Abstract
Supplementary irrigation can be strategic for forage cactus, aiming to guarantee higher levels of productivity and economic return. Therefore, this study aimed to evaluate the growth dynamics, productivity, evapotranspiration, and water economy indices of three forage cactus clones under different water regimes. The study was carried out in the Brazilian semi-arid region during two cycles between 2020 and 2022. Clones of forage cactus Orelha de Elefante Mexicana—OEM (Opuntia stricta (Haw.) Haw.) were evaluated; IPA Sertânia—IPA and Miúda—MIU (both species Nopalea cochenillifera (L.) Salm-Dyck) were subjected to four water regimes: rainfed and three irrigation depths (4, 8 and 12 mm). Growth dynamics and evapotranspiration were evaluated throughout the cycles, and productivity and water economy indices were based on data from the harvest time. The OEM clone showed higher dry matter productivity (21.8 Mg ha−1). The MIU clone, when intended for seed sales, presented a higher economic return (USD 20,823.33). The use of irrigation brought forward the cutting time and reduced water saving rates. With this, it can be concluded that the increase in water availability in cactus increases water consumption and reduces economic-water indices but brings important productive returns.
1. Introduction
In semi-arid environments, it is crucial to increase forage production sustainably. However, soil (e.g., low fertility, shallow soils, low water retention and salinity) and climate (e.g., high temperatures, high atmospheric demand, and drought) conditions have significant negative impacts on production [1,2,3]. Additionally, climate risk areas have been increasing in recent years [4,5]. To minimize production losses caused especially by the irregular distribution of rainfall and water scarcity, it is necessary to adopt resilience practices, for example, the use of species adapted to the local climate and soil [6,7,8] and supplementary irrigation in dry periods of the year [9,10,11].
In this context, forage cactus can be a strategic species to form resilient production systems, as it has the crassulacean acid metabolism (CAM) (e.g., stomatal opening at night and modified leaves), which provides a high capacity to withstand long periods of drought [12,13,14,15]. Furthermore, it has great potential as a source of bulk food for livestock, as the crop is an energy source for ruminants [16].
The use of varieties that stand out in productive and qualitative terms and that are resistant to Carmine cochineal is essential in the Brazilian semi-arid region, where clones of the genus Nopalea (IPA Sertânia—IPA and Miúda—MIU) and the genus Opuntia (Orelha de Elefante Mexicana—OEM) stand out [12]. However, the lack of information regarding the management of this crop compromises agricultural planning and decision-making on the property. As a result, there is a reduction in forage input and, consequently, instability in animal production.
Water management via irrigation, as well as meteorological conditions in cactus cultivation, alters its growth and productivity dynamics [17]. For example, the OEM clone, subjected to controlled water deficit, shows greater accumulation of dry matter (DM) compared to the MIU and IPA clones [18]. The clones of the genus Nopalea (IPA and MIU), on the other hand, showed a positive effect on the emission of cladodes when irrigated in water regimes of 80 and 120% of crop evapotranspiration (ETc); in addition, there was an anticipation of the moment of cutting of the three clones when irrigation was applied. Despite this, the water supply in cultivation systems with forage cactus can reduce the capacity to convert water into DM [19,20].
Economic information combined with water use efficiency indicators in crops can help develop strategies and tools to improve water and monetary management (i.e., better allocation of the farmer’s financial resources) of properties in semi-arid regions [21,22,23]. Furthermore, the morphophysiological indices (absolute growth rate—AGR, relative growth rate—RGR, net assimilation rate—NAR, specific cladode area—SCA, and cladode area index rate—CAIR) can be used in a simple and exploratory way to verify crop performance in different water availability conditions and even improve water use in the production system [18,24]. Studies with morphophysiological indices in forage cactus have been developed in recent years [7,18,24]. For example, in semi-arid conditions, ref. [18] found that a controlled reduction in the amount of water applied did not reduce the growth dynamics of OEM, MIU and IPA clones. However, these authors only evaluated one production cycle. Considering that the responses of this crop may vary depending on the growing season [7,18], there is a need to understand the morphological responses that contribute to the production of forage cacti when subjected to different water regimes at different seasons.
This study hypothesizes that supplementary irrigation positively affects the production and growth dynamics, water-economic indices, and the water balance of forage cactus. With this, the objective was to evaluate the effects of different irrigation levels on the growth and production dynamics, water-economic indices, and soil hydrodynamics of three forage cactus clones resistant to Carmine cochineal in the Brazilian semi-arid region.
2. Materials and Methods
2.1. Study Area
The experimental field is located at the International Reference Center for Agrometeorological Studies of Cactus and Other Forage Plants—REF Center, established at the Federal Rural University of Pernambuco—Academic Unit of Serra Talhada (UFRPE/UAST), municipality of Serra Talhada—PE, Brazil (latitude 7°56′20″ south, longitude 38°17′31″ west and altitude 499 m). The local climate was classified as BShw’ (hot semi-arid, with dry winter and rainy summer) according to the Köppen climate classification [25,26], with an average air temperature of 26.6 °C, and minimum and maximum of 20.1 to 32.9 °C, respectively, average rainfall of 642.1 mm year−1, average relative humidity of 62.5% and atmospheric demand above 1800 mm year−1 [27]. Historically, the region is marked by the cultivation of subsistence crops (e.g., maize and beans), as well as the cultivation of forage plants (e.g., cactus forage, sorghum, and grass, among others) to feed local livestock.
The soil in the experimental area was classified as typical Eutrophic Haplic Ta Cambisol [28], with a sandy texture, not stony, having a flat topography and being well-drained, with a medium soil density of 1.38 g cm−3, field capacity of 0.11 kg kg−1 and permanent wilting point of 0.05 kg kg−1 (Table 1).

Table 1.
Physical and chemical properties of typical Eutrophic Haplic Ta Cambisol at depths of 0–20 and 20–40 cm in Serra Talhada, PE, Brazil.
The trial was conducted in a randomized block design, in a 4 × 3 factorial arrangement, with subdivided plots, the plots being represented by rainfed conditions and three irrigation depths—IDs (4, 8, and 12 mm). In turn, the subplots were made up of three forage cactus clones: Orelha de Elefante Mexicana—OEM (Opuntia stricta (Haw.) Haw.); IPA Sertânia—IPA; and Miúda—MIU (both species Nopalea cochenillifera (L.) Salm-Dyck), in a density of 50,000 plants ha−1, with a spacing of 1.0 m between rows and 0.2 m between plants (1.0 × 0.2 m).
The culture was implemented in January 2016, when the first and second production cycle was conducted until November 2020. After this period, the evaluation periods for the current study began, where the first experimental cycle lasted 433 days and the second experimental cycle lasted 362 days.
The IDs (4, 8, and 12 mm) were applied weekly, being carried out only in the dry period, through a drip system, with emitters spaced 0.20 m and a flow rate of 1.87 L h−1, at a pressure of 1 atm. The IDs were based on crop evapotranspiration fractions (controlled water deficit—40%, controlled water deficit—80%, and full irrigation greater than the crop demand—120% of ETc [18]), considering a Kc of 0.52 and the average reference evapotranspiration (ET0) equal to 5 mm day−1. The water used, coming from an artesian well, was classified as C3S1 (i.e., water with high salinity and low sodium content) [29] and presented an average electrical conductivity of 1.51 dS m−1 and pH of 6.84. To ensure that irrigation could be set at weekly intervals, rainfall (R) was monitored daily. In this way, irrigation was carried out in weeks in which the R was lower than the imposed water depths, discounting the total precipitation of that week. On the other hand, when the total R was greater than the ID, there was no irrigation.
Meteorological variables were monitored (Figure 1) from a data collection platform located 10 m from the experimental area, belonging to the National Institute of Meteorology. The data comprise hourly measurements of average, maximum, and minimum relative air humidity (% day−1); maximum, minimum, and average air temperature (°C day−1); wind speed (m s−1 day−1); global solar radiation (MJ m−2 day−1) and rainfall (mm day−1). Therefore, it was used as a basis for calculating ET0, estimated by the Penman-Monteith equation parameterized in the FAO-56 bulletin [30].
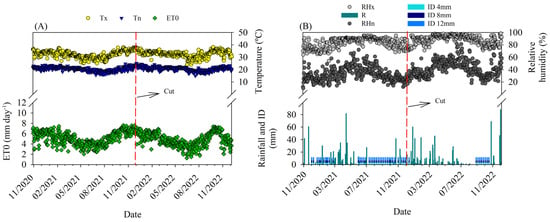
Figure 1.
(A) Maximum (Tx), minimum (Tn) temperatures and reference evapotranspiration (ET0); (B) Maximum (RHx), minimum (RHn) relative humidity, irrigation depths (ID 4 mm; ID 8 mm and ID 12 mm) and rainfall (R) in the period from September 2020 to December 2022, during two successive cactus crop cycles.
In Figure 1A, during the first cactus production cycle, the maximum and minimum air temperatures observed during this period were 37.9 and 14.9 °C, respectively, with the daily average being 25.7 °C. In the second cycle, the maximum and minimum air temperatures observed were 38.1 and 15.6 °C, respectively, with the daily average temperature being 25.1 °C. The vertical dashed line in Figure 1A,B corresponds to the harvest date of the first experimental cycle (5 January 2022), starting the subsequent cycle (5 January–5 December 2022). Throughout the experimental period, the accumulated rainfall was 1647 mm in total, 928.2 mm in the first experimental cycle, and 719.1 mm in the second cycle. The water volumes applied via irrigation in the first season were 131, 262, and 393 mm for the 4, 8, and 12 mm depths, respectively, and in the second cycle 30, 60, and 90 mm, respectively. It was possible to observe the highest RHx values during rainy periods, reaching 100%, on the other hand, the lowest RHn values were presented in the period with the lowest rainfall volume, reaching 10% (Figure 1B). On average, the RH was equal to 61.19%. In general, during the two periods studied, the average air temperature and relative humidity presented values similar to those observed historically in the region (average temperature = 26.6 °C and RH = 62.5) [27]. On the other hand, rainfall was 44 and 12% higher in the first and second cycles, respectively, about the normal observed at the study site (642.1 mm year−1) [27].
2.2. Monitoring Soil Water Dynamics
The soil water balance of the experiment was carried out according to [31], in which the volume of soil was considered depending on the root system of the crop under study (0.4 m depth). Therefore, the upper limit of this volume is the soil surface, and the lower limit is the depth of the crop’s root system.
where ET is actual evapotranspiration (mm); R is rainfall (mm); ID is the irrigation depth (mm); SR is soil runoff (mm); CR is the capillary rise (mm), and Δs is the variation in water storage in the soil (mm).
We monitored soil moisture during the experiment, where 36 PVC (polyvinyl chloride) access pipes were installed and inserted at a depth of 0.70 m from the soil surface. Readings were taken with the probe three times a week (i.e., Mondays, Wednesdays, and Fridays).
2.3. Growth, Yield, and Morpho-Physiological Indices
With the aid of a tape measure and caliper, biometrics were carried out at the end of each cycle to measure the morphological characteristics of the plant: plant height (PH), plant width (PW), the total number of cladodes (TNC), number of first-, second-, third- and fourth-order cladodes (NC1, NC2, NC3 and NC4, respectively), and some in the cladode: width (CW), length (CL), perimeter (CP) and thickness (CT) of the cladodes.
With these data, the cladode area (CA) in order (Equations (2)–(4)) and the cladode area index (CAI, Equation (5)) were estimated for each clone according to the methodology proposed by [32]:
where CL: cladode length (cm); CW: cladode width (cm); CP: cladode perimeter; 10,000: the conversion factor from cm2 to m2. S1 and S2 is the spacing between the lines and cactus plants (1 × 0.2 m).
Biomass events were carried out at the end of each cycle to obtain cactus productivity. With this, the total fresh mass (TFM) was obtained, in which the plant was weighed, and a representative cladode of each order was chosen for weighing on a semi-analytical balance. Then, the cladodes were fractionated and conditioned in a forced air circulation oven at a temperature of 55 °C, until reaching constant dry weight, making it possible to obtain dry mass (DM) and dry mass content (DMC).
The morpho-physiological indices were obtained by regression analysis, in which three-parameter sigmoid models (Equation (6)) were applied between the DM and CAI values and the thermal accumulation of the crop (accumulated degree days, ADD), which in turn was obtained as the sum of the difference between the daily values of the average air temperature (°C) and 22 °C, which is the lower base temperature of the culture (°C) [18].
Sigmoid models that presented a high coefficient of determination (R2 ≥ 0.85) were derived (Equation (7)) to calculate the daily dry matter accumulation rate, resulting in the absolute crop growth rate (AGR, Mg ha−1 °Cday−1). In addition to AGR, the net assimilation rate (NAR, Mg ha−1 °Cday−1), the relative growth rate (RGR, Mg Mg−1 °Cday−1), and the specific cladode area rate (SCA, ha Mg−1) calculating the ratios, NAR = AGR/CAI, RGR = AGR/DM and SCA = CAI/DM. To calculate the cladode area index rate (CAIR), the sigmoid model of the CAI was derived (Equation (7)).
where Y is the variable estimated at time x (ADD); a is the maximum value for the fee; b is the number of degree days needed to initiate the rate; and x0 is the time (ADD) necessary for the plant to reach 50% of the rate.
With the three-parameter sigmoid models (Equation (6)), the phenophases were determined by regression analysis to assess the interaction throughout the experimental period between the number of cladodes per order and the ADD. Sigmoid models (R2 ≥ 0.85) were derived to determine the rate of cladode production by order (Equation (7)). According to [33], the beginning of a new vegetative phase is when the emission rate of cladodes of a given order is lower than the emission rate of the subsequent order. When 25% of the peak of the culture’s DM accumulation rate was obtained, the cutting time was determined [18]. The adjustment of the regression curves was carried out in Excel 2019, with the help of the mathematical software Solver. The graphs were prepared using SigmaPlot (v14.5).
2.4. Water and Economic Indicators
Good agricultural management on properties associated with the use of economic indicators and water use is important for the socioeconomic development of the population in semi-arid environments. To understand the response of forage cactus subjected to different levels of irrigation, this study applied nine indices proposed by [22].
where WUEc is the relationship between the crop’s real evapotranspiration (ET) and the total water applied by irrigation (ID) and rainfall (R); Unit: m3 ha−1/m3 ha−1.
where *WUEc is the relationship between the biomass produced by the crop and the crop’s evapotranspiration (ET) throughout the cycle; Unit: kg ha−1/m3 ha−1.
where WPc is the crop water productivity, being a relationship between the yield produced and the water consumed by the crop (ET); Unit: kg ha−1/m3 ha−1.
*WPc is the crop water productivity, being the relationship between dry mass (DM) and the total water involved in cultivation (TWU), which in turn, is the sum of ET and the non-beneficial use of water (percolation and flow); Unit: kg ha−1/m3 ha−1.
where WPI is the productivity of water irrigation, which is the ratio between dry matter yield and water applied via irrigation in the crop cycle; Unit: kg ha−1/m3 ha−1.
where GEWPI is the gross economic productivity of irrigation water, being the relationship between the gross margin (Gross Margin = Revenues Variable costs) and the irrigation water applied; Unit: USD ha−1/m3 ha−1.
where NEWPI is the net economic productivity of irrigation water, being the relationship between net margin (Net Margin = Revenue − Fixed and variable costs) and irrigation water applied to the crop; Unit: USD ha−1/m3 ha−1.
where EWPc is the economic productivity of crop water, being the ratio between Profit (Opportunity cost profit) and total water involved in cultivation (TWU); Unit: USD ha−1/m3 ha−1.
where EWPI is the economic productivity of irrigation water, being the ratio between revenue and applied irrigation water (IWU); Unit: USD ha−1 m−3 ha−1. To quantify revenue and opportunity cost profit, the value of fresh mass was adopted to equal 28.79 USD per tonn and the value of the cladode unit for planting equal 0.03 USD, respectively. The opportunity cost profit is the value of one cladode unit multiplied by the number of middle third cladodes produced per plant. The costs of installing the irrigation system and implementing the crop were considered fixed costs, while the variable costs comprised the sum of energy costs, labor (maintenance) and the depreciation of the irrigation system components.
2.5. Statistical Analysis
The data were subjected to normality (Shapiro–Wilk) and homoscedasticity (Bartlett) tests. Considering the premises, analysis of variance (ANOVA) of the data was carried out, using the F test, at a 5% probability level. When significant, Tukey’s test was performed for qualitative factors and regression for quantitative factors. All analyses were carried out using the computational tool RStudio [34].
3. Results
3.1. Water Balance
Table 2 shows the hydrodynamic components of the soil, cultivated with three forage cactus clones (IPA, MIU, and OEM) and subjected to four levels of irrigation (0, 4, 8, and 12 mm weekly). Each cultivation system was divided into five periods, in which in cycles one and two, the first four intervals lasted 77 days; on the other hand, the fifth period lasted 91 days, and the second lasted 21 days. Therefore, the duration of the first cycle was 399 days and the second cycle was 329 days.
Table 2.
Sequential water balance in cactus cultivation subjected to different levels of mulch in the Brazilian semi-arid region.
The forms of water input into cultivation are given by the sum of rainfall and the volume of water input via irrigation (R + ID); when cultivated in rainfed conditions, it rained a total of 816 mm in the first cycle and 607 mm in the subsequent period. With the increase in water via irrigation in the first cycle, a total of 936, 1056, and 1177 mm was accumulated at depths of 4, 8, and 12 mm, respectively. In the following season, water input to cultivation accumulated a total volume of 647, 687, and 727 mm, in the same order (Table 2).
The variation in water storage in the soil (Δs), in turn, showed fluctuation in both cycles according to the cactus clone and water availability of the crop (water availability via irrigation and rainfall). In general terms, the OEM, MIU, and IPA clones, respectively expressed mean Δs values of −2, −1, and 3 mm in the first cycle and −4, −6, and −9 mm in the second.
When cultivated under rainfed conditions, the IPA clone had a Δs of −5 mm in cycle one and −11 mm in cycle two, the MIU clone demonstrated magnitudes of −4 and −9 mm and the OEM clone expressed values of 3 and −16 mm, in the same order. Inputs of 4, 8, and 12 mm weekly via irrigation had Δs of 3, −1, and −4 mm in the first season and −9, 3, and 2 mm in the second cultivation cycle of the IPA clone. In the MIU clone, there was an effect of irrigation depths on Δs, in which the weekly supply of 4, 8, and 12 mm, respectively, obtained Δs of 1, 2, and −2 mm. In turn, the water variation in the soil of the OEM clone was 3, 2, and 2 mm in the first cycle (for the depths of 4, 8, and 12 mm, respectively). In the second cycle, this clone expressed magnitudes of −16, −3, and −2 mm, respectively, for the 4, 8 and 12 mm depths.
With the increase in R + ID, there was a positive effect on ET in all evaluated clones, where the maximum value was found at the 12 mm depth and the minimum in cultivation without irrigation; in addition, this behavior was repeated for ETd. For the IPA clone, the maximum ET was 903 mm in the first cycle and 580 mm in the second. In the MIU clone, the maximum magnitude was 901 and 584 mm. In turn, the highest value of the OEM clone was 897 and 585 mm in cycles one and two, respectively. The lower values of the IPA, MIU, and OEM clones were, respectively, 595, 594, and 587 mm in the initial cycle and 481, 479, and 486 mm in the later cycle.
3.2. Structural Characteristics of Forage Cactus
The average productivity values in fresh matter (FMY), dry matter (DMY) and dry matter content (DMC) of the cactus clones cultivated in Experiment I are presented in Table 2. For FMY and DMY, the OEM clone exhibited higher values in comparison to the others (p < 0.05), with an average of 182.51 Mg ha−1 and 19.26 Mg ha−1, respectively. The MIU and IPA clones did not differ from each other (p > 0.05), presenting an average yield equal to 93.54 Mg ha−1 in FMY and 9.83 Mg ha−1 in DMY. An average DMC of 0.10 was found for the clones (p > 0.05).
In the first experimental cycle, plant height (PH) and the number of fourth-order cladodes (NC4) showed a significant effect (p < 0.05) on the interaction between clones (IPA, MIU, and OEM) and irrigation depths (0, 4, 8, and 12 mm); however, when evaluating isolated factors, it was possible to observe a significant difference in some specific variables, such as biometric variables—except for the number of second- and fifth-order cladodes (NC2 and NC5, respectively) and plant width (PW), for the clone factor—and some cladode variables, such as the length (CL), width (CW), and perimeter (CP) of the cladode (Table S1).
In the subsequent cycle, there was an interaction between clone factors and irrigation depths for the variables total number of cladodes (TNC) and number of fourth-order cladodes (NC4) (p < 0.05). As for the isolated factors, it was observed that the clone presented significance for the numbers of cladodes (TNC, NC1, NC3, and NC4) and the biometric variables of the cladodes (CL, CW, CT, CP, and CAI). The depth factor was significant in the variables NC4, CL, CT, and CP (Table S1).
The mean PH of IPA, MIU, and OEM clones in the first cycle was 84.7, 80.02, and 91.87 cm, respectively. The OEM clone irrigated with 12 mm weekly showed greater height, differing statistically (p < 0.05). The mean PW was 72.4, 65.8 and 69.8 cm for the IPA, MIU, and OEM clones, respectively; however, only the 8 mm depth demonstrated a difference between the evaluated clones, with the IPA and OEM clone expressing greater plant widths (78.7 and 66.8 cm) (p < 0.05). At the 12 mm depths, the MIU clone showed higher NC compared to the other clones irrigated with the same depths. On the other hand, there was no difference in the other treatments (Table 3).
Table 3.
Biometric variables of three forage cactus clones subjected to different levels of irrigation in the Brazilian semi-arid region.
In general, in the first cycle, the mean number of first-order cladodes (NC1) for the IPA, MIU, and OEM clones was 4.5, 3.3 and 5.2 units, respectively. However, there was no significant effect on the depth; when comparing the clones in rainfed cultivation, a better performance was noted for OEM and IPA (5.2 and 5.0 units, respectively). Despite this, when NC3 was observed, the MIU clone stood out in all water conditions (p < 0.05). Regarding NC2, NC4, and NC5, no significant difference was noted for the clones and irrigation depths (p > 0.05) (Table 3).
The biometric variables of the cladodes in the first cycle showed a significant effect for the clones, except CT, which differed only in rainfed cultivation, where the IPA clone presented greater thickness (33.0 mm) than the MIU and OEM clones (24.9 and 22.5 mm). The OEM and IPA clones demonstrated greater magnitudes for CL in all depths; however, only the IPA clone suffered the effect of water conditions, in which the irrigated systems (4, 8, and 12 mm) stood out. In all water management, the OEM clone stood out, presenting higher CW values, with no significant difference between irrigation depths. This behavior was also observed in CP, except in the 8 mm depths, where the IPA and OEM clones stood out (Table 3).
The CAI did not suffer a significant effect from the irrigation depths; however, the OEM and IPA clones demonstrated greater magnitudes when cultivated rainfed and irrigated 8 mm. For the 4 and 12 mm depth, the OEM clone was superior to the other clones (Table 3). In the second experimental cycle, there was no significant effect on PH and PW for clones and irrigation depths.
The mean PH for the IPA, MIU, and OEM clones was, respectively, 76.5, 70.4 and 82.0 cm, and the mean PW of these clones was 66.2, 65.7, and 68.5 cm, in due order. The TNC in the second season showed a significant difference between the clones at depths of 4 and 12 mm and in rainfed cultivation. Furthermore, the MIU clone was affected by the water condition of the culture (p < 0.05). When cultivated in rainfed cactus, the OEM and IPA clones demonstrated higher cladode emissions (26.0 and 17.0 units). When subjected to 4 mm irrigation, the MIU clone demonstrated better performance compared to the other clones, reaching a total of 29.8 cladodes. At the 12 mm irrigation depth, there was a greater emission of cladodes for the MIU and OEM clones (28 and 21.7 units). Regarding the irrigation depths in the MIU clone, it was noted that the highest magnitudes found were in the systems in which 4 and 12 mm were irrigated (Table 3).
When the number of cladodes was quantified in order of appearance in the second cycle, it was noted that the IPA and OEM clones differed from the MIU clone, where they showed greater emission of first-order cladodes (NC1) in rainfed and irrigated cultivation with 8 mm weekly; NC2 from OEM and IPA stood out only when cultivated in rainfed areas. On the other hand, when comparing the emission of third-order cladodes (NC3), the MIU clone irrigated with 12 mm stands out among the rest of the evaluated treatments. The emergence of fourth and fifth-order cladodes (NC4 and NC5) was evidenced only in the MIU clone, where the irrigation depths significantly affected NC4, in which the 4 and 12 mm depths expressed greater magnitudes (4.0 and 2.2 units) (Table 3).
The biometric parameters of the cladodes expressed a significant interaction between both factors (cactus and depths) in the second cycle, except the variable CT, in which a significant effect was found only in the clone factor of the 12 mm depth, where the IPA and MIU clones expressed larger magnitudes (36.2 and 27.5 mm, respectively). The CL was affected by water availability in the IPA clone, in which the crop when irrigated (4, 8, and 12 mm) presented greater magnitudes. It is also possible to observe better performance of the OEM and IPA clones at all irrigation levels. About CW, it is possible to note that the IPA clone suffers the effect of irrigation, in which the water supply independent of the irrigation depth affects the ability of the cladodes to increase their width. However, in general, the OEM and MIU clones demonstrated, respectively, the highest and lowest values (Table 3). Irrigation levels affected the CP of OEM and IPA clones, with rainfed cultivation showing lower magnitudes. Furthermore, the MIU clone presented lower results than the OEM and IPA cactus (Table 3).
The CAI in cycle two suffered a significant effect from the irrigation depths in the IPA clone, in which the rainfed and irrigated systems with 4 mm demonstrated lower CAI (2.1 and 2.3 m2 m−2). The OEM clone demonstrated greater magnitudes when cultivated in rainfed conditions and at depths of 8 and 12 mm (Table 3).
3.3. Forage Yield
Table S2 shows the summarized analysis of variance for plant density (PD), fresh mass (FM), dry mass (DM), and dry mass content (DMC) of forage cactus subjected to different irrigation depths in two successive cycles.
In the first and second experimental cycles, no significant effect was observed for the interaction between depths and cactus (p > 0.05); however, variation was noted in the isolated factors, in which the irrigation depths differed in PD and the cactus differed in all variables (PD, FM, DM, and DMC) (p < 0.05).
The initial PD was 50,000 plants ha−1. The irrigation depths had a significant effect only on the MIU clone, in both cycles, where the systems irrigated with 4 and 12 mm stood out, in which in the first cycle, the final PD was 14,673.9 and 14,945.6 plants ha−1, respectively. In the second experimental cycle, the MIU clone irrigated with 4 and 12 mm weekly, obtained, respectively, a PD of 13,858.7 and 14,673.9 plants ha−1.
Furthermore, in both cycles there was a significant effect for cactus, in all water conditions, in which the OEM clone (mean 41,711.9 and 41,100.5 plants ha−1 for the first and second cycle) stood out from the PD of the IPA cactus (mean 8220.1 and 7812.5 plants ha−1 for the first and second cycle) and the MIU one (mean 10,054.3 and 9578.8 plants ha−1 for the first and second cycle) (Table 4).
Table 4.
Forage yield, dry matter content, and plant density of three cactus, subjected to different levels of irrigation in two successive cycles in the Brazilian semi-arid region.
The OEM clone in both cycles showed higher FM and DM productivity than the IPA and MIU cactus. Furthermore, MF production in the first cycle had a significant effect on irrigation levels only in the OEM clone, in which the crop without irrigation and irrigated with 8 and 12 mm (337.6, 280.8, and 376.6 Mg ha−1) stood out about the 4 mm depth (227.8 Mg ha−1). About DMC, there was no significant effect for irrigation levels in cycles one and two; however, when rainfed cultivated in the second cycle, the MIU clone presented a higher DMC (0.11) (Table 4).
3.4. Morphophysiological Indices
3.4.1. First Experimental Cycle
In Figure 2, the morphophysiological indices (AGR, RGR, CAIR, NAR, and SCA) of three cactus subjected to different irrigation depths in the first experimental cycle are presented. AGR showed differences in behavior and magnitudes, depending on the clone and irrigation depth adopted.
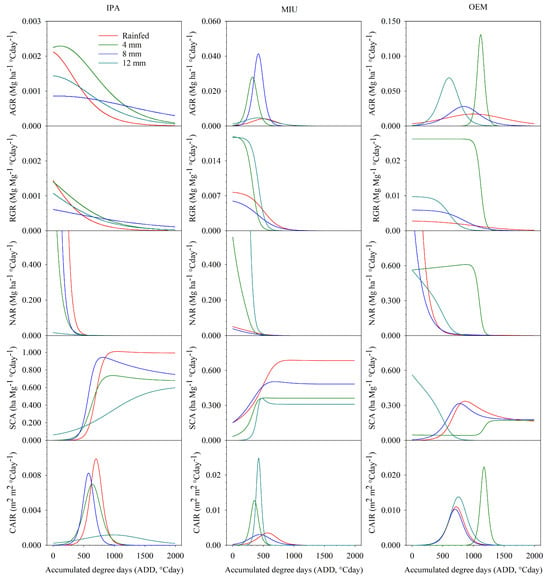
Figure 2.
Morpho-physiological indices (AGR—absolute growth rate; RGR—relative growth rate; CAIR—cladode area index rate; NAR—net assimilation rate; and SCA—specific cladode area) of cactus subjected to different irrigation levels in the first experimental cycle.
The IPA clone demonstrated higher AGR values in the initial period of the cycle, followed by a gradual reduction throughout the growing season. This clone, when cultivated rainfed and irrigated with a 4 mm water depth, demonstrated the highest values (0.0021 and 0.0023 Mg ha−1 °Cday−1, respectively); however, rainfed cultivation presented lower values at end of the cycle (0.00001 Mg ha−1 °Cday−1). On the other hand, when irrigated with 8 mm, the AGR of the IPA clone behaved in a stabilized manner, presenting, respectively, lower and higher values at the beginning and end of the cycle (0.0009 and 0.0003 Mg ha−1 °Cday−1) when compared to the other depths (Figure 2).
The AGR of the MIU clone demonstrated Gaussian behavior in all water conditions, with the highest values reached at depths of 12 and 4 mm (0.0281 and 0.0414 Mg ha−1 °Cday−1, respectively). On the other hand, lower values were observed, respectively, in rainfed cultivation and at 8 mm depth (0.0042 and 0.0047 Mg ha−1 °Cday−1). The OEM clone showed similar behavior to the MIU clone, where the 12 and 4 mm depths reached higher values (0.1308 and 0.0691 Mg ha−1 °Cday−1, respectively) and the rainfed cultivation and 8 mm depth expressed smaller magnitudes (0.0172 and 0.0281) (Figure 2).
In all treatments, RGR presented higher values at the beginning of the cycle and lower values at the end; however, the behavior varied between the cactus and the magnitudes differed between the depths. In the IPA clone, a negative exponential behavior was noted in the four water conditions, where the highest magnitudes at the beginning of the cycle were observed in the rainfed condition and at the 4 mm depth (0.0015 and 0.0014 Mg Mg−1 °Cday−1, respectively); however, at the end of the experimental period, lower values were found under these conditions (0.00001 and 0.00002 Mg Mg−1 °Cday−1) (Figure 2).
For the MIU clone, higher RGR values were initially observed under different water conditions, followed by a marked reduction and subsequent stabilization at the end of the cycle. The 4 and 12 mm depths initially reached higher values (Mg Mg−1 °Cday−1) than the cultivation without irrigation and the 8 mm depth (0.0188 and 0.0187 Mg Mg−1 °Cday−1, respectively); however, at the end of the cycle, all conditions were equal (Figure 2).
The same behavior was observed for the irrigated OEM clone (higher magnitudes of RGR initially, followed by an intense reduction and subsequent stabilization at the end of the season); however, when cultivated under rainfed conditions, it showed a slight gradual reduction in RGR throughout the cycle. The maximum RGR value was observed for this clone when irrigated with 4 mm (0.026 Mg Mg−1 °Cday−1) (Figure 2).
The NAR of the IPA clone expressed greater magnitudes at the beginning of the experimental period, in which the cultivation system without irrigation presented higher values (37.25 Mg ha−1 °Cday−1) and the cultivation irrigated with 12 mm presented the lowest values (0.02 Mg ha−1 °Cday−1); however, throughout the cycle, the values were equivalent in all treatments, reaching minimum values at the end of the cycle (Figure 2). In the MIU clone it is possible to identify similar behaviors, in which the highest value was initially observed at the 12 mm depth (109.33 Mg ha−1 °Cday−1), and the lowest magnitudes were found at the 8 mm depth (0.04 Mg ha−1 °Cday−1); as well as in the IPA clone, the NAR of the MIU clone was equal in all water conditions at the end of the cycle (Figure 2).
The behavior of NAR in the OEM clone followed the same behavior, except the 4 mm depth, which in turn demonstrated stable behavior at the beginning of the cycle, followed by a marked reduction and subsequent stabilization until the end of the cycle. The highest values were found in the initial period of all water conditions, where the maximum value was found in rainfed cultivation (4.52 Mg ha−1 °Cday−1) (Figure 2).
SCA demonstrated sigmoid behavior in the IPA clone, in the initial period of the cycle, the 12 mm irrigation depth expressed greater magnitudes; however, it was soon surpassed by the other water conditions, and at the end of the period it presented lower SCA values. The maximum SCA was found in rainfed cultivation and reached its peak after thermal accumulation of 1048 °Cday. In the MIU clone, SCA also showed sigmoid behavior and rainfed cultivation reached greater magnitudes, as in the IPA clone, demonstrating a pattern for cactus of the genus Nopalea; however, this system required less thermal accumulation to reach the SCA peak (949 °Cday) (Figure 2).
On the other hand, the behavior of SCA in the OEM clone (genus Opuntia) showed different behaviors, varying according to the water availability of the crop. In rainfed cultivation and at depths of 4 and 8 mm, the lowest values were found in the initial period of the cycle. In the 12 mm depth, the peak of SCA occurred at the beginning of the cycle (Figure 2).
The CAI presented sigmoid behavior; therefore, the CAIR expressed Gaussian behavior, in which it initially demonstrates smaller magnitudes, followed by an increase to the apex and a subsequent reduction in rate. Interestingly, the IPA clone grown under rainfed conditions expressed the highest maximum CAIR (0.0099 m2 m−2 °Cday−1); in addition, when irrigated with 12 mm, the peak rate was lower than at all irrigation levels (0.0002 and 0.0099 m2 m−2 °Cday−1). For the MIU clone irrigated with a weekly water depth of 12 mm, a maximum CAIR was identified (0.0414 0.0099 m2 m−2 °Cday−1). The OEM clone had the CAIR peak extended when irrigated with 4 mm, requiring a thermal accumulation of 1173 °Cday (Figure 2).
3.4.2. Second Experimental Cycle
In Figure 3, the morphophysiological indices (AGR, RGR, CAIR, NAR, and SCA) of the second cycle are highlighted. These indices change according to the cactus clone and the water management of the crop. The AGR of the IPA clone when irrigated with 4 and 8 mm depths reached greater magnitudes (0.017 and 0.007 Mg ha−1 °Cday−1, respectively), on the other hand, the cultivation of this clone under rainfed conditions and irrigated with a 12 mm depth showed smaller magnitudes and stable behavior throughout the experimental period, where the AGR peaks were 0.02 and 0.01 Mg ha−1 °Cday−1, respectively. The maximum rate of DM accumulation (AGR) of the MIU clone was observed when irrigated 4 mm weekly (0.024 Mg ha−1 °Cday−1). In the OEM clone, AGR reached maximum values when irrigated 8 and 12 mm weekly (0.031 and 0.062 Mg ha−1 °Cday−1) (Figure 3).
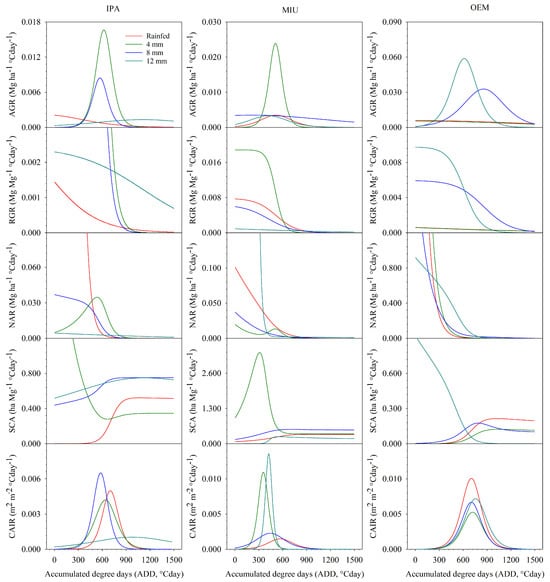
Figure 3.
Morpho-physiological indices (AGR—Absolute growth rate; RGR—Relative growth rate; CAIR—Cladode area index rate; NAR—Net assimilation rate; and SCA—Specific cladode area) of cactus subjected to different irrigation levels in the second experimental cycle.
The highest RGR values were observed in the initial period of the cycle, in which the IPA clone expressed greater magnitudes when irrigated with 4 and 8 mm weekly (0.015 and 0.016 Mg Mg−1 ha−1 °Cday−1). The MIU clone irrigated with 4 mm initially showed higher values (0.019 Mg Mg−1 ha−1 °Cday−1). In turn, the RGR of the OEM clone reached greater magnitudes when irrigated with 8 and 12 mm (0.006 and 0.010 Mg Mg−1 ha−1 °Cday−1). Minimum RGR values were observed in all treatments at the end of the cycle (Figure 3).
Interestingly, the NAR of the IPA clone cultivated under rainfed conditions was vastly superior to other water conditions in the initial period of the cycle, reaching 72.040 Mg ha−1 °Cday−1, followed by the 4 and 8 mm depths (0.034 and 0.036 Mg ha−1 °Cday−1). On the other hand, the MIU clone showed a higher NAR in the initial period of cultivation when irrigated with 12 mm weekly (10,486.520 Mg ha−1 °Cday−1), followed by rainfed cultivation (0.101 Mg ha−1 °Cday−1). In turn, the OEM clone obtained higher NAR in rainfed and irrigated crops with 4 mm, where the maximum values were 9.478 and 15.558 Mg ha−1 °Cday−1, respectively (Figure 3).
When irrigated with 4 mm, the IPA clone demonstrated the highest magnitudes in the initial period (3.089 ha Mg−1), on the other hand, this behavior was reversed for the other treatments (rainfed, 8 mm and 12 mm), in which the lowest values of SCA were initially expressed. This behavior was also noted for the MIU clone, in which the 4 mm depth had a maximum SCA of 3.36 ha Mg−1. In contrast, in the clone of the genus Opuntia (OEM), the maximum SCA value occurred at a depth of 12 mm (0.11 ha Mg−1) (Figure 3).
CAIR showed Gaussian behavior in all treatments. In the IPA clone, the CAIR peak (0.007 m2 m−2 °Cday−1) occurred after a thermal accumulation of 580 °Cday, when irrigated with the 8 mm depth. The MIU clone reached maximum CAIR when irrigated with 12 mm weekly (0.011 m2 m−2 °Cday−1). As for the OEM clone, the peak of CAIR occurred in rainfed cultivation (0.010 m2 m−2 °Cday−1) (Figure 3).
3.5. Phenophases and Cutting Moment
3.5.1. First Experimental Cycle
Figure 4 shows the phenological dynamics and the ideal moment for cutting the first cycle of three cactus, cultivated under different irrigation levels. The number of phenophases varied in different cactus, with the IPA clone showing P2 and P3 in all water conditions, the MIU clone reaching P4 in all irrigation depths and the OEM clone showing P2 and P3 when cultivated in rainfed and irrigated conditions, with 4 and 8 mm weekly and reaching P4 when 12 mm of water was applied. Furthermore, there is a difference in the duration of each phenophase depending on the clone and water supply.
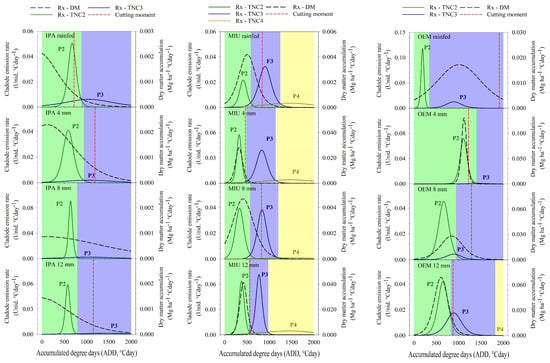
Figure 4.
Phenophases (P2—Phenophase 2; P3—Phenophase 3; P4—Phenophase 4), dry matter accumulation rate (Rx—DM), and ideal cutting moment of three cactus cactuses subjected to different irrigation depths in the first experimental cycle.
The IPA clone did not express a conclusive response regarding the effect of the depths on the duration of each phenological phase; however, the MIU clone exhibited a tendency towards a reduction in the duration of P2 and P3, which caused a prolongation of P4, whereas in the OEM clone, it was noted that when cultivated under rainfed conditions, there was a shorter duration of P2 and an extension of P3. However, when irrigation levels were applied, this behavior was gradually reversed, with a reduction in P2 and an extension of P3 as the water increases in cultivation occurred (Figure 4).
About the cladode emission peak in each phenophase, it is also possible to observe variation between cactus and irrigation depths. In the IPA clone, the peak of second-order cladode emission (P2) occurred when the weekly irrigation volume was 8 mm, reaching a cladode emission rate of 0.091 units °Cday−1; in addition, there was a drastic reduction in the emission of third-order cladodes (P3), in which the emission peak occurred when the IPA clone was cultivated in rainfed conditions (0.006 units °Cday−1).
When evaluating the effect of irrigation depths on the emission of cladodes from each phenological phase of the MIU clone, it is noted that there was a maximum emission rate of second, third and fourth order cladodes at the 12 mm depth (0.064, 0.071, and 0.005 units °Cday−1, respectively). Interestingly, the OEM clone reached the peak emission rate of first- and second-order cladodes when cultivated in rainfed conditions (0.12 and 0.013 units °Cday−1, respectively).
The cutting moment varied in the different systems, demonstrating more clearly in the OEM clone a gradual tendency to bring forward the harvest as the irrigation depth was increased. Therefore, this clone, when irrigated with 12 mm per week, reached 25% of the DM accumulation peak (cut-off moment) at 863 °Cday. On the other hand, the IPA clone did not show this behavior, since in rainfed cultivation the ideal harvest time was advanced in relation to irrigation treatments, reaching the cutoff point at 717 °Cday. As for the MIU clone, it was not possible to identify a trend in relation to the moment of harvest, where on the 4 mm depth, it was observed at 463 °Cday (Figure 4).
3.5.2. Second Experimental Cycle
As in the first experimental cycle, there was a difference in the phenological dynamics and moment of cutting, in which there was a difference between the three cactus and the effect of the irrigation depths. The IPA clone reached P2 and P3 when cultivated, rainfed and irrigated with 4 and 8 mm water depths; however, when it was irrigated 12 mm weekly, the clone did not express P3. The MIU clone expressed up to P4 in all treatments, while the OEM clone reached P3 in all systems (Figure 5).
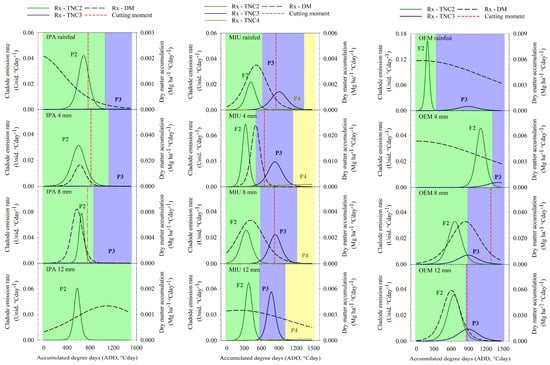
Figure 5.
Phenophases (P2—Phenophase 2; P3—Phenophase 3; P4—Phenophase 4), dry matter accumulation rate (Rx—DM) and ideal moment of cutting of three cactus subjected to different irrigation levels in the second experimental cycle.
Regarding the duration of the phenophases, there was differentiation according to the irrigation depth and clone adopted in management. The IPA clone, when irrigated with 12 mm weekly, only showed emission of second-order cladodes throughout the cycle (1472 °Cday); on the other hand, when subjected to an 8 mm water depth, a reduction in P2 was noted (1040 °Cday) and a prolongation of P3 (432 °Cday) (Figure 5). In general, the MIU clone when irrigated, regardless of the water volume applied via irrigation, presents a slight reduction in phenophases 1 and 2, which provides an increase in P3. The OEM clone cultivated under rainfed conditions showed a shorter duration of P2 (353 °Cday−1) and a longer duration of P3 (1119 °Cday−1), about the irrigated cultivation at different depths. However, when comparing the depths to each other, it was found that the 8 and 12 mm depths compared to the 4 mm weekly depths reduced the duration of P2 and extended the duration of P3 (Figure 5).
The maximum emission rates of cladodes in the different phenophases varied between cactus and depths. In the IPA clone, the peak of secondary and tertiary cladode emission occurred when irrigated with an 8 mm blade (0.078 and 0.0004 units °Cday−1, respectively), after thermal accumulation at 640 and 1051 °Cday, in the same order. When the 4 mm irrigation depth was applied to the MIU clone, the maximum cladode emission rate was noted in phenophases 2 and 4 (0.073 and 0.003 units °Cday−1, in due order), after reaching, respectively, 324 and 1450 °Cday; however, in phenophase 3, the maximum rate of cladode emission was observed in the 12 mm slide (0.029 units °Cday−1), after a thermal accumulation of 833 °Cday−1. Intriguingly, the OEM clone expressed higher emission rates of second- and third-order cladodes when cultivated in rainfed conditions, reaching 0.163 and 0.010 units °Cday−1, respectively; these cladode emission peaks occurred after the culture accumulated 189 and 891 °Cday, in the appropriate order (Figure 5).
The harvest time of the IPA clone was not affected by the irrigation depths, except when irrigated 12 mm weekly, which delayed the accumulation of DM and did not reach the cutting moment. This behavior was also repeated for the MIU clone, where the water input of 12 mm did not reach the ideal harvest time. The OEM clone showed a tendency to gradually bring forward the harvest as the irrigation depths increased, with the 12 mm treatment bringing forward the cutoff time to 873 °Cday (Figure 5).
3.6. Water Use and Economic Indicators
In Table 5, information for crops cycle one and two, such as rainfall (R), irrigation (ID) and actual evapotranspiration (ET), are presented. In addition, economic information on cultivation systems was verified (revenue, variable cost, fixed cost, opportunity cost, gross margin, and net margin). With this information, nine economic and water use indices were calculated: WUEc, *WUEc, WPc, *WPc, WPI, GEWPI, NEWPI, EWPc and EWPI.
Table 5.
Water use and economic indicators in two successive cycles of three cactus subjected to different levels of irrigation.
The rainfall volumes accumulated in the first and second experimental cycle were 816.2 and 719.1 mm, respectively. On the other hand, the volumes of water irrigated in the first cycle were 116.5; 233.0; and 349.4 mm at 4, 8, and 12 mm depths, respectively. The accumulation of irrigation in the second cycle was 40.0; 80.1; and 120.1 mm, respectively, for the 4, 8 and 12 mm weekly depths. The ET was 2017.5 mm in the first season, and 1579.3 mm in the second (Table 5).
Total system revenues fluctuated between the cactus (IPA, MIU, and OEM) and depths (0, 4, 8 and 12 mm) evaluated. In general, the OEM clone had an average revenue for the first and second cycle higher than the MIU and OEM cactus. In the initial season, the cultivation of the OEM clone yielded an average of USD 8802, and in the subsequent cycle the average revenue was USD 6971. The MIU clone in cycles one and two presented revenue of USD 1464 and USD 1269, respectively. In turn, the IPA clone demonstrated average revenue of USD 1144 in the first period and USD 922 in the subsequent period (Table 5).
Cultivation costs were made up of fixed costs and variable costs, in which in the first cycle, fixed costs in all systems were USD 4666. However, as the crops were already established in the field, there were no new investments in the second cycle. On the other hand, variable costs fluctuated according to the system and cultivation cycle, in which in the first cycle, variable costs were USD 255, 510, 765, and, 892 for depths of 0, 4, 8, and 12 mm, respectively. In the second cycle, the variable costs of the 0, 4, 8 and 12 mm depths were USD 188, 377, 565, and 660, respectively (Table 5).
The opportunity cost, which refers to the possible value of revenue if cladode units were sold at USD 0.03, expressed maximum values for the MIU clone in both cycles, where the magnitudes averaged USD 36,240 in the first cycle and USD 22,943 in the second (Table 5).
In general, only the OEM clone made a profit in the first cycle (positive net margin); this is because the forage yield of the IPA and MIU cactus was low, which made it possible to pay the variable costs and only a fraction of the fixed costs. However, in the second cycle, there was a net margin in all cactus since there were no fixed costs; however, the OEM clone stood out again (Table 5).
Regarding economic and water use indicators, WUEc expresses a reduction in efficiency as water availability increases in cactus cultivation. On the other hand, it is possible to notice an inversely proportional trend, in which the water supply increased the magnitudes of *WUEc for the three cactus evaluated. In general, the IPA clone, when irrigated with 4 mm weekly in the first and second cycle, expressed greater magnitudes for the WPc, *WPc, WPI, GEWPI, EWPC and EWPI indices; however, the NEWPI index expressed a lower magnitude in this system in the initial period since fixed costs were high. In the MIU clone, the WPc, and *WPc indices expressed higher values when 12 mm was irrigated in both experimental cycles. However, the WPI, GEWPI, EWPc, and EWPI indices were higher when irrigating 4 mm weekly. The OEM clone generally obtained greater efficiencies in all indices, presenting greater magnitudes when irrigated with the 4 mm depth (Table 5).
4. Discussion
4.1. Water Balance and Water Use Efficiency and Economic Indicators
In general, the variation in water storage (ΔA) in the soil was positively affected by the irrigation depths, in which the water supply to the soil favored positive water flow, regardless of the clone evaluated. The authors of [35] found that ΔA is highly dependent on the water input into the system, atmospheric demand, soil characteristics and the capacity to capture water by the roots. The actual evapotranspiration (ET) of the systems was also positively affected by the irrigation depths. This occurred due to the increase in water availability in the soil, favoring water consumption. However, ref. [36] highlighted that evaluating the water balance of systems composed of forage cactus without considering the water retained by the plant structure of the crop can lead to mistaken interpretations since the difference in the spatial architecture of cactus affects the water supply of the crop.
Studying the soil hydrodynamics of a crop and using this information, together with financial data, makes it possible to apply combined economic and water efficiency indicators, in addition to increasing the accuracy of the indices, so that they represent the real condition of the crop. Water-economic indicators provide useful information for policy makers in charge of promoting irrigation while preserving the environment [37,38,39]. In the current study, greater efficiencies were observed when a weekly irrigation depth of 4 mm was adopted, so that the increase in water input via irrigation increases costs and the crop does not respond positively in productive terms; with this, the magnitudes of efficiency indicators decrease.
The monetary indices of the OEM clone are vastly superior compared to the cactus IPA and MIU. This is due to its high biomass production, which enables profitable cultivation in the first cycle. Despite this, the MIU clone stood out economically when the destination of production was sale as seed; this is due to the high number of cladodes in this clone.
4.2. Morphological Characteristics and Forage Yield
The three cactus clones evaluated demonstrated productive and morphological differences. Recent studies report that these results are related to the intrinsic characteristics of each genus and clone, such as the plant’s growth habit, size, shape, and number of cladodes [7,9,40,41].
The MIU clone has a greater number of higher-order cladodes (third-, fourth- and fifth-order); however, its cladodes express a lower photosynthetic area than the other cacti. The OEM and IPA clones presented a greater number of lower cladodes (first- and second-order). Among the cactus evaluated, OEM expresses a greater photosynthetic area than the individual cladodes [32,42]. Furthermore, agricultural management results in different responses to cultivation conditions, modifying structural aspects of cladodes (i.e., width, length, thickness, and perimeter of cladodes) and plants (i.e., height and width of plants and number of cladodes) [7,15,27,33,40].
The shape and distribution of the cladodes in the canopy of the OEM clone enables a higher CAI than cactus of the genus Nopalea, which favors the interception of solar radiation and leads to a significant increase in forage yield. Furthermore, cacti of the genus Nopalea generally present higher mortality and lower biomass productivity when compared to cacti of the genus Opuntia (Table S2).
In general, water management did not have a significant effect on the morphological characteristics and forage yield of the crop. According to [24], in rainy years, water supply via irrigation does not significantly alter the morphological and productive characteristics of forage cactus. The accumulated rainfall in the first and second seasons was 928.2 mm (~14 months) and 719.1 mm (~11 months), respectively, (Figure 2). These water conditions restricted the cactus’ responses to water input via irrigation.
4.3. Phenophases and Cutting Moment
To monitor the development of the crop in the field and optimize decision-making, it is necessary to know the phenological phases of forage cactus [43,44]. The results showed that the MIU clone expressed a greater number of phenophases when compared to the IPA and OEM cactus (Figure 3 and Figure 4); this behavior is linked to the morphological characteristics of the clone (Figure 3 and Figure 4) [18,42].
In general, soil water conditions can alter the dynamics of phenophases [15,43,44]. In this study, the MIU clone gradually anticipated phenophase 4, as soil water availability increased with irrigation levels (rainfed, 4, 8, and 12 mm). As a result, there was a reduction in the duration of phenophases 2 and 3, which resulted in a prolongation of phenophase 4.
In the OEM clone, the shortest duration of phenophase 2 occurred in rainfed cultivation; however, when irrigated, the tendency to reduce the duration of P2 occurred with the increase in water depth, reaching P4 when irrigated with 12 mm weekly in the dry season in the first cycle. A reduction in the intensity of this behavior was noted in the phenophases of the IPA clone. These phenological dynamics may be associated with greater water availability, which allows for a higher rate of emission of higher-order cladodes [18,45].
The growth dynamics of the cactus were affected by water management and provided variations in the speed of accumulation of dry matter in the systems, because of which the harvesting time of the crop was also affected. In general, the use of supplementary irrigation in the IPA clone delayed harvest. The MIU clone generally expresses the peak of the dry matter accumulation rate still in P2; this makes it possible to perform the cut in P3, due to the low emission of fourth-order cladodes. The authors of [44] point out that the emission of higher-order cladodes can reduce the plant’s energy efficiency.
In general, the systems composed of the OEM clone showed higher DM accumulation rates, in which the increase in water supply via irrigation anticipated the cutting moment. The authors of [18], evaluating the controlled water deficit in forage cactus, found that greater soil water availability, even if in deficit for the crop, anticipates the harvest and optimizes the forage yield of these systems over time. This behavior was also observed by [14], where mulch was used on the soil, reducing water losses to the atmosphere, and thus increasing water use by the crop.
4.4. Morphophysiological Indices
The highest magnitudes of absolute growth rate (AGR) were expressed in the OEM clone, which responds proportionally to CAIR. The high photosynthetic area of cladodes allows for the greater interception of solar radiation, which is used by the chloroplast for CO2 assimilation and transpiration [46,47]. When irrigated at 12 mm weekly, the OEM clone expressed high AGR, since water supply via irrigation enhances the emission of cladodes [43,44,45].
The relative growth rate (RGR) expresses the highest values at the beginning of the cycle; however, factors such as self-shading increase the accumulation of non-photosynthetic biomass, the aging and renewal of tissues, and the reduction in nutrient absorption [18]. The photosynthetic efficiency of cladodes is expressed by the net assimilation rate (NAR), which is directly related to the plant’s growth habit, in which initially there is less shading that provides greater photosynthetic efficiency. The authors of [48], evaluating CO2 assimilation methods in Opuntia ficus-indica plants, found that shading was responsible for a 50% reduction.
The disposition of dry matter in the photosynthetic area of the cladodes is given by the SCA; this index presents a behavior inversely proportional to the thickness of the cladodes, in which greater thicknesses of the cladodes reach lower magnitudes of SCA [18,24]. In the present study, this behavior was evident in the first season, where rainfed cultivation, which generally has smaller thicknesses due to low water availability in the soil, reached higher SCA values. In turn, in the second experimental cycle there was a reduction in dry matter accumulation when cactus was cultivated without irrigation; this behavior affected the standard dynamics of SCA.
5. Conclusions
The present study showed that cacti resistant to carmine cochineal express different canopy architectures, in which the clone Miúda (N. cochenillifera (L.) Salm-Dyck) has a greater number of cladodes in relation to the IPA Sertânia cactus (N. cochenillifera (L.) Salm-Dyck) e Orelha de Elefante Mexicana (O. stricta (Haw.) Haw.). This favors crop yields when production is destined for seed sales. On the other hand, the clone Orelha de Elefante Mexicana presents higher forage yield, regardless of the water condition of the crop. The different levels of irrigation did not affect the overall productivity of forage cactus; however, a growing trend was noted of a reduction in the duration of the initial phenophases, which made it possible to anticipate the harvest when water availability was increased, favoring the productive optimization of cultivation in the time.
In general, there was no difference between cactus in terms of evapotranspiration; however, it was clear that the increase in water supply increased the use of water by the crop, despite reducing the economic and efficiency rates of water use in crops. Thus, we confirm the hypothesis that greater water availability in the system positively affects the growth and production dynamics of cactus; however, it negatively affects water-economic efficiency indices. However, it is necessary to understand in future studies how the crop behaves in dry years, since rainfall accumulation in the experimental period was high, which may have attenuated the crop’s responses to irrigation levels.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy14040691/s1, Table S1: Summary of the analysis of variance of biometric variables of three cactus clones subjected to different irrigation depths.; Table S2: Summary of the analysis of variance of forage yield of cactus subjected to different levels of irrigation in two successive cycles.
Author Contributions
Conceptualization, G.d.N.A.J., R.M.C.L. and T.G.F.d.S.; methodology, G.d.N.A.J., R.M.C.L., J.E.F.d.M., L.S.B.d.S. and T.G.F.d.S.; validation, G.d.N.A.J., J.E.F.d.M., L.S.B.d.S. and C.P.A.; formal analysis, G.d.N.A.J., R.M.C.L., J.E.F.d.M., C.P.A., C.A.A.d.S., A.C.d.S.A., A.M.d.R.F.J., L.S.B.d.S., D.d.S.E. and T.G.F.d.S.; investigation, G.d.N.A.J., R.M.C.L., J.E.F.d.M., T.G.F.d.S., C.P.A., A.M.d.R.F.J. and C.A.A.d.S.; resources, T.G.F.d.S.; data curation, R.M.C.L. and T.G.F.d.S.; writing—original draft preparation, G.d.N.A.J., R.M.C.L. and J.E.F.d.M.; writing—review and editing, G.d.N.A.J., J.E.F.d.M. and T.G.F.d.S.; visualization, T.G.F.d.S., A.C.d.S.A., J.E.F.d.M. and L.S.B.d.S.; supervision, L.S.B.d.S., J.E.F.d.M. and T.G.F.d.S.; project administration, T.G.F.d.S.; funding acquisition, T.G.F.d.S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Foundation for the Support of Science and Technology of Pernambuco [APQ-0428-5.01/21], and the National Council for Scientific and Technological Development [402622/2021-9 and 309558/2021-2].
Data Availability Statement
Data are contained within the article.
Acknowledgments
The authors would like to thank the Coordination for the Improvement of Higher Education Personnel (CAPES) and the Foundation for the Support of Science and Technology of Pernambuco (FACEPE) for granting the scholarship to students, and the members of the Brazilian Semiarid Agrometeorology Group—GAS for assistance in experimenting.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Chikwanha, O.C.; Mupfiga, S.; Olagbegi, B.R.; Katiyatiya, C.L.F.; Molotsi, A.H.; Abiodun, B.J.; Dzama, K.; Mapiye, C. Impact of water scarcity on dryland sheep meat production and quality: Key recovery and resilience strategies. J. Arid. Environ. 2021, 190, 104511. [Google Scholar] [CrossRef]
- Lai, X.; Shen, Y.; Wang, Z.; Ma, J.; Yang, X.; Ma, L. Impact of precipitation variation on summer forage crop productivity and precipitation use efficiency in a semi-arid environment. Eur. J. Agron. 2022, 141, 126616. [Google Scholar] [CrossRef]
- Lal, B.; Sarkar, S.; Gautam, P.; Meena, R.L.; Bhatt, R.S.; Sahoo, A. Environmental impacts and resource use for sheep production in semi-arid India investigated by life cycle assessment. J. Clean. Prod. 2022, 345, 131088. [Google Scholar] [CrossRef]
- Kumar, S.; Mishra, A.K.; Pramanik, S.; Mamidanna, S.; Whitbread, A. Climate risk, vulnerability, and resilience: Supporting livelihood of smallholders in semiarid India. Land Use Policy 2020, 97, 104729. [Google Scholar] [CrossRef]
- Fust, P.; Schlecht, E. Importance of timing: Vulnerability of semi-arid rangeland systems to increased variability in temporal distribution of rainfall events as predicted by future climate change. Ecol. Model. 2022, 468, 109961. [Google Scholar] [CrossRef]
- Queiroz, M.G.; Silva, T.G.F.; Zolnier, S.; Jardim, A.M.R.F.; Souza, C.A.A.; Araújo Júnior, G.N.; Morais, J.E.F.; Souza, L.S.B. Spatial and temporal dynamics of soil moisture for surfaces with a change in land use in the semi-arid region of Brazil. Catena 2020, 188, 104457. [Google Scholar] [CrossRef]
- Jardim, A.M.R.F.; Souza, L.S.B.; Alves, C.P.; Araújo, J.F.N.; Souza, C.A.A.; Pinheiro, A.T.; Araújo, G.G.L.; Campos, F.S.; Tabosa, J.N.; Silva, T.G.F. Intercropping forage cactus with sorghum affects the morphophysiology and phenology of forage cactus. Afr. J. Range Forage Sci. 2021, 40, 129–140. [Google Scholar] [CrossRef]
- Alves, C.P.; Jardim, A.M.R.F.; Araújo Júnior, G.N.; Souza, L.S.B.; Araújo, G.G.L.; Souza, C.A.A.; Salvador, K.R.S.; Leite, R.M.C.; Pinheiro, A.G.; Silva, T.G.F. How to enhance the agronomic performance of cactus-sorghum intercropped system: Planting configurations, density, and orientation. Ind. Crop. Prod. 2022, 184, 115059. [Google Scholar] [CrossRef]
- Araújo, G.N., Jr.; Silva, T.G.F.; Souza, L.S.B.; Souza, M.S.; Araújo, G.G.L.; Moura, M.S.B.; Santos, J.P.A.S.; Jardim, A.M.R.F.; Alves, C.P.; Alves, H.K.M.N. Productivity, bromatological composition and economic benefits of using irrigation in the forage cactus under regulated deficit irrigation in a semiarid environment. Bragantia 2021, 80, e1221. [Google Scholar] [CrossRef]
- Campos, A.R.F.; Silva, A.J.P.; Van Lier, Q.J.; Nascimento, F.A.L.; Fernandes, R.D.M.; Almeida, J.N.; Paz, V.P.S. Yield and morphology of forage cactus cultivars under drip irrigation management based on soil water matric potential thresholds. J. Arid. Environ. 2021, 193, 104564. [Google Scholar] [CrossRef]
- Lankford, B.; Pringle, C.; McCosh, J.; Shabalala, M.; Hess, T.; Knox, J.W. Irrigation area, efficiency, and water storage mediate the drought resilience of irrigated agriculture in a semi-arid catchment. Sci. Total Environ. 2023, 859, 160263. [Google Scholar] [CrossRef]
- Jardim, A.M.R.F.; Santos, H.R.B.; Alves, H.K.M.N.; Ferreira-Silva, S.L.; Souza, L.S.B.; Araújo Júnior, G.N.; Souza, M.S.; Araújo, G.G.L.; Souza, C.A.A.; Silva, T.G.F. Genotypic differences relative photochemical activity, inorganic and organic solutes and yield performance in clones of the forage cactus under semi-arid environment. Plant Physiol. Biochem. 2021, 162, 421–430. [Google Scholar] [CrossRef]
- Silva, M.V.; Pandorf, H.; Almeida, G.L.P.; Lima, R.P.; Santos, A.; Jardim, A.M.R.F.; Rolim, M.M.; Silva, J.L.B.; Batista, P.H.D.; Silva, R.A.B.; et al. Spatio-temporal monitoring of soil and plant indicators under forage cactus cultivation by geoprocessing in Brazilian semi-arid region. J. S. Am. Earth Sci. 2021, 107, 103155. [Google Scholar] [CrossRef]
- Alves, H.K.M.N.; Silva, T.G.F.; Jardim, A.M.R.F.; Souza, L.S.B.; Araújo Júnior, G.N.; Souza, C.A.A.; Moura, M.S.B.; Araújo, G.G.L.; Campos, F.S.; Cruz Neto, J.F. The use mulch in cultivating the forage cactus optimizes yield in less time and increases the water use efficiency of the crop. Irrig. Drain. 2022, 72, 75–89. [Google Scholar] [CrossRef]
- Souza, M.S.; Araújo Júnior, G.N.; Souza, L.S.B.; Jardim, A.M.R.F.; Silva, G.I.N.; Araújo, G.G.L.; Campos, F.S.; Leite, M.L.M.V.; Tabosa, J.N.; Silva, T.G. Forage yield, competition and economic benefit of intercropping cactus and millet with mulch in a semi-arid environment. Afr. J. Range Forage Sci. 2022, 40, 219–230. [Google Scholar] [CrossRef]
- Rocha Filho, R.R.; Santos, D.C.; Véras, A.S.C.; Siqueira, M.C.B.; Novaes, L.P.; Mora-Luna, R.; Monteiro, C.C.F.; Ferreira, M.A. Can spineless forage cactus be the queen of forage crops in dryland areas? J. Arid. Environ. 2021, 186, 104426. [Google Scholar] [CrossRef]
- Araújo Júnior, G.N.; Jardim, A.M.R.F.; Silva, M.J.; Alves, C.P.; Souza, C.A.A.; Costa, S.A.T.; Cunha, M.V.; Simões, A.N.; Silva, J.R.I.; Souza, L.S.B.; et al. Growth dynamics and accumulation of forage mass of forage cactus clones as affected by meteorological variables and water regime. Eur. J. Agron. 2021, 131, 126375. [Google Scholar] [CrossRef]
- Araújo Júnior, G.N.; Silva, T.G.F.; Souza, L.S.B.; Araújo, G.G.L.; Moura, M.S.B.; Alves, C.P.; Salvador, K.R.S.; Souza, C.A.A.; Montenegro, A.A.A.; Silva, M.J. Phenophases, morphophysiological indices and cutting time in clone of the forage cacti under controlled water regimes in a semiarid environment. J. Arid. Environ. 2021, 190, 104510. [Google Scholar] [CrossRef]
- Lima, L.R.; Silva, T.G.F.; Jardim, A.M.R.F.; Souza, C.A.A.; Queiroz, M.G.; Tabosa, J.N. Growth, water use, and efficiency of forage cactus sorghum intercropping under different water depths. Rev. Bras. Eng. Agríc. Ambient. 2018, 22, 113–118. [Google Scholar] [CrossRef]
- Silva, E.C.B.; Lima, J.R.S.; Antonino, A.C.D.; Melo, A.A.S.; Souza, E.S.; Souza, R.M.S.; Silva, V.P.; Oliveira, C.L. Efeito da Irrigação Suplementar na Produtividade e Eficiência no Uso de Água da Palma Forrageira. Rev. Bras. Geogr. Física 2020, 13, 2744–2759. [Google Scholar] [CrossRef]
- Fonseca, V.A.; Santos, M.R.; Silva, J.A.; Donato, S.L.R.; Rodrigues, C.S.; Brito, C.F.B. Morpho-physiology, yield, and water-use efficiency of Opuntia ficus-indica irrigated with saline water. Acta Sci.—Agron. 2019, 41, 42631. [Google Scholar] [CrossRef]
- Fernández, J.E.; Alcon, F.; Diaz-Espejo, A.; Hernandez-Santana, V.; Cuevas, M.V. Water use indicators and economic analysis for on-farm irrigation decision: A case study of a super high density olive tree orchard. Agric. Water Manag. 2020, 237, 106074. [Google Scholar] [CrossRef]
- Singh, A. Judicious and optimal use of water and land resources for long-term agricultural sustainability. Resour. Conserv. Recycl. Adv. 2022, 13, 200067. [Google Scholar] [CrossRef]
- Queiroz, M.G.; Silva, T.G.F.; Zolnier, S.; Silva, S.M.S.; Lima, L.R.; Alves, J.O. Características morfofisiológicas e produtividade da palma forrageira em diferentes lâminas de irrigação. Rev. Bras. Eng. Agríc. Ambient. 2015, 19, 931–938. [Google Scholar] [CrossRef]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Gonçalves, J.L.M.; Sparovek, G. Koppen’s climate classification map of Brazil. Meteorol. Z. 2013, 22, 711–728. [Google Scholar] [CrossRef]
- Beck, H.E.; Zimmerman, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef]
- Pereira, P.D.C.; Silva, T.G.F.; Zolnier, S.; Morais, J.E.F.; Santos, D.C. Growth evolution of cactus forage drip irrigated. Rev. Caatinga 2015, 28, 184–195. [Google Scholar] [CrossRef]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; USDA-Natural Resources Conservation Service: Washington, DC, USA, 2014. [Google Scholar]
- Richards, L.A. Diagnose and Improvement of Saline and Alkali Soils; Handbook, 60; USDA: Washington, DC, USA, 1954; 60p. [Google Scholar]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration: Guidelines for Computing Crop Requirements; FAO. Irrigation and Dranaige Paper, 56; FAO: Rome, Italy, 1998; 300p. [Google Scholar]
- Libardi, P.L. Dinâmica da Água no Solo; Editora da Universidade de São Paulo: São Paulo, Brazil, 2005. [Google Scholar]
- Silva, T.G.F.; Miranda, K.R.; Santos, D.C.; Queiroz, M.G.; Silva, M.C.; Cruz Neto, J.F.; Araújo, J.E.M. Área do cladódio de clones de palma forrageira: Modelagem, análise e aplicabilidade. Rev. Bras. Ciências Agrárias 2014, 9, 633–641. [Google Scholar] [CrossRef]
- Souza, M.S.; Silva, T.G.F.; Souza, L.S.B.; Alves, H.C.M.N.; Leite, R.M.C.; Souza, C.A.A.; Araújo, G.G.L.; Campos, F.S.; Silva, M.J.D.; Souza, P.J.D.O.P. Growth, phenology and harvesting time oof cactus-millet intercropping system under biotic mulching. Arch. Agron. Soil Sci. 2021, 67, 1852553. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 25 January 2023).
- Araújo Primo, J.T.; Silva, T.G.F.; Silva, S.M.S.; Moura, M.S.B.; Souza, L.S.B. Calibração de sondas capacitivas, funções físico-hídricas e variação do armazenamento de água em um argissolo cultivado com palma forrageira. Rev. Ceres 2015, 62, 20–29. [Google Scholar] [CrossRef]
- Morais, J.E.F.; Silva, T.G.F.; Queiroz, M.G.; Araújo, G.G.L.; Moura, M.S.B.; Araújo Júnior, G.N. Hydrodynamic changes of the soil-cactus interface, effective actual evapotranspiration, and its water efficiency under irrigation. Rev. Bras. Eng. Agríc. Ambient. 2017, 21, 273–278. [Google Scholar] [CrossRef]
- Expósito, A.; Berbel, J. Agricultural irrigation water use in a closed basin and the impacts on water productivity: The case of the Guadalquivir River Basin (Southern Spain). Water 2017, 9, 136. [Google Scholar] [CrossRef]
- Grafton, R.Q.; Williams, J.; Perry, C.J.; Molle, F.; Ringler, C.; Steduto, P.; Udall, B.; Wheeler, S.A.; Wang, Y.; Garrick, D.; et al. The paradox of irrigation efficiency. Science 2018, 361, 748–750. [Google Scholar] [CrossRef] [PubMed]
- Velasco-Muñoz, J.F.; Aznar-Sánchez, J.; Belmonte-Ureña, L.J.; Román-Sánchez, I.M. Sustainable water use in agriculture: A review of worldwide research. Sustainability 2018, 10, 1084. [Google Scholar] [CrossRef]
- Dubeux Júnior, J.C.B.; Santos, M.V.F.; Cunha, M.V.; Santos, D.C.; Souza, R.T.A.; Mello, A.C.L.; Souza, T.C. Cactus (Opuntia and Nopalea) nutritive value: A review. Anim. Feed. Sci. Technol. 2021, 275, 114890. [Google Scholar] [CrossRef]
- Araújo, G.N., Jr.; Souza, L.S.B.; Jardim, A.M.R.F.; Alves, C.P.; Assis, M.C.S.; Silva, J.O.N.; Souza, C.A.A.; Pinheiro, A.G.; Oliveira, A.C.; Silva, T.G.F. The association between morphological characteristics and yield in forage cactus cactus varies according to water regimes: A principal component analysis. J. Prof. Assoc. Cactus Dev. 2022, 24, 83–95. [Google Scholar] [CrossRef]
- Silva, T.G.F.; Araújo Primo, J.T.; Morais, J.E.F.; Diniz, W.J.S.; Souza, C.A.A.; Silva, M.C. Crescimento e produtividade de clones de palma forrageira no Semiárido e relações com variáveis meteorológicas. Rev. Caat 2015, 28, 10–18. [Google Scholar]
- Cruz, P.E.; Pavón, N.P. Reproductive phenology of Isolatocereus dumortieri (Cactaceae) in semiarid scrub in central Mexico: Effect of rain during the dry season. J. Arid. Environ. 2013, 92, 53–58. [Google Scholar] [CrossRef]
- Amorim, D.M.; Silva, T.G.F.; Pereira, P.C.; Souza, L.S.B.; Minuzzi, R.B. Phenophases and cutting time of forage cactus under irrigation and cropping systems. Pesqui. Agropecu. Trop. 2017, 47, 62–71. [Google Scholar] [CrossRef]
- Arba, M.; Falisse, A.; Choukr-Allah, R.; Sindic, M. Effect of irrigation at critical stages on the phenology of flowering and fruiting of the cactus Opuntia spp. Braz. J. Biol. 2018, 78, 653–660. [Google Scholar] [CrossRef]
- Garofalo, P.; Rinaldi, M. Leaf gas exchange and radiation use efficiency of sunflower (Helianthus annum L.) in response to different deficit irrigation strategies: From solar radiation to plant growth analysis. Eur. J. Agron. 2015, 64, 88–97. [Google Scholar] [CrossRef]
- Zhang, Z.; Christensen, M.; Nan, Z.; Whish, J.P.M.; Bell, L.W.; Wang, J.; Wang, Z.; Sim, R. Plant development and solar radiation interception of four annual forage plants in response to sowing date in a semi-arid environment. Ind. Crop. Prod. 2019, 131, 41–53. [Google Scholar] [CrossRef]
- Franck, N.; Muños, V.; Alfaro, F.; Arancibia, D.; Pérez-Quezada, J. Estimating the Carbon Assimilation of Growing Cactus Pear Cladodes through Different Methods. Acta Hortic. 2013, 19, 157–164. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).