1. Introduction
Currently, there is growing interest in ancient, forgotten cereal species, whose lower yields are partially compensated for by the high nutritional value of the grains [
1]. Spelt wheat, known for more than 5000 years and increasingly grown on organic farms, is currently enjoying a renaissance. In modern agriculture, increasing attention is being paid to the use of organic grain growing technologies based on the application of bacterial formulations. Among the available crop management techniques, biofertilizers have been attracting increasing attention from researchers around the world over the past few decades [
2]. Biofertilizers can be defined as a group of living microorganisms used in agriculture through seed inoculation and foliar and soil application [
3]. A proven way of increasing crop yields is the use of bacterial formulations based on bacteria referred to as Plant-Growth-Promoting Rhizobacteria (PGPR) [
4,
5]. PGPR stimulate plant growth and development through a variety of mechanisms directly affecting plant growth, including phytohormone production [
6,
7,
8], ACC deaminase activity, biological fixation of atmospheric nitrogen (BNF), and mineral solubilization [
9,
10,
11]. Particularly promising PGPR include bacteria of the genus
Bacillus. In addition to the rhizosphere, they can also reside on the surfaces of above-ground plant organs and in plant tissues [
12,
13].
Bacillus spp. promote plant growth by producing phytohormones, siderophores, lipopeptides, and enzymes [
6,
9,
14]. They also affect plant homeostasis by regulating the ratio of antioxidant enzymes, including under drought stress [
10]. Most research reports the positive effects of
Bacillus spp. on the growth and productivity of common wheat [
4,
15,
16]. According to research conducted by many authors, bacteria of the genus
Pseudomonas are (among other things) involved in disease control and promoting plant growth [
17], and those of the genus
Azotobacter positively affect the seed germination, root length, and shoots of crop plants through the synthesis of phytohormones and BNF [
18]. All the above facets are also important from the point of view of agricultural practices. In addition, the research by Naseri et al. [
19] demonstrated a positive effect of inoculation with
Azotobacter and
Pseudomonas putida bacteria. After the application of these bacteria, the highest grain yield of spring barley was obtained. The application of PGPR, including
Azotobacter spp. bacteria, has a positive effect not only on grain yield but also on the growth and development of cover crops (CC) and especially on the post-harvest yield of grain crops. The introduction of CCs in organic farming also has positive effects on crop management. One of the benefits of a CC is that it provides environmental and soil protection [
20,
21]. Incorporating CCs into cropping systems is recommended to reduce soil erosion and nutrient losses in agroecosystems [
22,
23]. By capturing nutrients that are present after crop harvesting, CCs can also significantly improve soil conditions for subsequent crops, thereby increasing their yield [
24,
25]. There are significant differences between CCs, which largely depend on the timing of a CC’s sowing and its inclusion in the annual rotation. CCs are grown as a green fertilizer for cereals in organic farming [
21,
26]. The potential benefits of using a CC as a green fertilizer depend on the amount of biomass introduced into the soil and its chemical composition, especially its N content and C: N ratio, which determine the rate of biomass decomposition and the availability of nutrients for successor crops [
27,
28]. The beneficial effect of green fertilizer made from legumes in organic cereal cultivation was reported in research conducted by Pickoff et al. [
29]. This benefit was also confirmed by the research conducted by Moyo et al. [
30] on the application of a red clover CC and its mixture with perennial ryegrass and crabapple as a green fertilizer for winter wheat.
The objective of the field research reported herein was to evaluate the effect of CC cultivation and the application of bacterial consortia on soil microorganisms and CC fresh weight yield, in addition to the subsequent effects of CCs applied simultaneously with bacterial consortia and the application of bacterial consortia during the growing season of spelt wheat on spelt wheat yield.
3. Results
The B/A ratios determined after harvesting spring barley were significantly differentiated by the experimental factors researched and their interaction (
Table 2).
The highest B/A ratio was found after the application of bacterial consortium 2; this value was significantly lower after the application of bacterial consortium 1. The lowest B/A was found for control objects. The forecrop of a CC also significantly differentiated the B/A ratio. The post-harvest soil layer of spring barley grown with a CC was higher compared to that of the control objects. A bacterial consortia × CC interaction was also found, with the result being that after the application of bacterial consortium 2, the B/A ratio was significantly higher on the objects with a CC compared to that for the control objects. The B/A ratio for the objects following the application of bacterial consortium 1 in combination with a CC of red clover as well as a mixture of red clover and Italian ryegrass was significantly the highest. On the objects where Italian ryegrass constituted the CC, the B/A ratio was similar to that induced by the other CCs and did not differ significantly from that recorded on the control objects. On the other hand, on the control objects to which bacterial consortia had not been applied, the B/A ratios in the variants without a CC and to which all CCs had been applied were the same.
The weather conditions of the growing season also significantly differentiated the B/A ratio in the soil after harvesting spring barley (
Figure 1).
The highest B/A ratio was recorded in the wet season in 2020, and it was significantly lower in 2019 and 2021, which had less precipitation. An interaction between weather conditions and bacterial consortia was observed, showing that in both the dry spell in 2019 and the wet period in 2020, the highest B/A ratio was recorded on sites for which bacterial consortium 2 had been applied; this value was lower after the application of bacterial consortium 1, and it was the lowest on control objects without bacterial consortia. In 2021, on the other hand, the lowest B/A ratio was recorded on sites after inoculation with bacterial consortium 2, and it was significantly lower after application of bacterial consortium 1, as well as on control objects for which bacterial consortia had not been applied.
An interaction between growing season weather conditions and CC was also observed (
Figure 2).
In the dry year, 2019, the highest B/A ratio was recorded on objects with a CC of Italian ryegrass; this value was significantly lower when the CC was red clover. The B/A ratio on objects with a CC consisting of the Italian ryegrass mixture was at the same level as those with a CC of red clover but was not significantly different from the value recorded on control objects without a CC. In the moist year, 2020, the highest B/A ratio was recorded on the objects for which the CC was a mixture of red clover and Italian ryegrass; this value was significantly lower with a CC of red clover followed by a CC of Italian ryegrass, and it was the lowest on the control objects. In 2021, a year with less precipitation, the B/A ratio on the objects with a CC was significantly higher than that on the control objects.
Statistical analysis demonstrated that there was a significant effect of the researched experimental factors and their interaction on the number of fungi in the soil after harvesting spring barley (
Table 3).
The highest number of fungi was recorded after the application of bacterial consortium 2; this value was significantly lower after the application of consortium 1, and it was the lowest on the control objects. The increase in fungal abundance compared to the control objects was 34% for bacterial consortium 1 and 53% for bacterial consortium 2. The CC also significantly differentiated the abundance of fungi in the soil after harvesting spring barley. The highest number of fungi was recorded on the objects for which the CC was a mixture of red clover and Italian ryegrass; this quantity was significantly lower on the object with a CC of Italian ryegrass, followed by that with a CC of red clover, and it was the lowest on the control objects without a CC. There was an increase in the number of fungi by 25% for red clover CC, by 46% for the Italian ryegrass CC, and by 66% for the mixture compared to the control objects. An interaction was observed wherein on the objects on which bacterial consortium 2 was applied, the highest number of fungi was recorded with the CC consisting of a mixture of red clover and Italian ryegrass and the CC of Italian ryegrass; this value was significantly lower with the red clover CC, and it was the lowest on the control objects without a CC. On the objects for which bacterial consortium 1 was applied, the highest number of fungi was revealed with a CC consisting of a mixture of red clover and Italian ryegrass; this value was significantly lower with a CC of Italian ryegrass. On objects with a CC of red clover, the number of fungi in the soil was similar to those with a CC of Italian ryegrass and did not differ significantly from the number recorded on the control objects without a CC. On the other hand, on the control objects to which bacterial consortia had not been applied, the highest number of fungi in the soil was recorded for the CC consisting of a mixture of red clover and Italian ryegrass and for the CC of perennial ryegrass. The number of fungi in the combination with the CC of red clover was similar to that on other combinations with a CC and did not differ significantly from that recorded on the control object without a CC.
The CC fresh weight yield was significantly differentiated by the application of bacterial consortia and the type of CC (
Table 4).
The highest CC yield was obtained after the application of bacterial consortium 2. It was significantly lower when bacterial consortium 1 was applied, and it was the lowest on the objects to which no bacterial consortia were applied. Concerning the control objects, the application of bacterial consortium 1 resulted in an average CC yield that was 23% higher, while it was 60% higher with bacterial consortium 2 inoculation. The CC variants researched also significantly differentiated with respect to fresh biomass yield. The highest CC yield was obtained from a mixture of red clover and Italian ryegrass; a lower yield was obtained from red clover, and the lowest was obtained from Italian ryegrass. An interaction between the analysed factors was also observed, showing that with the application of the Italian ryegrass of the CC variant researched, the highest yield was obtained after the application of bacterial consortium 2; this value was significantly lower after the application of consortium 1, and it was the lowest on the control objects to which bacterial consortia were not applied.
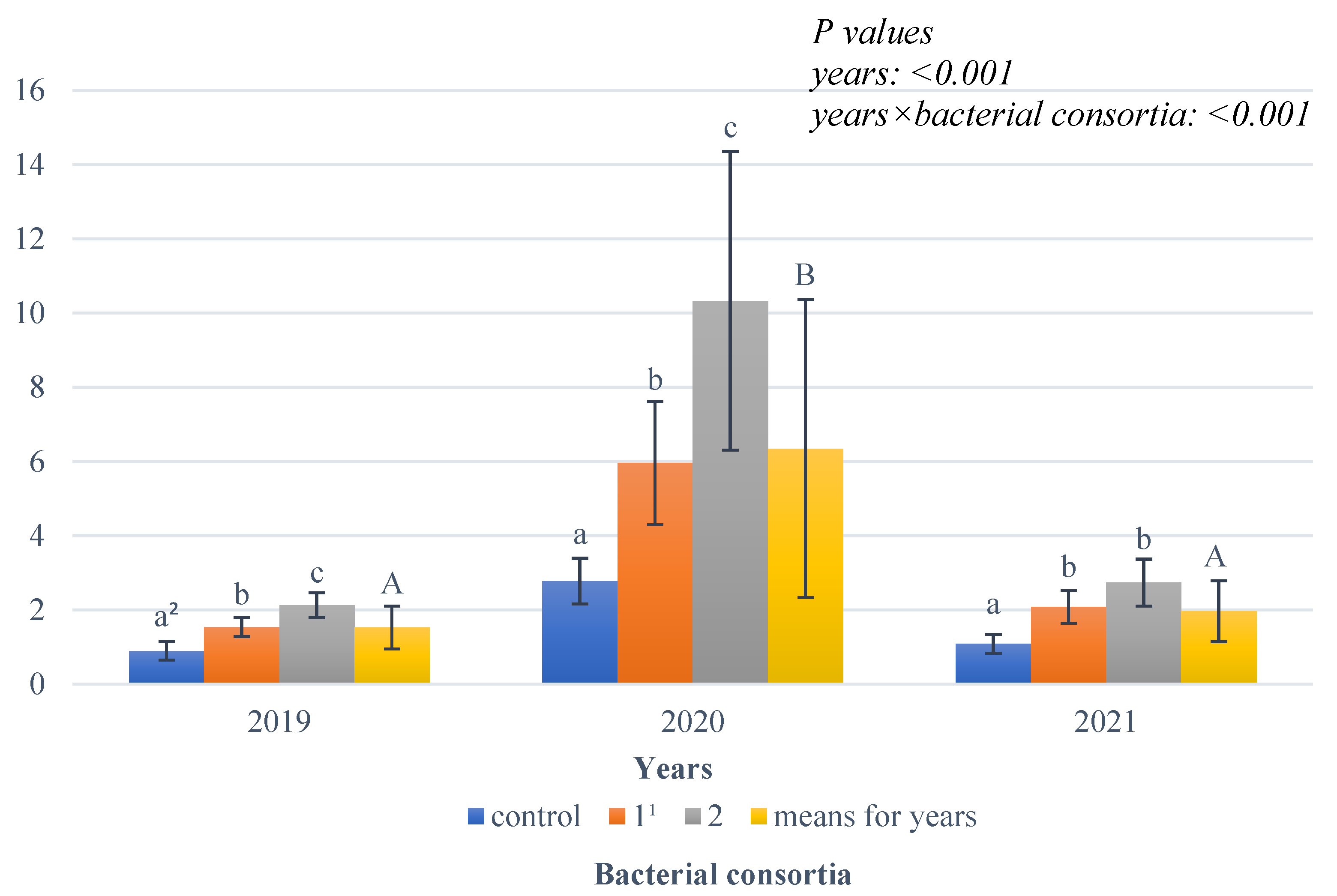
Statistical analysis demonstrated a significant effect of growing season conditions on CC yield (
Figure 3).
The highest fresh CC biomass yield was recorded in 2021, the favourable year; this yield was significantly lower in 2020, and it was the lowest in the dry year, 2019. An interaction of weather conditions with the analysed bacterial consortia was observed, with the result showing that in all the years studied, the highest CC biomass yield was obtained when bacterial consortium 2 was used, while a lower yield was obtained when bacterial consortium 1 was used, and the lowest was obtained when no bacterial consortia were used.
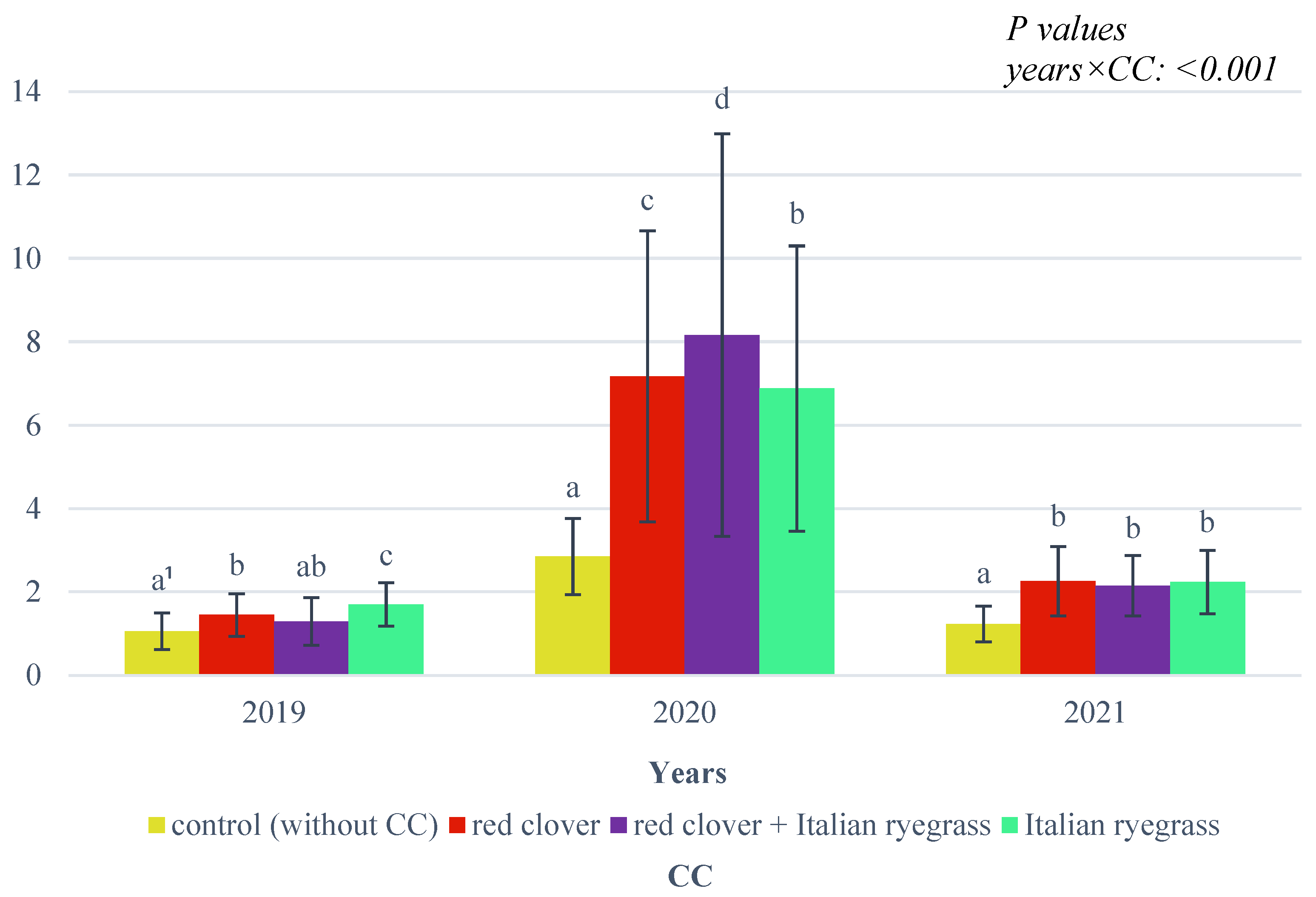
An interaction of weather conditions with the researched CC combinations with respect to fresh biomass yield was also observed (
Figure 4).
In all the years researched, the highest biomass yield was obtained from a mixture of red clover and Italian ryegrass; a significantly lower yield was obtained from red clover, and the lowest was obtained from Italian ryegrass.
Spelt wheat ear yield was significantly differentiated by the application of bacterial consortia and forecrop¬¬s (CCs) and their interaction (
Table 5).
The highest spelt wheat ear yield was obtained after the application of bacterial consortium 2; this value was significantly lower after the application of bacterial consortium 1, and it was the lowest on control objects. For the control objects, the average yield of ears after the application of bacterial consortium 1 was 17% higher, while that of bacterial consortium 2 was 51% higher. The forecrops consisting of a CC also significantly differentiated spelt wheat ear yield. The highest spelt wheat ear yield was recorded after ploughing the entire biomass consisting of a mixture of red clover and Italian ryegrass; this value was significantly lower after red clover followed by Italian ryegrass, and it was the lowest on control objects without a CC. In this case, spelt wheat was grown in the stand after spring barley. Compared to the control objects, the CC ploughing of a mixture of red clover and Italian ryegrass increased spelt wheat ear yield by 30%, that of red clover by 22%, and that of Italian ryegrass by 8%. An interaction between the analysed factors of the experiment was also observed, showing that when using Italian ryegrass for a CC, the highest ear yield was recorded after the application of bacterial consortium 2; it was lower after the application of bacterial consortium 1, and it was the lowest on the objects where no bacterial consortia were applied.
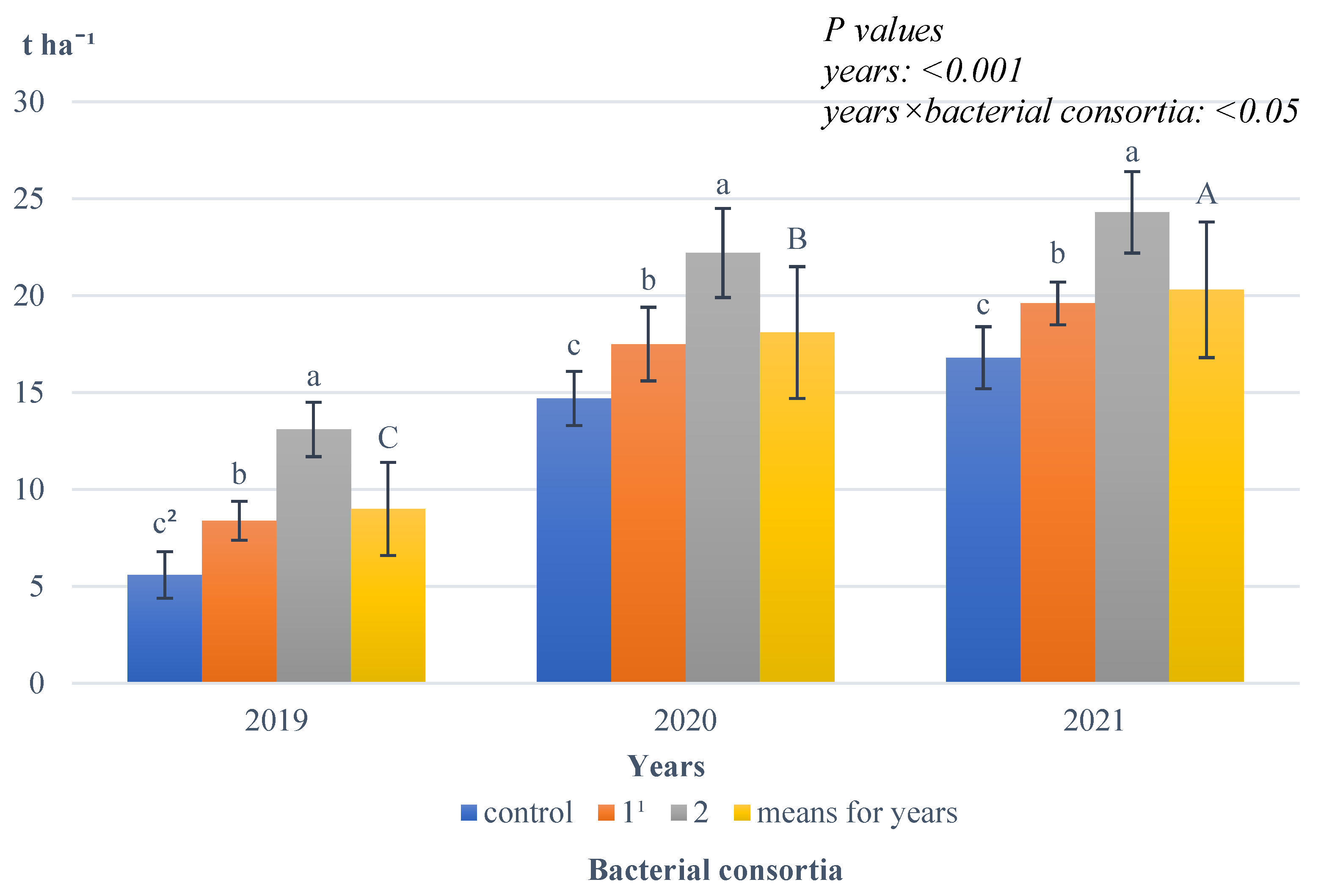
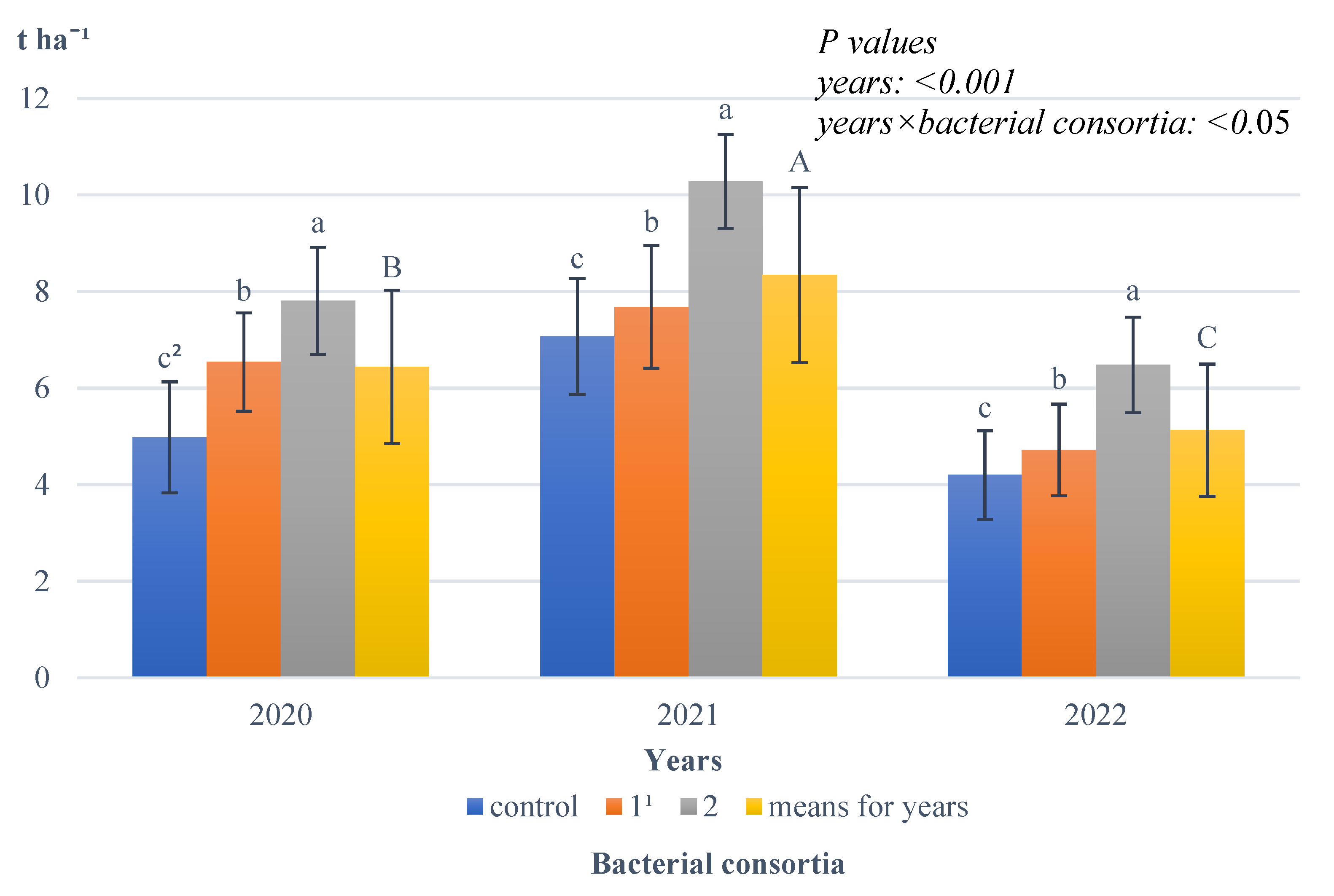
Growing season conditions also significantly affected spelt wheat ear yield (
Figure 5).
The highest ear yield was harvested in the favourable year, 2021; this value was significantly lower in 2020, and it was the lowest in 2022, the year with the least precipitation. An interaction between the years studied and the bacterial consortia used was also found, with the results showing that in each year researched, the highest yield of spelt wheat ears was obtained after the application of bacterial consortium 2; this value was significantly lower after the application of bacterial consortium 1 and the lowest on objects where no bacterial consortia were applied.
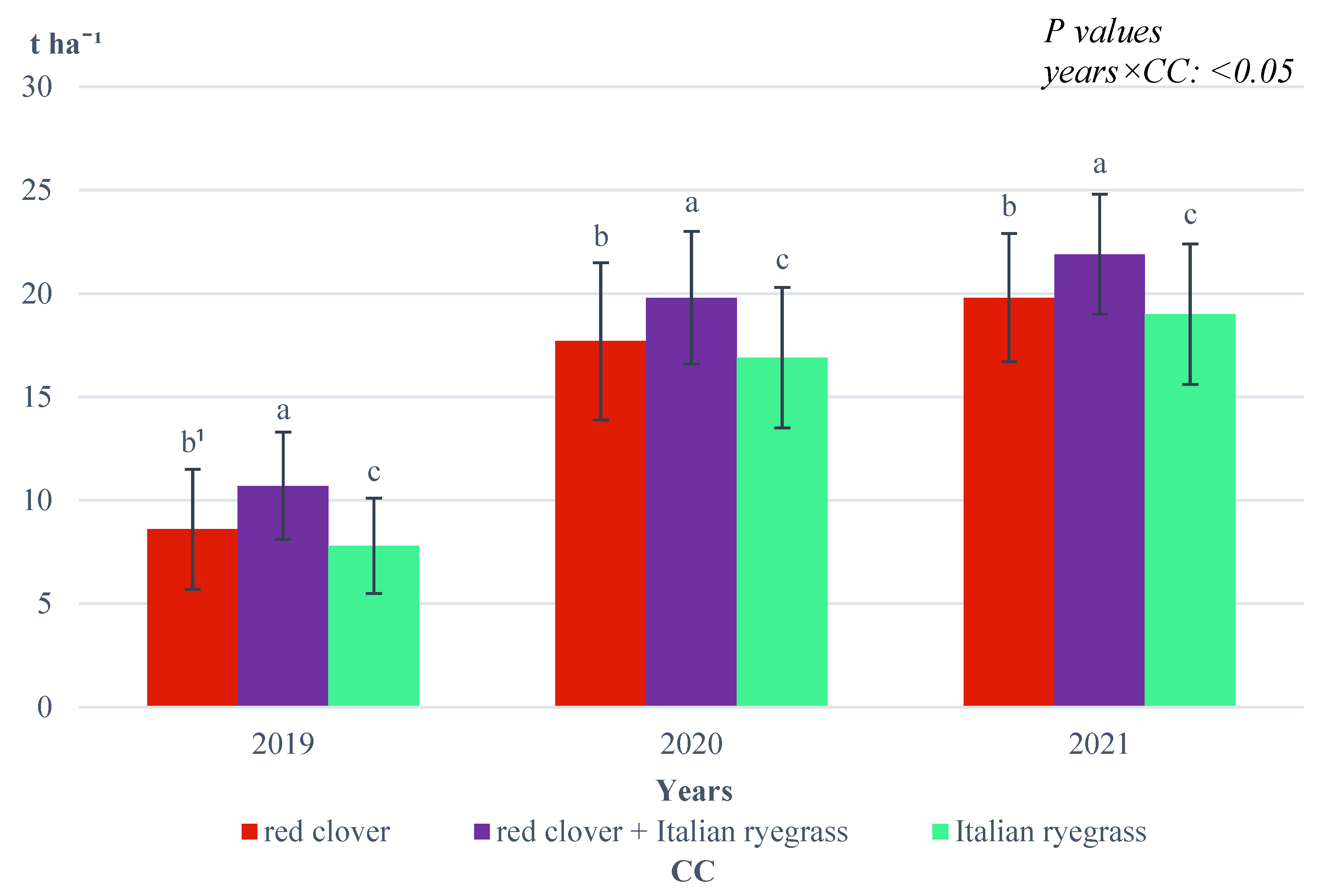
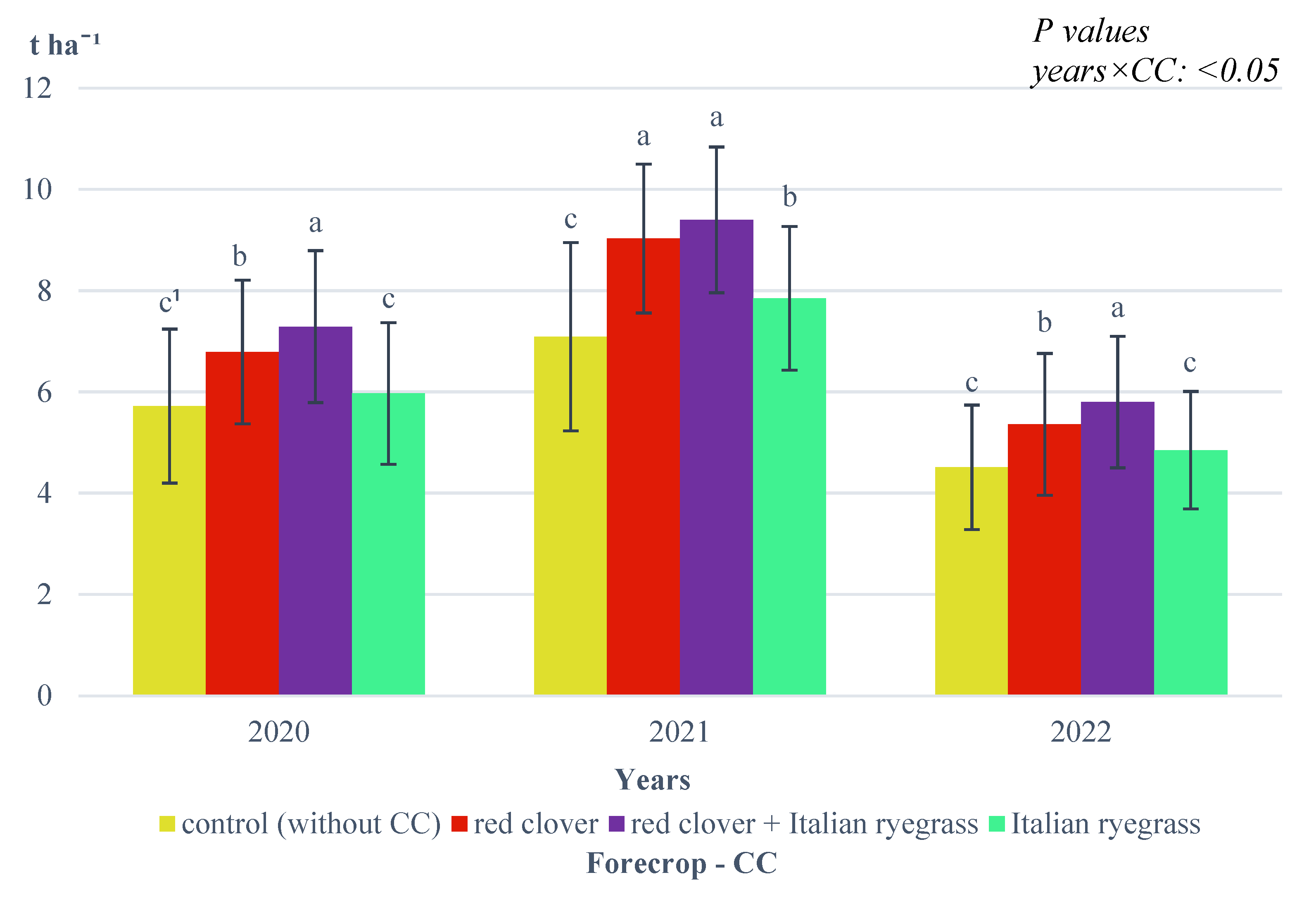
The interaction between growing season conditions and the forecrop of CC was also found (
Figure 6).
In 2020 and 2022, the highest spelt wheat ear yield was recorded when wheat was grown in a stand after ploughing a mixture of red clover and Italian ryegrass and in 2021 also after ploughing red clover. In contrast, in 2020 and 2022, the yield of spelt wheat ears after the CC of red clover was applied was significantly lower. In 2021, the ear yield of spelt wheat grown in a stand after the CC of Italian ryegrass was applied was significantly higher than that for the control, where spelt wheat was grown in objects immediately after spring barley. In 2020 and 2022 on these objects, the spelt wheat ear yields were at the same level, and they were the lowest.
Spelt wheat grain yield was significantly differentiated by the analysed factors of the experiment and their interaction (
Table 6).
The highest grain yield of spelt wheat was recorded after the application of bacterial consortium 2. It was significantly lower after the application of bacterial consortium 1, and it was the lowest on objects for which no bacterial consortium was applied. With respect to the control objects, the application of bacterial consortium 1 resulted in a 26% increase in grain yield, while bacterial consortium 1 increased grain yield by 69%. The forecrops that were CCs also significantly differentiated the grain yield of spelt wheat. The highest grain yield was obtained when spelt wheat was grown in a stand after a CC consisting of a mixture of red clover and Italian ryegrass was applied; this value was significantly lower after a CC of red clover was applied, followed by that after a CC of Italian ryegrass was applied. Significantly, the lowest grain yield was obtained for the objects that were not treated with a CC. Compared to these objects, cultivation after the application of a CC th was a mixture increased grain yield by 38%, with red clover increasing this value by 27% and Italian ryegrass increasing it by 10%. An interaction was also found, showing that when Italian ryegrass was the forecrop, the highest grain yield of spelt wheat was obtained after the application of bacterial consortium 2; this value was significantly lower after bacterial consortium 1 was applied, and it was the lowest on the objects to which no bacterial consortiums were applied.
Pearson correlation analysis demonstrated a significant relationship between the analysed characteristics (
Table 7). A highly significant relationship was found between the number of fungi, the B/A ratio, and the yield of spelt wheat obtained. A significant relationship was also found between CC yield with respect to the number of fungi and B/A ratio. Similarly, a significant relationship was found for CC yield and the subsequent spelt wheat yield obtained.
4. Discussion
The implementation of soil revitalization strategies, especially in organic agriculture, can include the use of a CC. This has become a worldwide phenomenon, especially in light of precipitation deficiencies during the crop growing season [
35,
36]. Also, in our study, the application of a CC to a spring barley crop increased the B/A ratio and the number of fungi, which may be indicative of increased soil moisture. A mixture of red clover with Italian ryegrass and Italian ryegrass alone proved valuable. Increased soil microbial activity was noted under these conditions. CCs cover soil, causing a reduction in water evaporation and limiting access to sunlight, thereby reducing soil temperature [
37]. In addition, according to Gaudin et al. [
38], CCs help maintain high levels of organic matter and biological activity in the soil on which they are applied. Muhammad et al. [
39] reported an increase in the number of fungi and bacteria after the application of CCs to a crop. Therefore, assessing microbial succession in the process of plant growth and development can contribute to a better understanding of the dynamics of soil ecological restoration. Housman et al. [
40] and Scavo et al. [
41] reported increases in the enzymatic activity and microbial biomass of CC mixtures compared to single-species green fertilizer. The number of fungi and bacteria also increases, but fungi thrive better in grass CCs, while the survival conditions of bacteria and actinobacteria are promoted in the N-rich environment of legume CCs [
39,
41,
42]. This is consistent with the results of our research. The population of soil microorganisms was also influenced by the use of bacterial consortia. Analogous to the study conducted by Angelina et al. [
43], an increase in microorganisms was noted after the application of PGPR. Nunan et al. [
44] linked increased microbial communities to increased nutrient availability under PGPR application. The application of PGPR during cultivation also causes an increase in the number of fungi in the soil [
45]. In our study, higher numbers of fungi were recorded as a result of the application of consortium 2 compared to those for consortium 1. This could be due to the association between
Bacillus species and some fungi strains that occurs through the synthesis of chitinases, which are hydrolytic enzymes that break down glycosidic bonds in chitin, a component of fungi cell walls, or through the production of antifungal lipopeptides [
46]. Therefore, the introduction of
Azotobacter additionally into the consortium had a positive effect on the number of fungi in the soil.
In the cited authors’ study, the application of bacterial consortia during the initial period of CC growth and development resulted in a significant increase in biomass yield determined in the fall before ploughing. Also in their study, Mirskaya et al. [
47] demonstrated that the application of PGPR stimulates plant growth and development through various mechanisms: directly through the production of phytohormones [
48], ACC deaminase activity, nitrogen fixation activity [
9,
10], and mineral solubilization [
49] and indirectly through the production of hydrolytic enzymes and siderophores [
6]. The reason for the increase in CC biomass can also be linked to the increased availability of nutrients in the soil, especially nitrogen as a result of the use of microorganisms that exhibit BNF characteristics [
50]. The use of microorganisms in crop cultivation can also contribute to stronger root system development, which translates into more efficient absorption of water and nutrients, resulting in increased yield [
51].
The authors’ research demonstrated that the advantage of using bacterial consortia is an increase in CC biomass even under conditions of precipitation deficiency; these results were recorded in 2019. It should be borne in mind that in the coming years, longer periods of insufficient moisture due to precipitation deficiency and rising temperatures are expected. This trend is linked to climate change, which adversely affects crop production [
52]. Water deficiency, especially during critical growth phases, increases the relative water content of leaves, transpiration rates, the conductivity of stomatal apparatuses, and the rate of cell enlargement and ultimately impairs plant growth [
53]. These difficulties of growing under adverse conditions are remedied by PGPR, as demonstrated by research by Sarma and Saikia [
54], and our research shows that their use in crops increases plant resistance to environmental stresses, including drought. PGPR help plants by changing the morphology of their roots, which results, among other things, in better absorption of water from the soil [
52]. In addition, the introduction of bacterial consortia composed of PGPR can compensate for the effects of drought by improving plant development by increasing the production of proline, amino acids, and soluble sugars, which results in better absorption of water and nutrients from the soil, thereby alleviating high salinity levels in the soil caused by water shortages. The accumulation of proline, which acts as an intercellular substance for osmotic regulation under drought stress conditions, has been widely documented in the literature [
55]. Our research demonstrated that the amount of CC biomass available for green fertilizer is also determined by species composition. The highest fresh matter yield recorded corresponded to red clover and Italian ryegrass mixtures. Also, in a study by Bourgeois et al. [
21], CC crops that included different plant species provided better yields. CCs are an integral part of organic cereal farming given the numerous benefits they provide to the environment and soil [
20,
56,
57,
58]. One application of a CC is for use as green fertilizer. In this group, legumes and their mixtures with grasses, including grasses that are of the least importance in organic farming, are recommended for cultivation. Mixtures of legumes with grasses turn out to be the most reliable in terms of yield. This is because the plants complement each other, as deep-rooted legumes draw water and nutrients from deeper soil layers compared to shallower-rooted grasses [
59]. In addition, grasses can benefit from nitrogen that is bound symbiotically and secreted into the environment by legumes [
60].
Incorporating a CC into cropping systems is recommended to reduce soil erosion and nutrient losses in agroecosystems [
21,
22,
23]. By capturing nutrients remaining in the soil after harvesting crops, CC plants can significantly improve soil conditions for subsequent income crops, thereby increasing their yields [
24,
25]. Green fertilizer from a CC is an important source of nutrients for successively grown crops, especially in organic farming [
21]. This is confirmed by the results of our research, in which ploughing a CC helped improve spelt wheat yields. The potential benefits of CCs for crop productivity are largely determined by the production and composition of CC biomass, among other things, including the N content and C: N ratio, which determine the rate of decomposition of the ploughed biomass and the availability of mineral nutrients for successor crops [
27,
28]. This may explain why the best results for spelt wheat cultivation were obtained after the application of a mixture of red clover and Italian ryegrass. However, the follow-up effect of green fertilizer is determined by the C:N ratio. In the case of legume–grass mixtures, this effect is broader than that in the case of legumes, for which this effect slows down the decomposition process of the ploughed biomass. In such a case, the availability of mineral nutrients extends over time, guaranteeing longer, increased availability to successor plants [
61]. In the case of legumes, the C:N ratio is narrower, so the decomposition of the ploughed biomass is faster, and nutrient losses, including with respect to N, can occur. Nitrates are leached into deeper soil layers because successor plants are unable to take up large amounts of N in the early development stages [
62]. In the experiment in question, the use of bacterial consortia also had a positive effect on the yield of spelt wheat. As reported in the research of many authors [
63,
64,
65], the use of microbial preparations has a positive effect on the growth and development of cereals and, as a result, on their yield. According to Chandra et al. [
66], PGPR directly enhance plant growth through the production of phytohormones such as auxins, cytokinins, gibberellins, and ethylene, which play an important role in root revitalization. Damam et al. [
67] report that phytohormones are the most important growth regulators due to their ability to activate plant metabolism and stimulate defence processes. In addition,
Azotobacter spp. has a beneficial influence on grain yield growth through BNF, the biosynthesis of biologically active substances, the stimulation of rhizosphere microorganisms, the production of phytopathogenic inhibitors, and the alteration of nutrient absorption [
68]. Research by Naseri et al. [
19] also demonstrated the beneficial effect of the combined use of
Azotobacter with
Pseudomonas putida on the yield of spring barley. Our research demonstrated a positive effect of PGPR application on spelt wheat yields, even in the year with the lowest precipitation, during the growing season. Weather predictions suggest that drought problems will occur with an increasing frequency, causing yield reductions [
69]. The use of PGPR in agriculture can offset to some extent the negative effect of reduced water availability for plants. Experiments conducted by Zaheer [
70], Rehman et al. [
71], and Chandra et al. [
72] demonstrated that increased grain yield was one of the effects of drought. Mitigation of the effects of water shortage as a result of PGPR has been linked to an increase in root dry weight and thus an increase in water transport to plants [
73] and the production of phytohormones that make plants resistant to water stress [
48].
5. Conclusions
The use of a spring barley crop with a CC consisting of a mixture of red clover and Italian ryegrass using PGPR resulted in a favourable ratio of total number of bacteria to actinobacteria and number of fungi. Also, the application of PGPR positively affected the amount of CC biomass obtained. These factors induced an increase in the obtained yield of the successor crop, which was spelt wheat. Also, the application of PGPR in spelt wheat cultivation caused an increase in yield, which was also important in years with the lowest precipitation. Thus, introducing a CC as a forecrop and applying PGPR to organic farming show positive effects. The best results were obtained when spelt wheat was grown after the application of a CC consisting of a mixture of red clover and Italian ryegrass and during the growing season when a consortium consisting of Bacillus subtilis, Bacillus amyloliquefaciens, Pseudomonas fluorescens, and Azotobacter chroococcum was used. Such a management technique should be recommended for use in broad agricultural practices on organic farms.
PGPR microorganisms, due to their ability to improve plant vitality through nitrogen fixation, the production of growth hormones, the solubilization of phosphate, the management of plant diseases, and the regeneration of soil health, appear to be among the best options for use as biofertilizers in organic and sustainable crop production. Understanding and manipulating all of these beneficial properties manifested in the PGPR we researched may prove to be a key focus for future crop improvement efforts. According to research, consortia formed from native bacterial species isolated under given climatic conditions and applied under the same conditions are likely to be successful in crop applications. Literature data also indicate that to ensure a maximum benefit is obtained from biopreparations constructed from microbial consortia, the challenge for the research community will be to find compatible partners, i.e., a particular strain of microorganism that will form a good relationship with a particular plant genotype, i.e., a suitable plant–bacteria partnership, as in the case of symbiotic systems, bean plants, and Rhizobium bacteria. In the future, these microorganisms in properly constructed consortia will be able to replace agrochemicals, which have several side effects on sustainable agriculture.

 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}