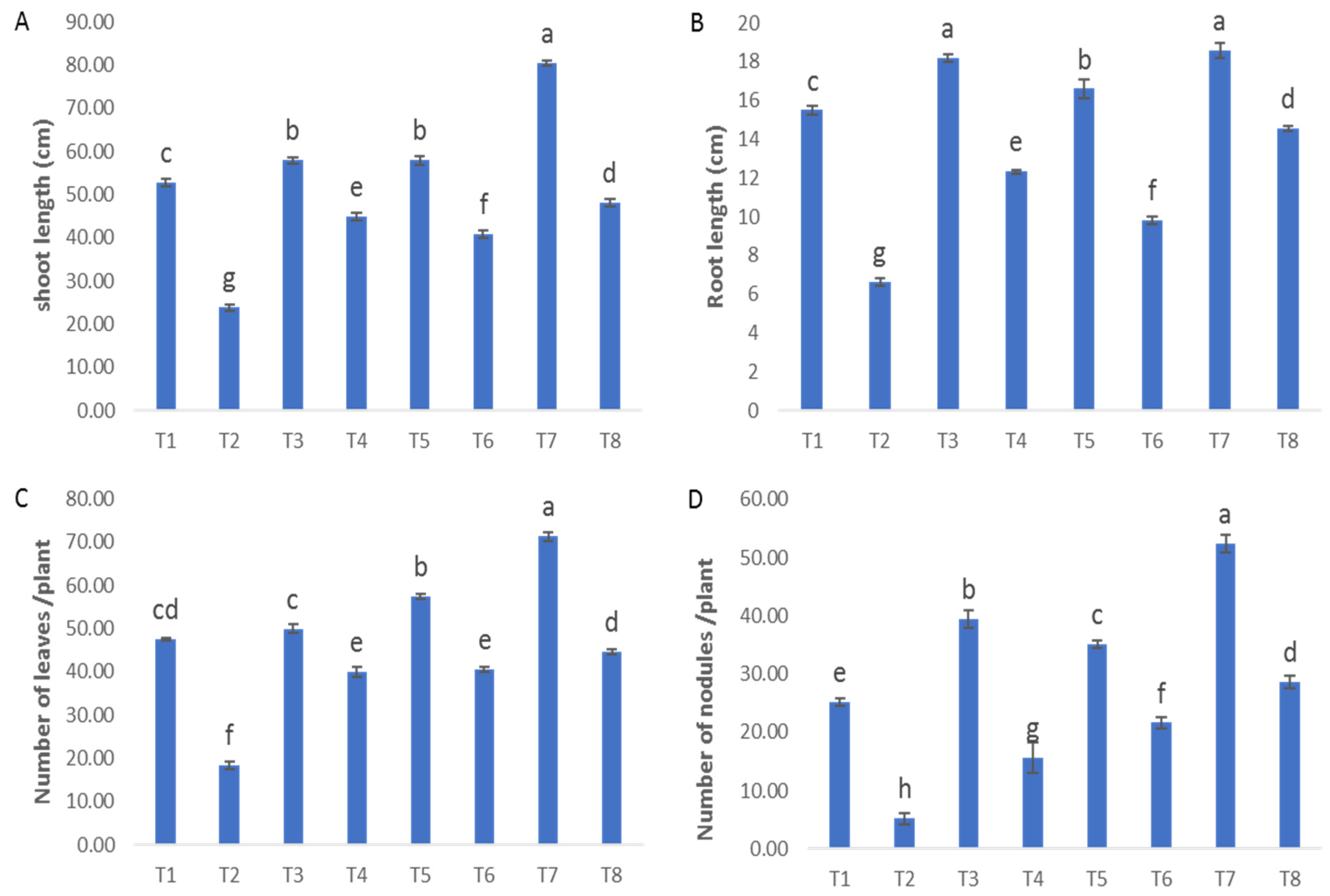
It is evident from Figure 2A–C that Fusarium-infected plants exhibited great reductions in plant height (54.8%), root length (57.8%), number of leaves (61.28%) and number of nodules (79.3%) compared to the control plants. These data agree with the recent research by Elsakhawy, T. et al. [32]. Fusarium infection was reported to clog the vessels carrying water and salts, so the plants become dwarfs and eventually die [33]. Plants that were both healthy and diseased and were subjected to treatment with ZnO-CuO nanoparticles (NPs), ZnO NPs, and CuO NPs exhibited noteworthy increases in many growth parameters, including plant height, root length, number of leaves, and number of nodules when compared to plants that did not receive any treatment. In the investigation of the impact of ZnO nanoparticles (NPs), CuO nanoparticles (NPs), and ZnO-CuO nanoparticles (NPs) on plants in a healthy state, it was observed that the treatment involving ZnO-CuO NPs exhibited the most notable and statistically significant enhancements in several plant growth parameters. Specifically, this treatment resulted in a substantial increase in plant height (52.59%), root length (19.18%), number of leaves (50.01%), and number of nodules (108%), as depicted in Figure 2A–C. Concerning the effect of ZnO NPs, CuO NPs and ZnO-CuO NPs on the challenged plants with F. oxysporum, it was found that ZnO-CuO NPs were the best treatment that showed the greatest significant increase in plant height (101%), root length (119%), number of leaves (142%) and number of nodules (450%). Such a remarkable enhancement in growth parameters could be attributed to the ability of the used treatments (ZnO-CuO NPs, ZnO NPs and CuO NPs) to improve the plant’s defense as well as their anti-Fusarium ability that prevent the fungal penetration into the plant roots. Moreover, the treatments enhanced the formation of nitrogen fixation nodules, and the release of secondary metabolites, thus facilitating nutrient uptake from the root environment. These results are supported by Ragab, Turoop [34]; they reported that treatment with ZnO NPs resulted in a significant improvement in the morphological characteristics of faba bean plants. García-López et al. [35] stated that zinc oxide has the potential to elevate the yield and growth of some crops. Nanoparticles can travel long distances through the plant’s vasculature after entering the leaf apoplast [36]. It is noted that treatment with CuO NPs resulted in an improvement in the plant’s morphological characteristics, as well as an increase in the plant’s defensive ability [37].
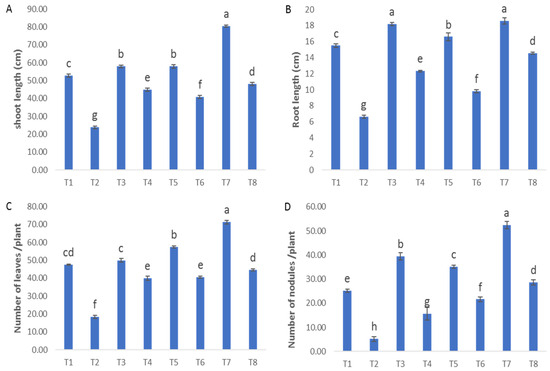
Figure 2.
Effect of NPs of ZnO, CuO, and ZnO-CuO on (A) shoot length, (B) root length, (C) number of leaves and (D) number of nodules of F. oxysporum wilt disease of faba bean plant. T1: healthy control; T2: infected control; T3: healthy and treated with ZnO NPs; T4: infected and treated with ZnO NPs; T5: healthy and treated with CuO NPs; T6: infected and treated with CuO NPs; T7: healthy and treated with ZnO-CuO NPs; T8: infected and treated with ZnO-CuO NPs. (Data represent mean ± SD, n = 3, letters “a–h” refer to significance in statically analysis).
3.2. Photosynthetic Pigments
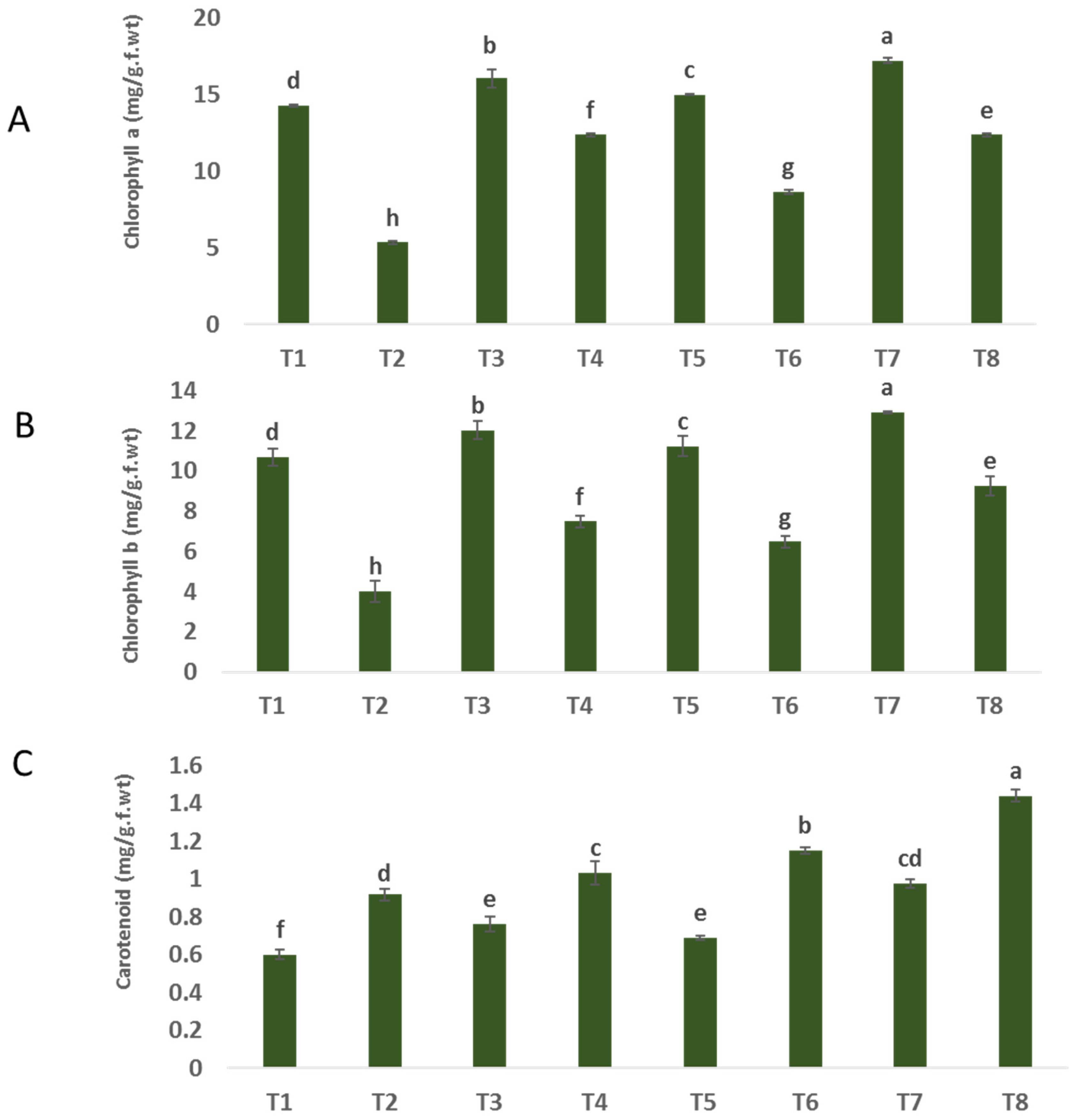
The findings depicted in Figure 3A–C indicate a significant decrease in the levels of chlorophyll a and b in plants infected with F. oxysporum (5.34 and 5.007 mg/g. f.wt) compared to uninfected plants (14.28 and 9.71 mg/g. f.wt), respectively. The significant reduction in chlorophyll pigment content can be attributed to the plant’s inability to effectively catch light and perform photosynthesis, as well as the impact of oxidative bursts on chloroplasts caused by disease infection [38]. Plants that were both healthy and sick and subjected to treatment with nanoparticles (NPs) of ZnO, CuO, and ZnO-CuO exhibited notable increases in the levels of chlorophyll a and b, in comparison to plants that did not receive any treatment. In relation to the impact of treatments on plants affected by F. oxysporum, it was observed that the application of ZnO-CuO nanoparticles (NPs) yielded the most favorable results. This treatment exhibited a substantial and statistically significant increase in the levels of chlorophyll a (131.46%) and chlorophyll b (131.37%) when compared to the effects of ZnO NPs (86.70% and 86.74%) and CuO NPs (61.98% and 61.71%), respectively. The findings depicted in Figure 2A–C indicate a notable increase in carotenoid levels in plants that were infected. In the case of Fusarium-infected plants, the application of zinc oxide (ZnO), copper oxide (CuO), and a combination of ZnO and CuO nanoparticles (NPs) resulted in an observed rise in carotenoid levels as compared to non-treated plants infected with F. oxysporum, as depicted in Figure 2A–C.
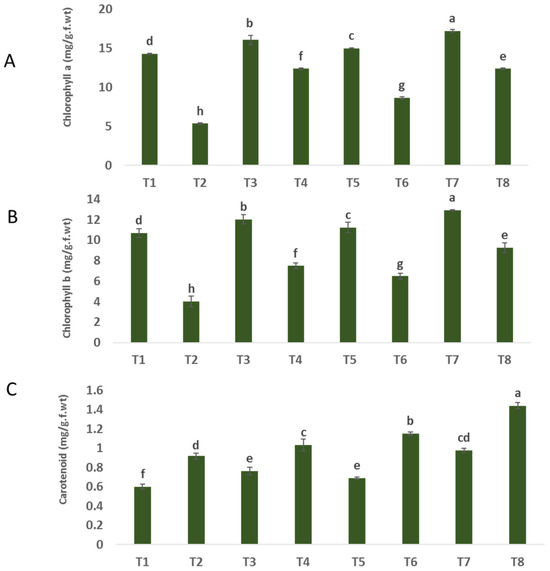
Figure 3.
Effect of NPs of ZnO, CuO, and ZnO-CuO on (A) chlorophyll a, (B) chlorophyll b and (C) carotenoids of F. oxysporum wilt disease of faba bean plant. (Data represent mean ± SD, n = 3, letters “a–h” refer to significance in statically analysis).
Also, the achieved results showed that in both healthy and F. oxysporum-infected plants, contents of carotenoids were increased in response to the treatment with ZnO-CuO NPs, ZnO NPs and CuO NPs. Increased chlorophyll contents in infected plants treated with ZnO NPs, CuO NPs and ZnO-CuO NPs could have resulted from supplying the plant with zinc and copper, where ZnO NPs play an important role in cell division [39,40], and CuO NPs in the formation of chlorophyll [41], as scientific reports have proven that up to 70% of the copper found in plants is found in chlorophyll, which is largely associated with chloroplasts [42,43]. It is involved in lignin formation, and protein and carbohydrate synthesis, and may be necessary for symbiotic nitrogen fixation [44]. Copper is part of the plastocyanin, which forms a link in the electron transport chain and is involved in photosynthesis [45].
3.3. Osmolytes
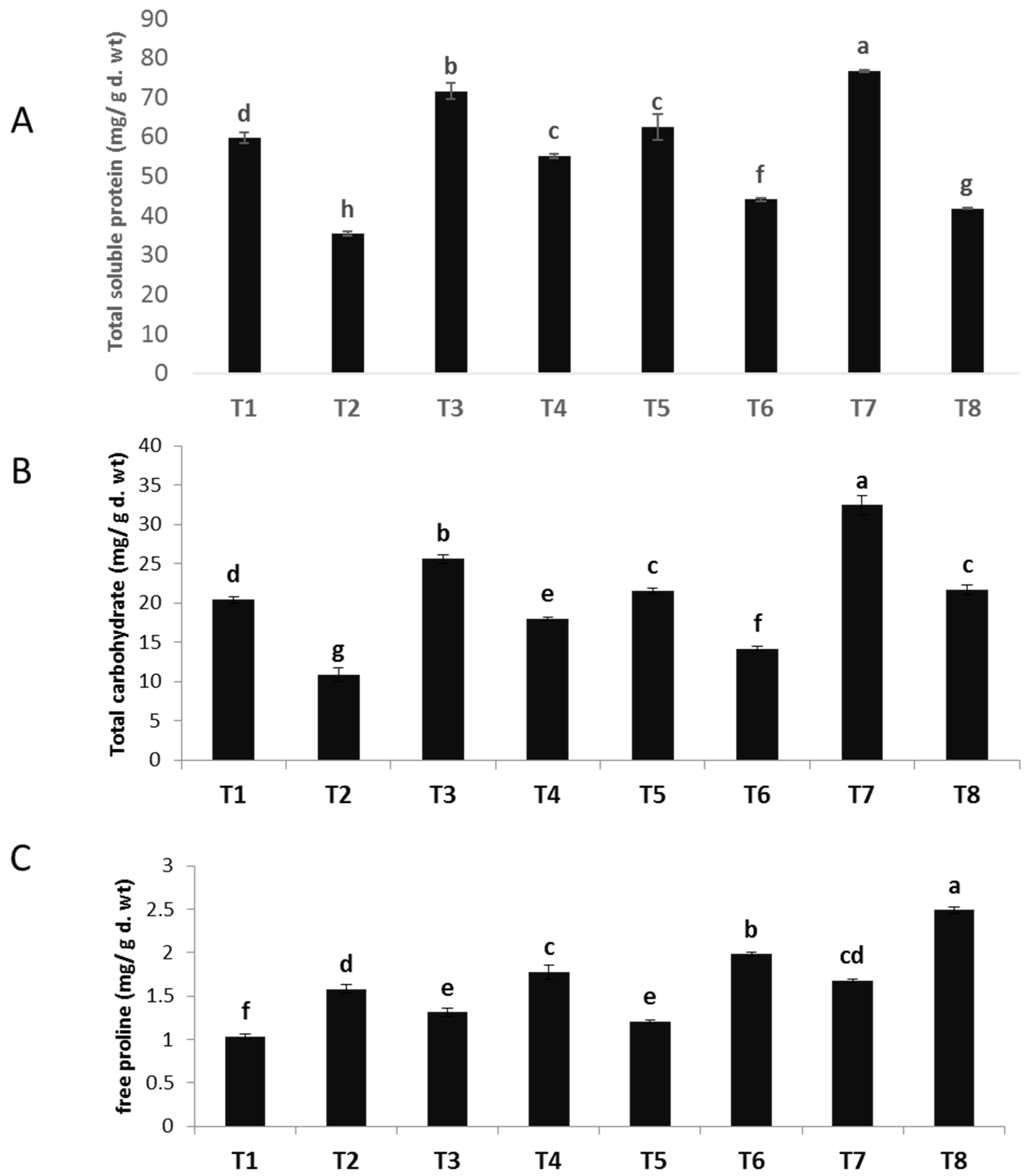
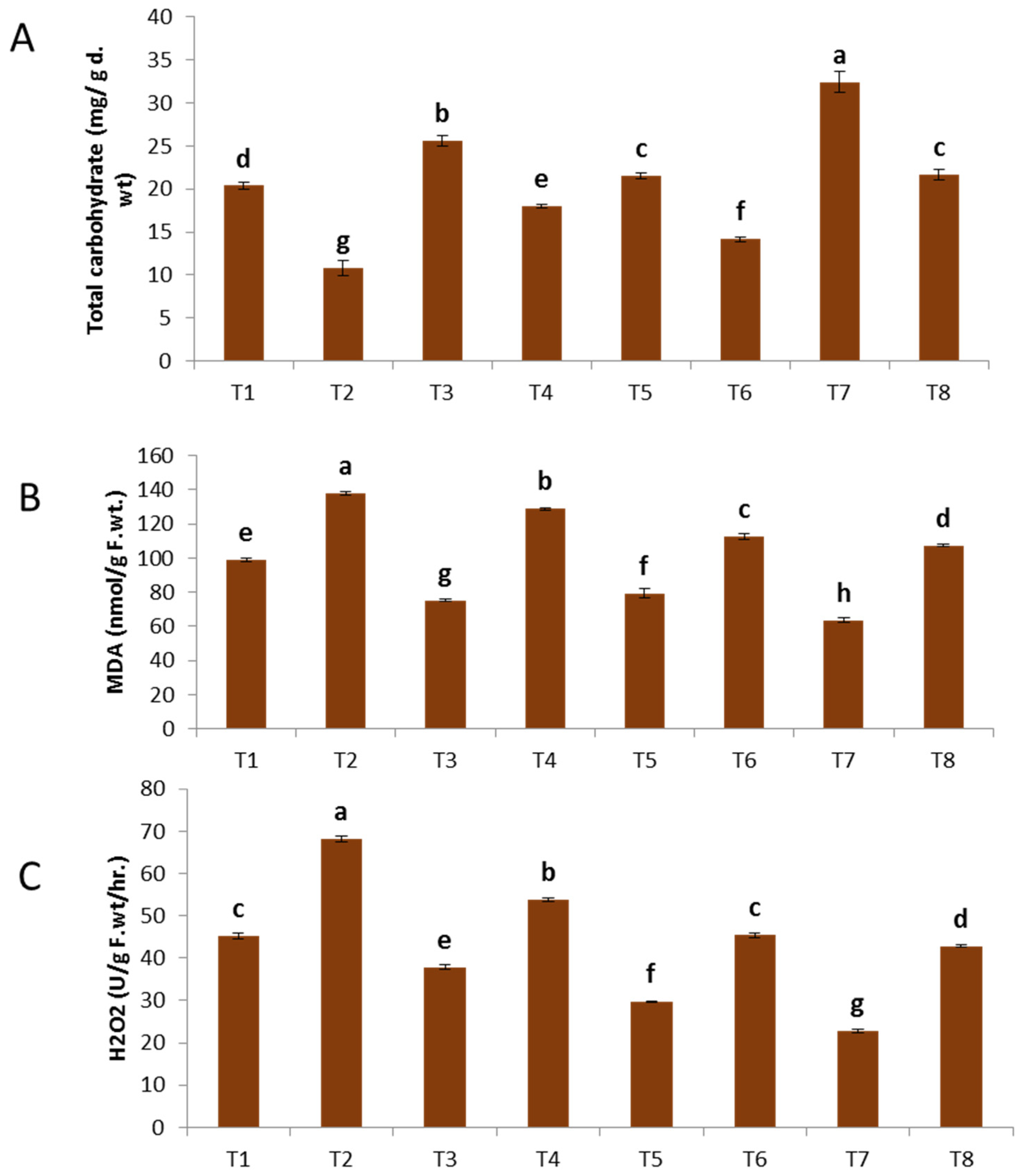
The data depicted in Figure 4A–C demonstrate a considerable decrease in the total soluble protein and carbohydrate content of faba bean as a result of F. oxysporum infection. These findings are corroborated by numerous prior investigations [46]. In contrast, a study revealed that the use of zinc oxide nanoparticles (ZnO NPs), copper oxide nanoparticles (CuO NPs), and a combination of ZnO-CuO NPs on both healthy and diseased plants resulted in a noteworthy augmentation in total soluble protein and carbohydrate levels, as compared to plants that were not treated. Regarding the impact of ZnO nanoparticles (NPs), CuO nanoparticles (NPs), and ZnO-CuO nanoparticles (NPs) on plants infected with F. oxysporum, it was observed that ZnO NPs exhibited a significant increase in the overall soluble protein content (55.90%). This was followed by CuO NPs (24.70%) and ZnO-CuO NPs (18.34%), in comparison to the infected control group that remained untreated. In relation to the impact of ZnO nanoparticles (NPs), CuO nanoparticles (NPs), and ZnO-CuO nanoparticles (NPs) on plants exposed to F. oxysporum, it was observed that the bimetallic ZnO-CuO NPs exhibited a significant rise in total soluble carbohydrates (99.35%). This was followed by ZnO NPs (65.71%) and CuO NPs (30.50%), when compared to the infected control group that was not treated. These findings are consistent with the findings reported in reference [47]. According to a recent study [48], zinc has been identified as a constituent of glyco-dehydrogenases, which play a crucial role in protein synthesis, as well as glycine dipeptidases, which are essential for the process of glycolysis. Copper assumes a crucial function in the process of chlorophyll creation and metabolism within plant cells, so it serves as a significant contributor in promoting the synthesis of proteins and carbohydrates.
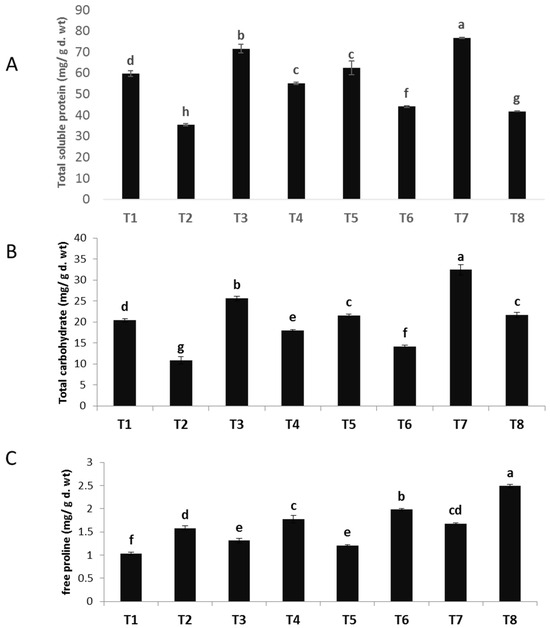
Figure 4.
Effect of NPs of ZnO, CuO, and ZnO-CuO on (A) total soluble protein, (B) total soluble carbohydrates and (C) free proline contents of F. oxysporum wilt of V. faba. (Data represent mean ± SD, n = 3, letters “a–h” refer to significance in statically analysis).
The quantification of proline levels in plant tissues serves as an indicator of the host plants’ response to infection and its possible resistance to pathogens. The findings of the current study (Figure 4A–C) demonstrated notable elevations in the levels of free proline in plants infected with Fusarium. The utilization of nanoparticles (NPs) derived from ZnO, CuO, and ZnO-CuO has demonstrated notable enhancements in the levels of free proline in both uninfected and diseased plants. Regarding the impact of nanoparticles (NPs) composed of ZnO, CuO, and ZnO-CuO on plants exposed to F. oxysporum, it was shown that bimetallic ZnO-CuO NPs exhibited a noteworthy elevation in proline levels (57.59%) compared to CuO NPs (25.31%) and ZnO NPs (12.65%). The present findings indicate a clear correlation between the reduction in primary metabolite levels, specifically soluble carbohydrates and soluble proteins, and the severe depletion of chlorophylls in response to Fusarium infection. Nevertheless, the proline level exhibited an increase as a result of its involvement in osmoregulation and the scavenging of reactive oxygen species (ROS) [49].
3.4. Oxidative Stress
The data shown in Figure 5A–C demonstrate that the presence of F. oxysporum resulted in a substantial and statistically significant elevation in the total phenolic content of the infected plants. A noteworthy observation was made regarding the treatment of plants with nanoparticles (NPs) of zinc oxide (ZnO), copper oxide (CuO), and a combination of ZnO and CuO (ZnO-CuO). This treatment resulted in a substantial increase in the overall phenolic content of both healthy plants and those infected with F. oxysporum. In relation to the impact of ZnO, CuO, and ZnO-CuO nanoparticles (NPs) on plants exposed to F. oxysporum, it was shown that the bimetallic ZnO-CuO NPs exhibited a more pronounced elevation in total phenolic content compared to both ZnO NPs and CuO NPs individually.
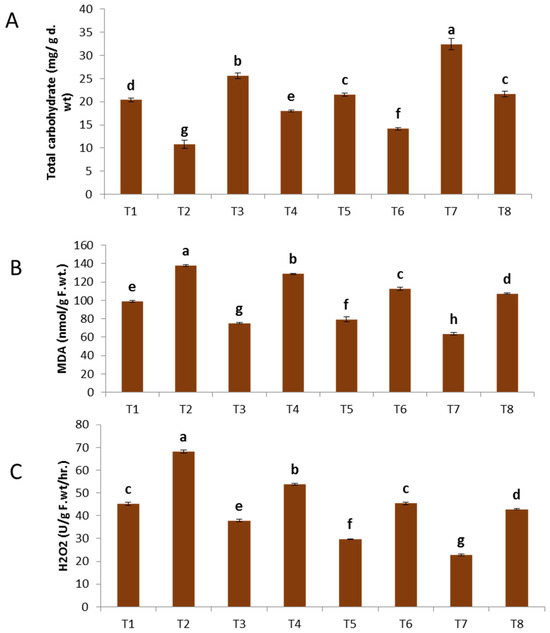
Figure 5.
Effect of NPs of ZnO, CuO, and ZnO-CuO on (A) total phenol, (B) MDA and (C) H2O2 levels of F. oxysporum wilt disease of faba bean plant. (Data represent mean ± SD, n = 3, letters “a–h” refer to significance in statically analysis).
The infection caused by F. oxysporum resulted in the buildup of MDA (137.94 nmol/g f.wt) and H2O2 (68.12) in comparison to the untreated plants, as shown in Figure 2B. The aforementioned results are corroborated by a multitude of prior research studies [50]. The concentration of malondialdehyde (MDA) was seen to decrease upon exposure to bimetallic ZnO-CuO nanoparticles, as well as individual CuO nanoparticles and ZnO nanoparticles. The respective concentrations of MDA were measured as 107.38, 112.59, and 128.84 n mol/g f.wt. The concentration of H2O2 exhibited a decrease of 42.81%, 45.19%, and 53.79% when compared to the untreated infected plant, as depicted in Figure 5C.
The utilization of bimetallic ZnO-CuO nanoparticles resulted in a reduction in the generation of malondialdehyde (MDA) and hydrogen peroxide (H2O2) by enhancing the presence of antioxidant chemicals that effectively scavenge reactive oxygen species (ROS) and safeguard cellular membranes. Avoidance and reduction of oxidative stress, as well as the capture of free radicals, are considered significant indicators of resistance to stress [51]. The findings of this study are consistent with prior research that has shown a reduction in the concentrations of MDA and H2O2 following the administration of CuO nanoparticles and bimetallic ZnO-CuO nanoparticles [52].
3.5. Antioxidant Enzyme Activity
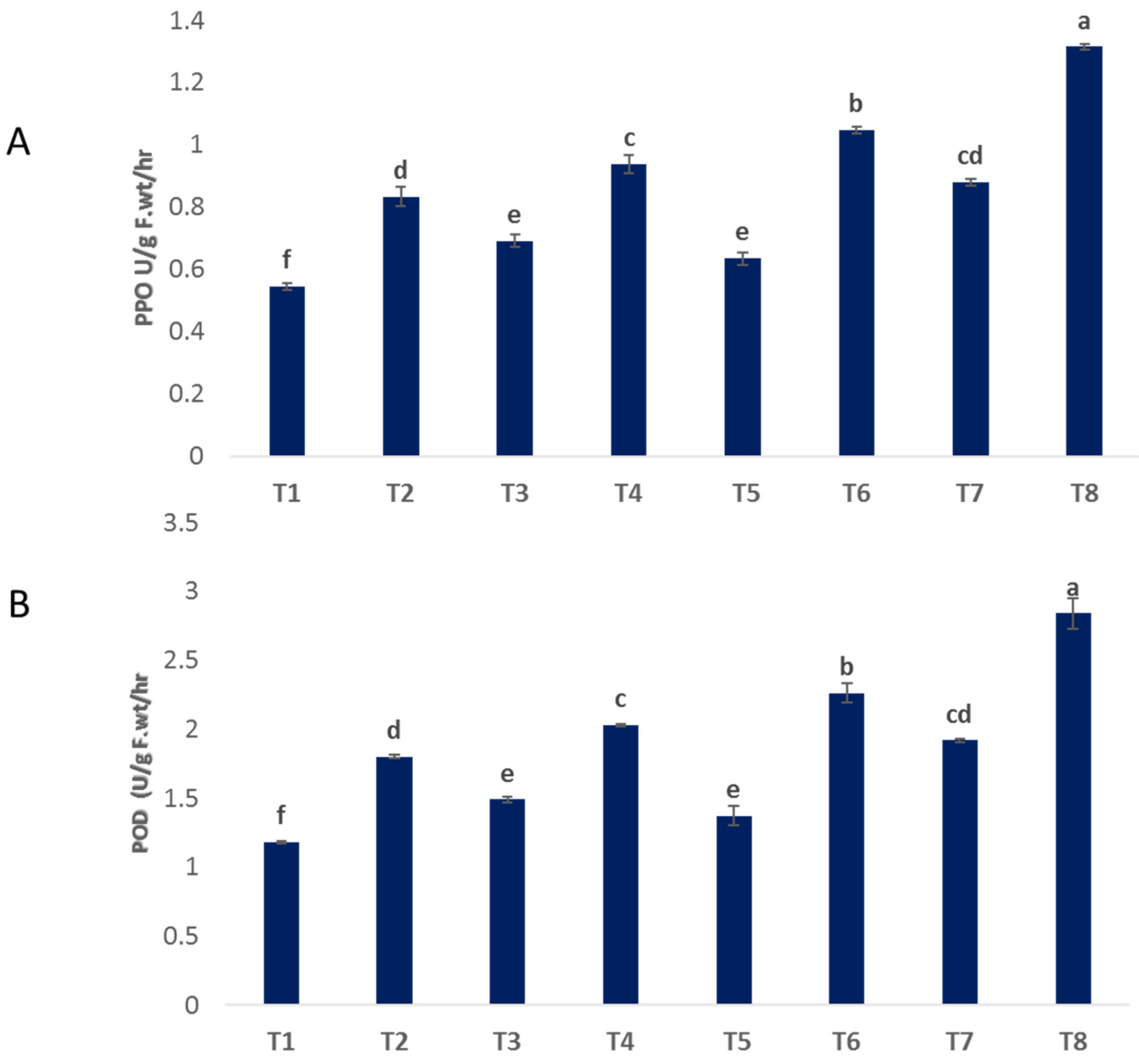
The activity of peroxidase (POD) and polyphenol oxidase (PPO) was increased in infected plants that recorded as 1.801 and 0.834, respectively, compared to untreated control plants (1.179 and 0.545) (Figure 6A,B). Moreover, spraying with bimetallic ZnO-CuO NPs, CuO NPs and ZnO NPs increased the activity of POD by 2.841, 2.261, and 2.026, respectively, and PPO by 1.315, 1.047, and 0.938, respectively, over infected plants (Figure 6A,B). Under non-infected conditions, the foliar administration of nanoparticles (NPs) containing zinc oxide (ZnO), copper oxide (CuO), and a combination of ZnO and CuO resulted in enhanced activities of polyphenol oxidase (PPO) and peroxidase (POD) in the treated plants, when compared to the control group. Enhancing the activity of antioxidant enzymes is a crucial factor in bolstering plant physiological immunity and safeguarding cellular integrity against oxidative damage caused by infections [53].
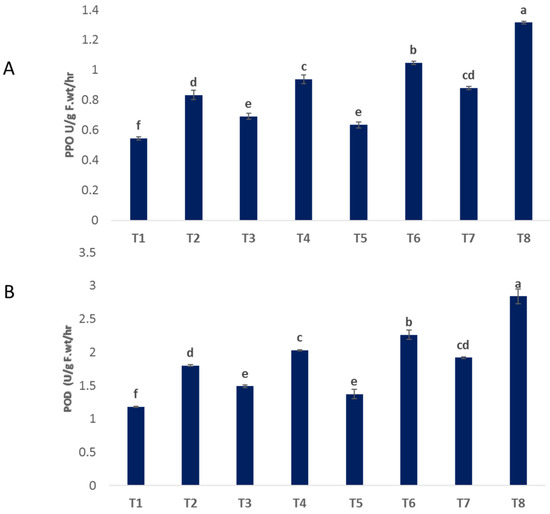
Figure 6.
Effect of NPs of ZnO, CuO, and ZnO-CuO on antioxidant enzyme activity (POD (A) and PPO (B)) of F. oxysporum wilt disease of faba bean plant. (Data represent mean ± SD, n = 3, letters “a–f” refer to significance in statically analysis).
3.6. Yield Characters
Results of the present study (Table 2) revealed that F. oxysporum caused highly significant decrease in the number of pods/plant (17.91%) and weight of pods (24.95%). The results of this study agree with a recent work [54], where they reported that infection with Fusarium leads to a severe decrease in yield. On the other hand, application of ZnO NPs, CuO NPs and ZnO-CuO NPs resulted in a highly significant increase in yield characters in both healthy and infected plants. However, treatment with bimetallic ZnO-CuO NPs caused a highly significant increase in the amount of pods/plant (966.51%) and weight of pods g/plant (486.08%). Spraying of zinc on bean plants led to increases in all yield components [55].
Table 2.
Effect of NPs of ZnO, CuO, and ZnO-CuO on yield characters of faba bean plant.
4. Conclusions
The study provides promising results on the potential use of myco-synthetic bimetallic ZnO-CuO NPs as an alternative agent to control the spread of Fusarium wilt disease and improving the growth and yield parameters (photosynthetic pigments, proline, phenolics, antioxidant enzymes, MDA, H2O2, and number of pods and weight of pods of Vicia faba plants. The results showed that both ZnO NPs and CuO NPs alone efficiently reduced the severity of the fusarium wilt disease, with average DIs (50% and 37.5%) and protections (42.8% and 57.1%), respectively. Healthy and infected plants treated with NPs of ZnO, CuO, and ZnO-CuO presented significant rises in the plant height, root length, number of leaves and number of nodules, compared with non-treated plants ones. Concerning the effect NPs of ZnO, CuO, and ZnO-CuO on the challenged plants with F. oxysporum, it was found that ZnO-CuO NPs were the best treatment that showed the greatest significant increase in the contents of chlorophyll a by 131.46% and chlorophyll b by 131.37%, in comparison with ZnO NPs (86.70% and 86.74%), and CuO NPs (61.98% and 61.71%), respectively. Concerning the effect NPs of ZnO, CuO, and ZnO-CuO on the challenged plants with F. oxysporum, it was found that ZnO NPs show a considerable increase in total soluble protein (55.90%), followed by CuO NPs (24.70%) and ZnO-CuO NPs (18.34%), when being compared with untreated infected control. The content of MDA was reduced in response to bimetallic ZnO-CuO NPs, CuO NPs and ZnO NPs to 107.38, 112.59, and 128.84, n mol/g f.wt., respectively. While the content of H2O2 declined by 42.81, 45.19 and 53.79, respectively, compared to the untreated infected plant. Moreover, spraying with NPs of ZnO, CuO, and ZnO-CuO increased the activity of POD by 2.841, 2.261, and 2.026, respectively, and PPO by 1.315, 1.047, and 0.938, respectively, over infected plants. The outcomes showed that actino-synthesized ZnO-CuO NPs might increase plant biochemical resistance while also inhibiting F. oxysporum wilt impacts on faba bean plants. However, this study needs to be undertaken to fully understand the environmental and health impacts of the use of nanoparticles in agriculture.
Author Contributions
Conceptualization, M.S.A., A.H.H., G.S.E.-S., A.M.A. and A.M.A.K.; methodology, M.S.A., A.H.H., G.S.E.-S., A.M.A., S.E.G., O.M.A.z., A.M.S. and A.M.A.K.; software, M.S.A., A.H.H., G.S.E.-S., A.M.A., S.E.G., O.M.A.z., A.M.S. and A.M.A.K.; validation, M.S.A., A.H.H., G.S.E.-S., A.M.A., S.E.G., O.M.A.z., A.M.S. and A.M.A.K.; formal analysis, M.S.A., A.H.H., G.S.E.-S., A.M.A., S.E.G., O.M.A.z., A.M.S. and A.M.A.K.; investigation, M.S.A., A.H.H., G.S.E.-S., A.M.A., S.E.G., O.M.A.z., A.M.S. and A.M.A.K.; resources, M.S.A., A.H.H., G.S.E.-S., A.M.A., S.E.G., O.M.A.z., A.M.S. and A.M.A.K.; data curation, M.S.A., A.H.H., G.S.E.-S., A.M.A., S.E.G., O.M.A.z., A.M.S. and A.M.A.K.; writing—original draft preparation, M.S.A., A.H.H., G.S.E.-S., A.M.A., S.E.G., O.M.A.z., A.M.S. and A.M.A.K.; writing—review and editing, M.S.A., A.H.H., G.S.E.-S., A.M.A., S.E.G., O.M.A.z., A.M.S. and A.M.A.K.; project administration, A.M.S.; funding acquisition, O.M.A.z., A.M.A.K. and A.M.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Deputyship for Research & Innovation, Ministry of Education in Saudi Arabia [project number 445-9-560].
Data Availability Statement
The data generated and analyzed during this study are included in this article.
Acknowledgments
The authors extend their appreciation to the Deputyship for Research & Innovation, Ministry of Education in Saudi Arabia for funding this research work through the project number 445-9-560.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Russell, G.E. Plant Breeding for Pest and Disease Resistance: Studies in The Agricultural and Food Sciences; Elsevier Ltd.: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Djébali, N.; Elkahoui, S.; Taamalli, W.; Hessini, K.; Tarhouni, B.; Mrabet, M. Tunisian Rhizoctonia solani AG3 strains affect potato shoot macronutrients content, infect faba bean plants and show in vitro resistance to azoxystrobin. Australas. Plant Pathol. 2014, 43, 347–358. [Google Scholar] [CrossRef]
- Abebe, T.; Birhane, T.; Nega, Y.; Workineh, A. The prevalence and importance of faba bean diseases with special consideration to the newly emerging faba bean gall in Tigray, Ethiopia. Discourse J. Agric. Food Sci. 2014, 2, 33–38. [Google Scholar]
- Rosenzweig, C.; Iglesius, A.; Yang, X.-B.; Epstein, P.R.; Chivian, E. Climate Change and Extreme Weather Events-Implications for Food Production, Plant Diseases, and Pests; NASA: Washington, DC, USA, 2001. [Google Scholar]
- Alhakim, A.A.; Hashem, A.; Abdelaziz, A.M.; Attia, M.S. Impact of plant growth promoting fungi on biochemical defense performance of Tomato under Fusarial infection. Egypt. J. Chem. 2022, 8, 775. [Google Scholar]
- Hashem, A.H.; Abdelaziz, A.M.; Askar, A.A.; Fouda, H.M.; Khalil, A.M.; Abd-Elsalam, K.A.; Khaleil, M.M. Bacillus megaterium-mediated synthesis of selenium nanoparticles and their antifungal activity against Rhizoctonia solani in faba bean plants. J. Fungi 2021, 7, 195. [Google Scholar] [CrossRef] [PubMed]
- Hasanin, M.; Hashem, A.H.; Lashin, I.; Hassan, S.A. In vitro improvement and rooting of banana plantlets using antifungal nanocomposite based on myco-synthesized copper oxide nanoparticles and starch. Biomass Convers. Biorefinery 2023, 13, 8865–8875. [Google Scholar] [CrossRef]
- Abdelaziz, A.M.; Salem, S.S.; Khalil, A.M.; El-Wakil, D.A.; Fouda, H.M.; Hashem, A.H. Potential of biosynthesized zinc oxide nanoparticles to control Fusarium wilt disease in eggplant (Solanum melongena) and promote plant growth. BioMetals 2022, 35, 601–616. [Google Scholar] [CrossRef] [PubMed]
- Vallad, G.E.; Goodman, R.M. Systemic acquired resistance and induced systemic resistance in conventional agriculture. Crop Sci. 2004, 44, 1920–1934. [Google Scholar] [CrossRef]
- Gong, D.; Celi, N.; Zhang, D.; Cai, J. Magnetic Biohybrid Microrobot Multimers Based on Chlorella Cells for Enhanced Targeted Drug Delivery. ACS Appl. Mater. Interfaces 2022, 14, 6320–6330. [Google Scholar] [CrossRef]
- Gong, D.; Sun, L.; Li, X.; Zhang, W.; Zhang, D.; Cai, J. Micro/Nanofabrication, Assembly, and Actuation Based on Microorganisms: Recent Advances and Perspectives. Small Struct. 2023, 4, 2200356. [Google Scholar] [CrossRef]
- Elmer, W.H.; Zuverza-Mena, N.; Triplett, L.R.; Roberts, E.L.; Silady, R.A.; White, J.C. Foliar application of copper oxide nanoparticles suppresses fusarium wilt development on chrysanthemum. Environ. Sci. Technol. 2021, 55, 10805–10810. [Google Scholar] [CrossRef]
- Ashraf, H.; Anjum, T.; Riaz, S.; Ahmad, I.S.; Irudayaraj, J.; Javed, S.; Qaiser, U.; Naseem, S. Inhibition mechanism of green-synthesized copper oxide nanoparticles from Cassia fistula towards Fusarium oxysporum by boosting growth and defense response in tomatoes. Environ. Sci. Nano 2021, 8, 1729–1748. [Google Scholar] [CrossRef]
- Gaber, S.E.; Hashem, A.H.; El-Sayyad, G.S.; Attia, M.S. Antifungal activity of myco-synthesized bimetallic ZnO-CuO nanoparticles against fungal plant pathogen Fusarium oxysporum. Biomass Convers. Biorefinery 2023, 13, 13729–14299. [Google Scholar] [CrossRef]
- Attia, M.S.; Salem, M.S.; Abdelaziz, A.M. Endophytic fungi Aspergillus spp. reduce fusarial wilt disease severity, enhance growth, metabolism and stimulate the plant defense system in pepper plants. Biomass Convers. Biorefinery 2022, 1–11. [Google Scholar] [CrossRef]
- Vernon, L.P.; Seely, G.R. The Chlorophylls; Academic Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Classics Lowry, O.; Rosebrough, N.; Farr, A.; Randall, R. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Umbreit, W.W.; Burris, R.H.; Stauffer, J.F. Manometric Techniques: A Manual Describing Methods Applicable to the Study of Tissue Metabolism; Burgess: Clayton, NC, USA, 1964. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Dai, G.; Andary, C.; Cosson-Mondolot, L.; Boubals, D. Polyphenols and resistance of grapevines to downy mildew. In Proceedings of International Symposium on Natural Phenols in Plant Resistance 381; ISHS: Brugge, Belgium, 1993; pp. 763–766. [Google Scholar]
- Hu, Z.; Richter, H.; Sparovek, G.; Schnug, E. Physiological and biochemical effects of rare earth elements on plants and their agricultural significance: A review. J. Plant Nutr. 2004, 27, 183–220. [Google Scholar] [CrossRef]
- Mukherjee, S.; Choudhuri, M. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Srivastava, S. Peroxidase and poly-phenol oxidase in Brassica juncea plants infected with Macrophomina phaseolina (Tassai) Goid. and their implication in disease resistance. J. Phytopathol. 1987, 120, 249–254. [Google Scholar]
- Matta, A. Accumulation of phenols in tomato plants infected by different forms of Fusarium oxysporum. Phytopathology 1969, 59, 512–513. [Google Scholar]
- Ma, C.; Borgatta, J.; De La Torre-Roche, R.; Zuverza-Mena, N.; White, J.C.; Hamers, R.J.; Elmer, W.H. Time-dependent transcriptional response of tomato (Solanum lycopersicum L.) to Cu nanoparticle exposure upon infection with Fusarium oxysporum f. sp. lycopersici. ACS Sustain. Chem. Eng. 2019, 7, 10064–10074. [Google Scholar]
- Shah, I.H.; Ashraf, M.; Sabir, I.A.; Manzoor, M.A.; Malik, M.S.; Gulzar, S.; Ashraf, F.; Iqbal, J.; Niu, Q.; Zhang, Y. Green synthesis and Characterization of Copper oxide nanoparticles using Calotropis procera leaf extract and their different biological potentials. J. Mol. Struct. 2022, 1259, 132696. [Google Scholar] [CrossRef]
- Ali, M.A.; Ahmed, T.; Wu, W.; Hossain, A.; Hafeez, R.; Islam Masum, M.M.; Wang, Y.; An, Q.; Sun, G.; Li, B. Advancements in plant and microbe-based synthesis of metallic nanoparticles and their antimicrobial activity against plant pathogens. Nanomaterials 2020, 10, 1146. [Google Scholar] [CrossRef]
- Elsakhawy, T.; ALKahtani, M.D.; Sharshar, A.A.; Attia, K.A.; Hafez, Y.M.; Abdelaal, K.A. Efficacy of mushroom metabolites (Pleurotus ostreatus) as a natural product for the suppression of broomrape growth (Orobanche crenata Forsk) in faba bean plants. Plants 2020, 9, 1265. [Google Scholar] [CrossRef]
- Prakash, O. Diseases and disorders of mango and their management. In Diseases of Fruits and Vegetables Volume I: Diagnosis and Management; Springer: Berlin/Heidelberg, Germany, 2004; pp. 511–619. [Google Scholar]
- Ragab, S.M.; Turoop, L.; Runo, S.; Nyanjom, S. The effect of foliar application of zinc oxide nanoparticles and Moringa oleifera leaf extract on growth, biochemical parameters and in promoting salt stress tolerance in faba bean. Afr. J. Biotechnol. 2022, 21, 252–266. [Google Scholar]
- García-López, J.I.; Niño-Medina, G.; Olivares-Sáenz, E.; Lira-Saldivar, R.H.; Barriga-Castro, E.D.; Vázquez-Alvarado, R.; Rodríguez-Salinas, P.A.; Zavala-García, F. Foliar application of zinc oxide nanoparticles and zinc sulfate boosts the content of bioactive compounds in habanero peppers. Plants 2019, 8, 254. [Google Scholar] [CrossRef] [PubMed]
- Avellan, A.; Yun, J.; Morais, B.P.; Clement, E.T.; Rodrigues, S.M.; Lowry, G.V. Critical review: Role of inorganic nanoparticle properties on their foliar uptake and in planta translocation. Environ. Sci. Technol. 2021, 55, 13417–13431. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-N.; Wu, L.-T.; Kun, S.; Zhu, Y.-S.; Wei, D. Nonphytotoxic copper oxide nanoparticles are powerful “nanoweapons” that trigger resistance in tobacco against the soil-borne fungal pathogen Phytophthora nicotianae. J. Integr. Agric. 2022, 21, 3245–3262. [Google Scholar] [CrossRef]
- de Sousa, T.P.; Chaibub, A.A.; da Silva, G.B.; de Filippi, M.C.C. Trichoderma asperellum modulates defense genes and potentiates gas exchanges in upland rice plants. Physiol. Mol. Plant Pathol. 2020, 112, 101561. [Google Scholar] [CrossRef]
- Murali, M.; Anandan, S.; Ansari, M.A.; Alzohairy, M.A.; Alomary, M.N.; Asiri, S.M.M.; Almatroudi, A.; Thriveni, M.; Singh, S.B.; Gowtham, H.G. Genotoxic and cytotoxic properties of zinc oxide nanoparticles phyto-fabricated from the obscure morning glory plant Ipomoea obscura (L.) Ker Gawl. Molecules 2021, 26, 891. [Google Scholar]
- Patel, P.; Kansara, K.; Senapati, V.A.; Shanker, R.; Dhawan, A.; Kumar, A. Cell cycle dependent cellular uptake of zinc oxide nanoparticles in human epidermal cells. Mutagenesis 2016, 31, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, P.K.; Singh, A.K.; Singh, V.P.; Prasad, S.M.; Ramawat, N.; Tripathi, D.K.; Chauhan, D.K.; Rai, A.K. Liquid assisted pulsed laser ablation synthesized copper oxide nanoparticles (CuO-NPs) and their differential impact on rice seedlings. Ecotoxicol. Environ. Saf. 2019, 176, 321–329. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1. [Google Scholar]
- Yruela, I. Copper in plants: Acquisition, transport and interactions. Funct. Plant Biol. 2009, 36, 409–430. [Google Scholar] [CrossRef]
- Hänsch, R.; Mendel, R.R. Physiological functions of mineral micronutrients (cu, Zn, Mn, Fe, Ni, Mo, B, cl). Curr. Opin. Plant Biol. 2009, 12, 259–266. [Google Scholar] [CrossRef]
- Ubbink, M.; Ejdebäck, M.; Karlsson, B.G.; Bendall, D.S. The structure of the complex of plastocyanin and cytochrome f, determined by paramagnetic NMR and restrained rigid-body molecular dynamics. Structure 1998, 6, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Attia, M.S.; Younis, A.M.; Ahmed, A.F.; Elaziz, A. Comprehensive management for wilt disease caused by Fusarium oxysporum in tomato plant. Int. J. Innov. Sci. Eng. Technol. 2016, 4, 2348–7968. [Google Scholar]
- Pandey, S.; Giri, K.; Kumar, R.; Mishra, G.; Raja Rishi, R. Nanopesticides: Opportunities in crop protection and associated environmental risks. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2018, 88, 1287–1308. [Google Scholar] [CrossRef]
- Al-Doori, S.A.M.A. Response of Three Flax Genotypes (Linum usitatissimum L.) to Foliar Spraying with Different Concentration of Zinc and Boron under the Dryland Conditions of Nineveh Governorate. Coll. Basic Educ. Res. J. 2021, 17, 1680–1700. [Google Scholar]
- Abdelaziz, A.M.; Elshaer, M.A.; Abd-Elraheem, M.A.; Ali, O.M.O.M.; Haggag, M.I.; El-Sayyad, G.S.; Attia, M.S. Ziziphus spina-christi extract-stabilized novel silver nanoparticle synthesis for combating Fusarium oxysporum-causing pepper wilt disease: In vitro and in vivo studies. Arch. Microbiol. 2023, 205, 69. [Google Scholar]
- Banerjee, A.; Mittra, B.; Das, A.B. Biochemical and histological characterisation of Fusarium oxysporum infected wheat (Triticum aestivum) seedlings in vitro. Indian Phytopathol. 2022, 75, 559–563. [Google Scholar] [CrossRef]
- El-Batal, A.I.; El-Sayyad, G.S.; Al-Shammari, B.M.; Abdelaziz, A.M.; Nofel, M.M.; Gobara, M.; Elkhatib, W.F.; Eid, N.A.; Salem, M.S.; Attia, M.S. Protective role of iron oxide nanocomposites on disease index, and biochemical resistance indicators against Fusarium oxysporum induced-cucumber wilt disease: In vitro, and in vivo studies. Microb. Pathog. 2023, 180, 106131. [Google Scholar]
- McCord, J.M. The evolution of free radicals and oxidative stress. Am. J. Med. 2000, 108, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Abdelaziz, A.M.; El-Wakil, D.A.; Hashem, A.H.; Al-Askar, A.A.; AbdElgawad, H.; Attia, M.S. Efficient Role of Endophytic Aspergillus terreus in Biocontrol of Rhizoctonia solani Causing Damping-off Disease of Phaseolus vulgaris and Vicia faba. Microorganisms 2023, 11, 1487. [Google Scholar]
- Roy, D.; Adhikari, S.; Adhikari, A.; Ghosh, S.; Azahar, I.; Basuli, D.; Hossain, Z. Impact of CuO nanoparticles on maize: Comparison with CuO bulk particles with special reference to oxidative stress damages and antioxidant defense status. Chemosphere 2022, 287, 131911. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; He, Z.; Chen, N.; Tang, Z.; Wang, Q.; Cai, Y. The roles of environmental factors in regulation of oxidative stress in plant. BioMed Res. Int. 2019, 2019, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Attia, M.S.; Abdelaziz, A.M.; Hassanin, M.M.; al-askar, A.A.; Marey, S.A.; Abdelgawad, H.; Hashem, A.H. Eco-friendly preparation of thyme essential oil nano emulsion: Characterization, antifungal activity and resistance of Fusarium wilt disease of Foeniculum vulgare. Not. Bot. Horti Agrobot. Cluj-Napoca 2023, 51, 13312. [Google Scholar]
- Daigham, G.E.; Mahfouz, A.Y.; Abdelaziz, A.M.; Nofel, M.M.; Attia, M.S. Protective role of plant growth-promoting fungi Aspergillus chevalieri OP593083 and Aspergillus egyptiacus OP593080 as biocontrol approach against Alternaria leaf spot disease of Vicia faba plant. Biomass Convers. Biorefinery 2023, 1–17. [Google Scholar]
- Abou El Nour, M.M.; Mona, E.; Wadi, J. Suppressive effect of compost/pomegranate peel tea combination against Fusarium oxysporum f. sp. lupini, and Rhizoctonia solani as an alternative synthetic fungicide. Egypt. J. Exp. Biol. 2020, 16, 13–25. [Google Scholar]
- Thapa, S.; Bhandari, A.; Ghimire, R.; Xue, Q.; Kidwaro, F.; Ghatrehsamani, S.; Maharjan, B.; Goodwin, M. Managing micronutrients for improving soil fertility, health, and soybean yield. Sustainability 2021, 13, 11766. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).