
Effects of Conventional Non-Biodegradable Film-Derived Microplastics and New Biodegradable Film-Derived Microplastics on Soil Properties and Microorganisms after Entering Sub-Surface Soil


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Soil Sampling and Analysis
2.3. DNA Extraction, Sequencing, and Analysis
2.4. Statistical Analysis
3. Results
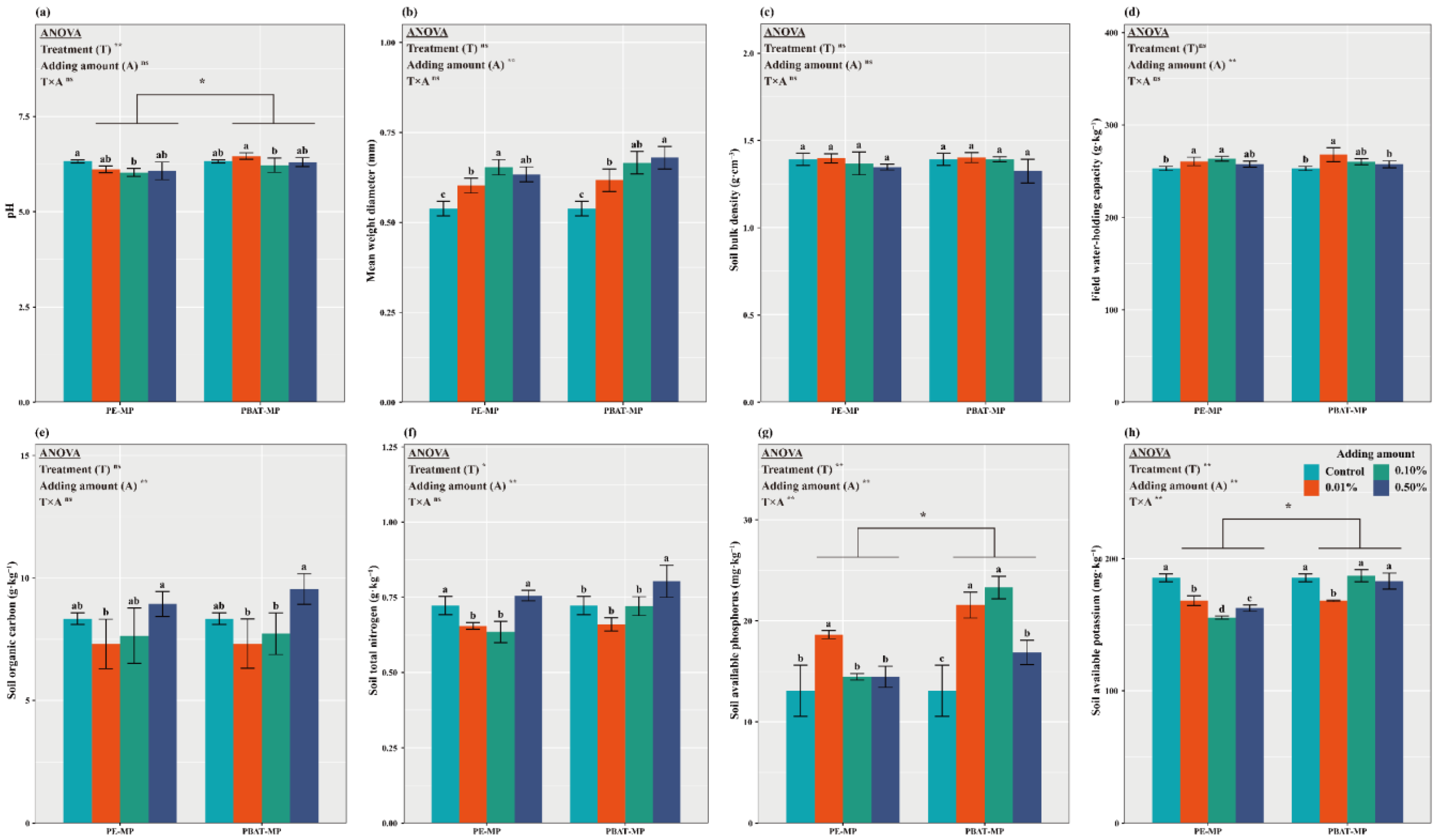
3.1. Impact of MPs on Soil Physicochemical Properties and Nutrient Dynamics
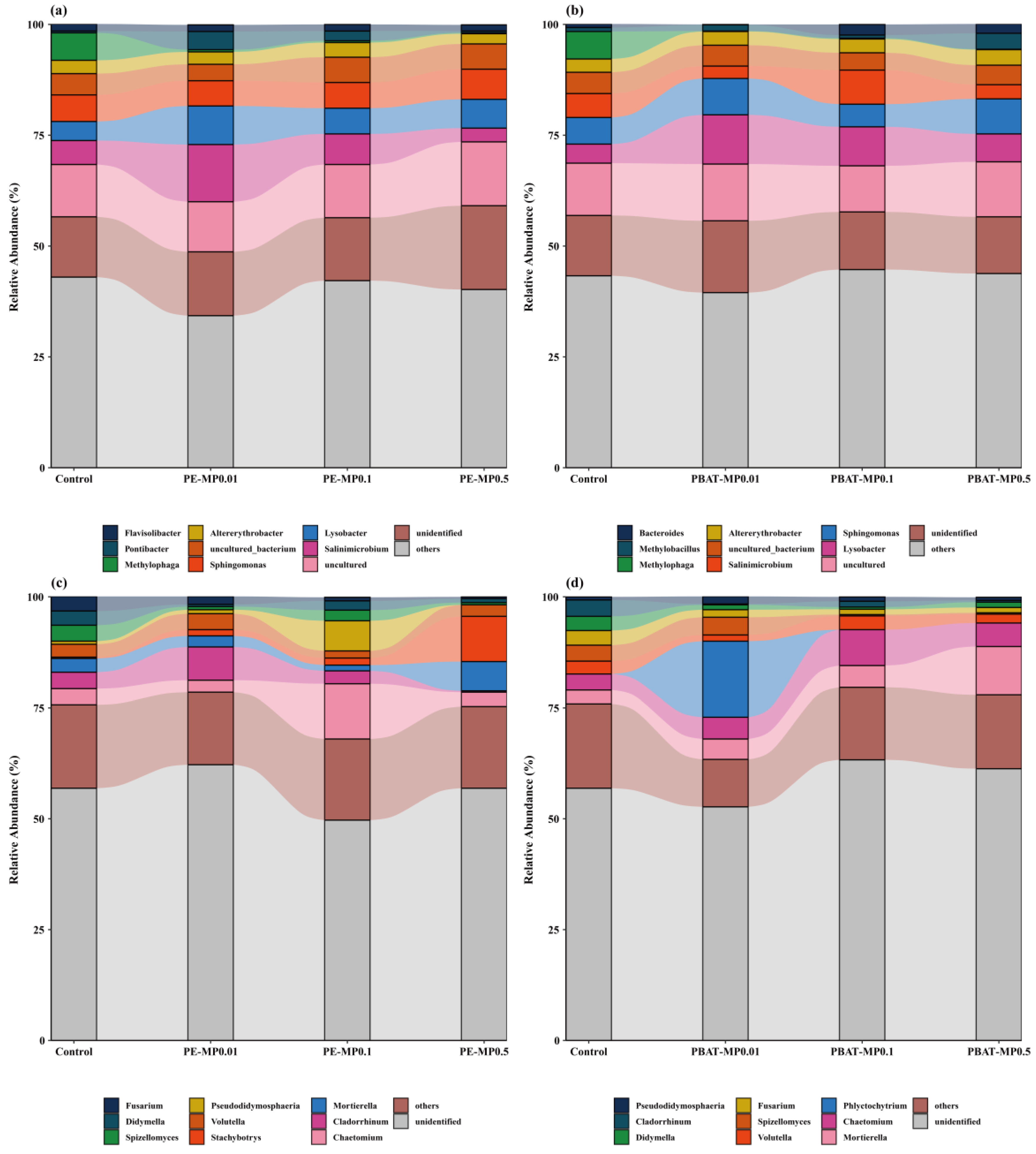
3.2. Impact of MPs on the Community Composition of Bacteria and Fungi
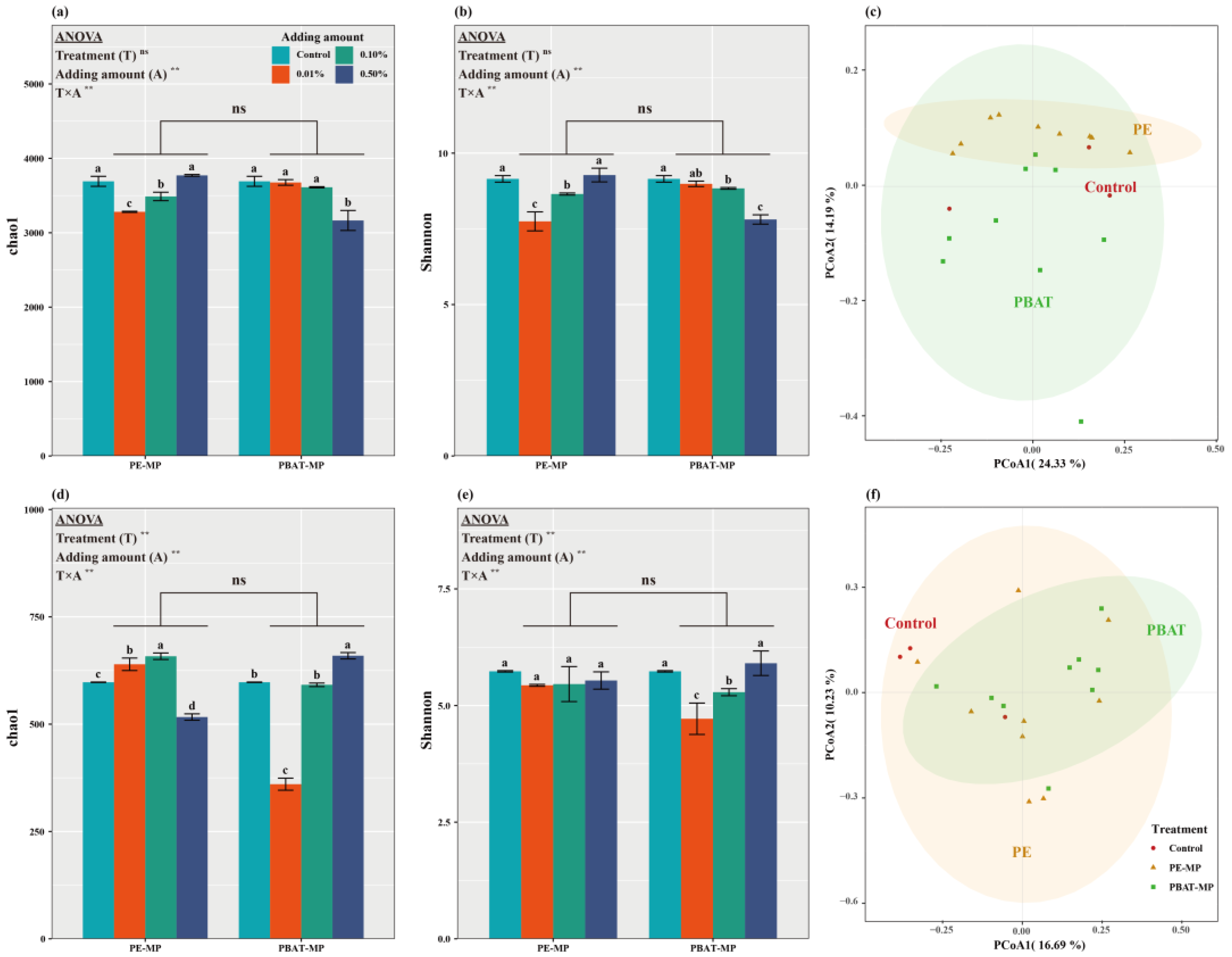
3.3. Impact of MPs on the Diversity of Bacterial and Fungal
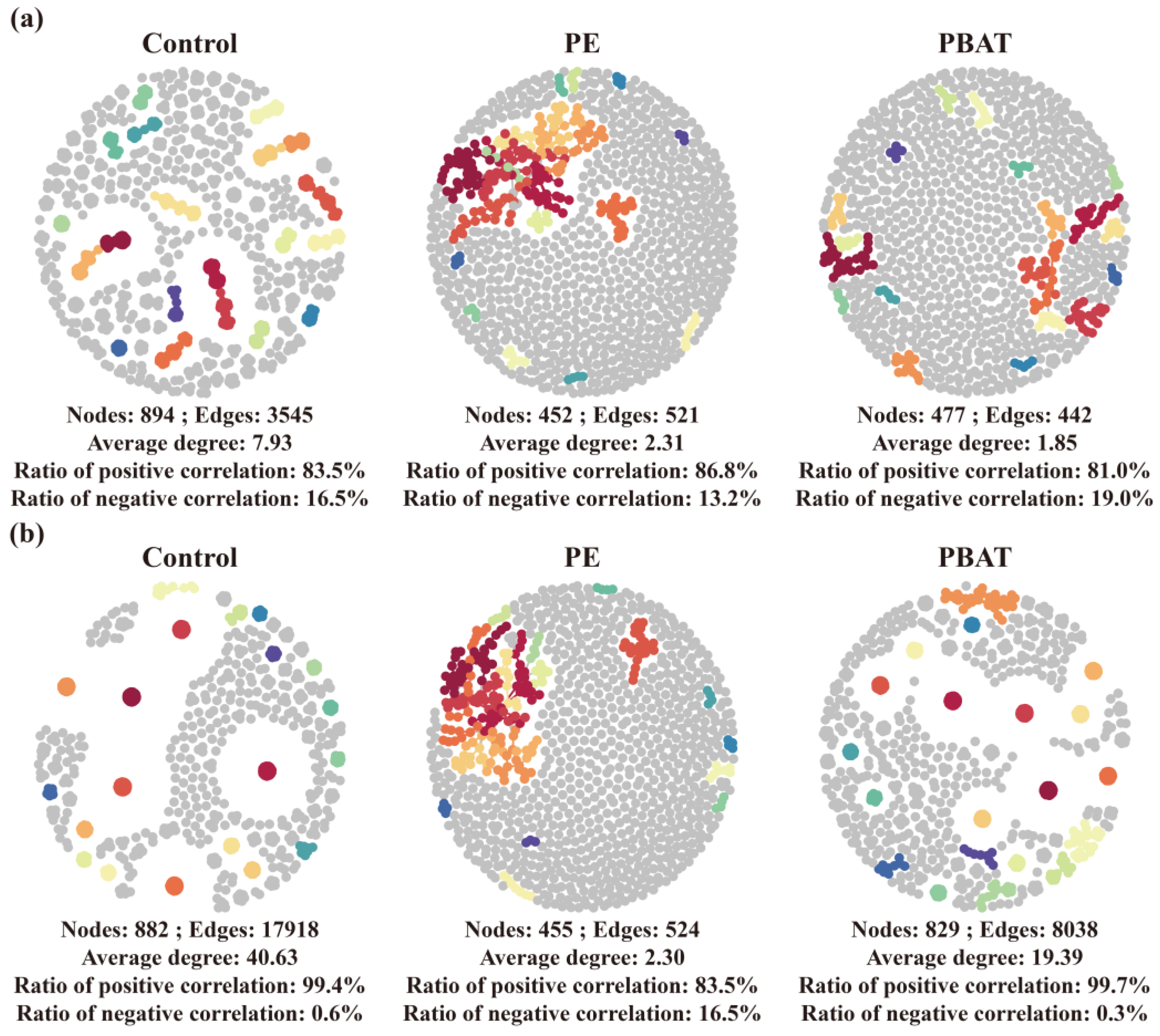
3.4. Impact of MPs on the Co-Occurrence Networks of Soil Bacteria and Fungi
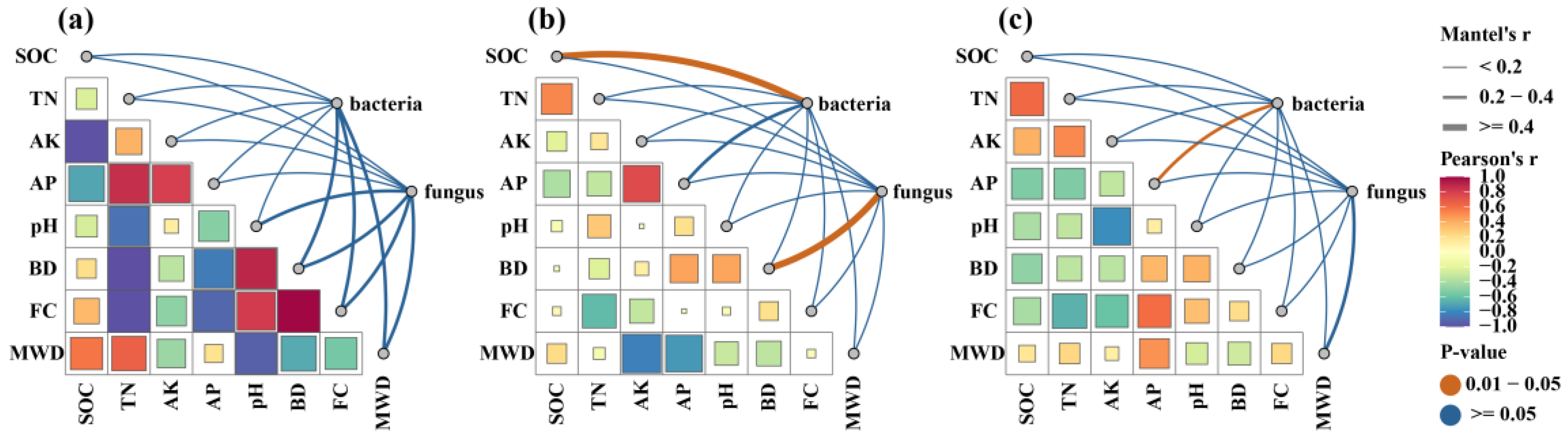
3.5. Relationship between Soil Properties and Microbial Communities
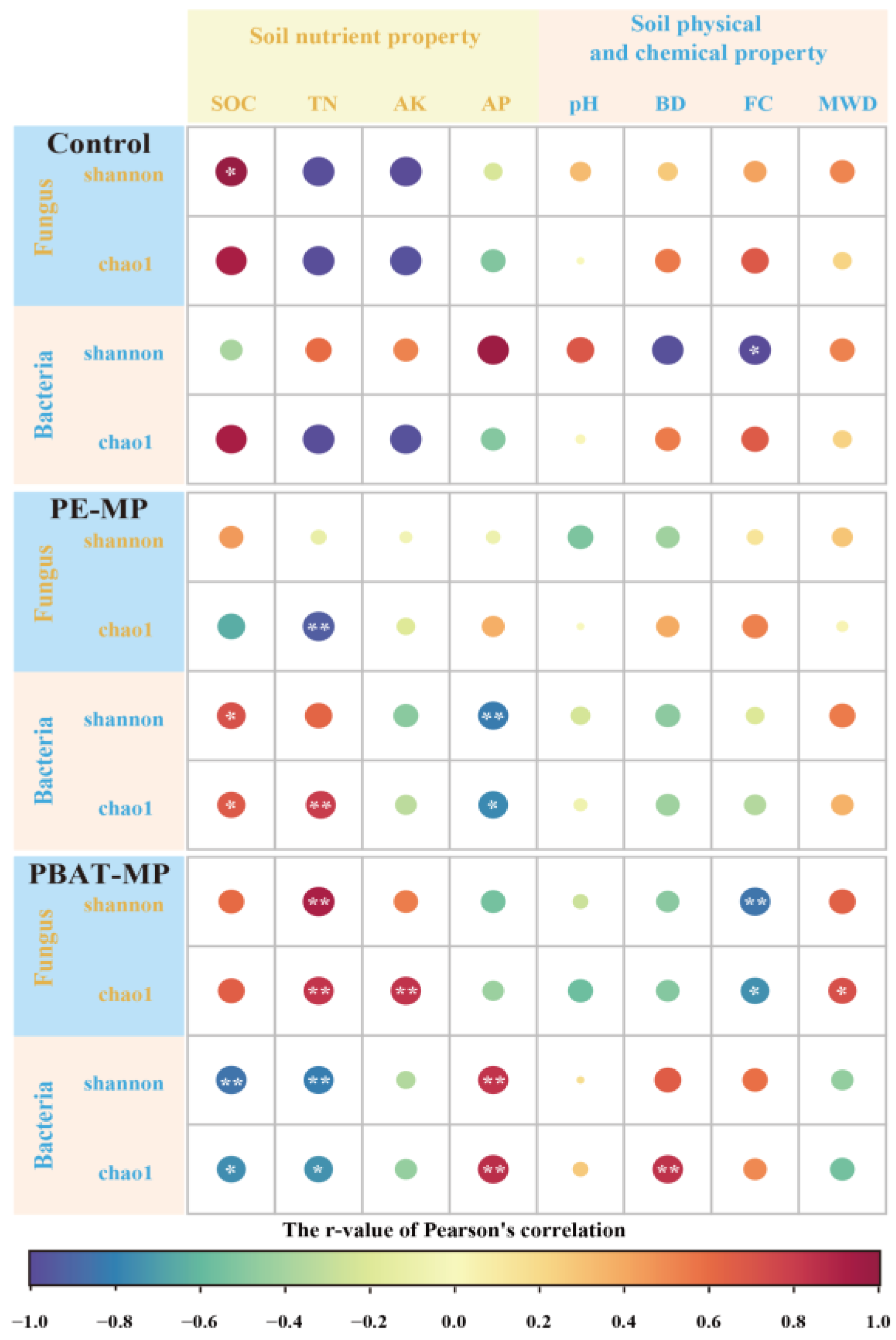
3.6. Relationship between Soil Properties and Microbial Diversity
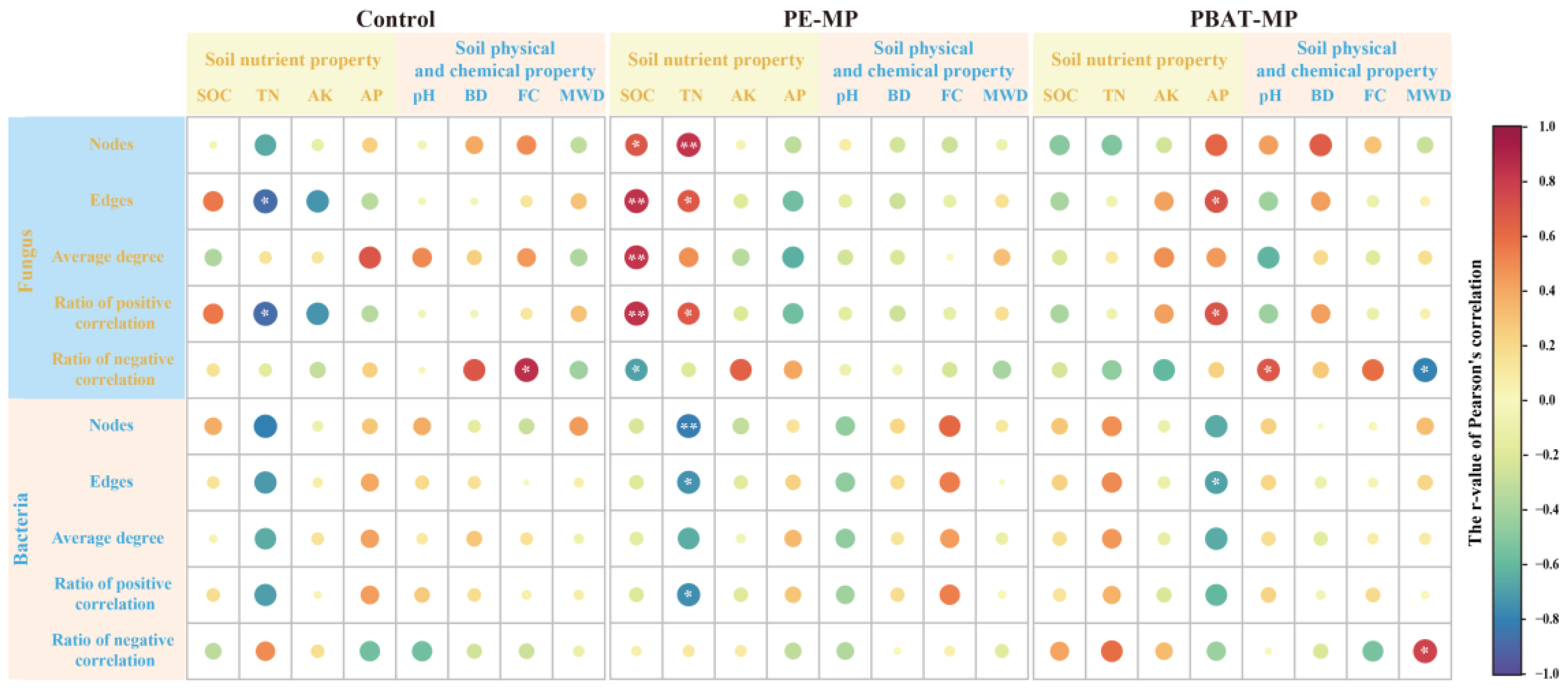
3.7. Relationship between Soil Properties and Network Characteristics
4. Discussion
4.1. Effect of Different MPs on Soil Properties
4.2. Effects of Different MPs on Soil Microbial Community Composition and Diversity
4.3. Effects of MP Addition on Soil Microbial Co-occurrence Networks
4.4. Correlation between Soil Properties and Microbial Properties under MP Addition Conditions
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sun, D.; Li, H.; Wang, E.; He, W.; Hao, W.; Yan, C.; Li, Y.; Mei, X.; Zhang, Y.; Sun, Z. An overview of the use of plastic-film mulching in China to increase crop yield and water-use efficiency. Natl. Sci. Rev. 2020, 7, 1523–1526. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.D.; Chen, L.; Qu, H.C.; Wang, Y.L.; Misselbrook, T.; Jiang, R. Impacts of plastic film mulching on crop yields, soil water, nitrate, and organic carbon in Northwestern China: A meta-analysis. Agric. Water Manag. 2018, 202, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wei, T.; Cai, T.; Ali, S.; Han, Q.F.; Ren, X.L.; Jia, Z.K. Plastic-film mulching for enhanced water-use efficiency and economic returns from maize fields in semiarid China. Front. Plant Sci. 2017, 8, 512. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Liu, Q.; Jia, W.Q.; Yan, C.R.; Wang, J. Agricultural plastic mulching as a source of microplastics in the terrestrial environment. Environ. Pollut. 2020, 260, 114096. [Google Scholar] [CrossRef] [PubMed]
- Liu, E.K.; He, W.Q.; Yan, C.R. ‘White revolution’ to ‘white pollution’- agricultural plastic film mulch in China. Environ. Res. Lett. 2014, 9, 091001. [Google Scholar] [CrossRef]
- Qi, Y.L.; Beriot, N.; Gort, G.; Lwanga, E.H.; Gooren, H.; Yang, X.M.; Geissen, V. Impact of plastic mulch film debris on soil physicochemical and hydrological properties. Environ. Pollut. 2020, 266, 115097. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.L.; Yang, X.M.; Pelaez, A.M.; Lwanga, E.H.; Beriot, N.; Gertsen, H.; Garbeva, P.; Geissen, V. Macro- and micro-plastics in soil-plant system: Effects of plastic mulch film residues on wheat (Triticum aestivum) growth. Sci. Total Environ. 2018, 645, 1048–1056. [Google Scholar] [CrossRef]
- Jiang, M.; Wang, B.; Ye, R.; Yu, N.; Xie, Z.; Hua, Y.; Zhou, R.; Tian, B.; Dai, S. Evidence and impacts of nanoplastic accumulation on crop grains. Adv. Sci. 2022, 9, 2202336. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.J.; Huang, X.P.; Xiang, L.; Wang, Y.Z.; Li, Y.W.; Li, H.; Cai, Q.Y.; Mo, C.H.; Wong, M.H. Source, migration and toxicology of microplastics in soil. Environ. Int. 2020, 137, 105263. [Google Scholar] [CrossRef] [PubMed]
- Qi, R.M.; Jones, D.L.; Li, Z.; Liu, Q.; Yan, C.R. Behavior of microplastics and plastic film residues in the soil environment: A critical review. Sci. Total Environ. 2020, 703, 134722. [Google Scholar] [CrossRef] [PubMed]
- Fu, F.; Long, B.B.; Huang, Q.; Li, J.J.; Zhou, W.J.; Yang, C. Integrated effects of residual plastic films on soil-rhizosphere microbe-plant ecosystem. J. Hazard. Mater. 2023, 445, 130420. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Ossowicki, A.; Yergeau, É.; Vigani, G.; Geissen, V.; Garbeva, P. Plastic mulch film residues in agriculture: Impact on soil suppressiveness, plant growth, and microbial communities. FEMS Microbiol. Ecol. 2022, 98, fiac17. [Google Scholar] [CrossRef] [PubMed]
- Boots, B.; Russell, C.W.; Green, D.S. Effects of microplastics in soil ecosystems: Above and below ground. Environ. Sci. Technol. 2019, 53, 11496–11506. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Shi, H.; Li, L.; Li, J.; Jabeen, K.; Kolandhasamy, P. Microplastic pollution in table salts from China. Environ. Sci. Technol. 2015, 49, 13622–13627. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Xu, Y.; Cheng, Z.; Yang, Y.; Guan, Z.; Jiang, L.; Tang, K. The impact of microplastic pollution on ecological environment: A review. Front. Biosci. 2022, 27, 46. [Google Scholar] [CrossRef] [PubMed]
- Gross, R.A.; Kalra, B. Biodegradable polymers for the environment. Science 2002, 297, 803–807. [Google Scholar] [CrossRef]
- Tanunchai, B.; Juncheed, K.; Wahdan, S.F.M.; Guliyev, V.; Udovenko, M.; Lehnert, A.; Alves, E.G.; Glaser, B.; Noll, M.; Buscot, F. Analysis of microbial populations in plastic–soil systems after exposure to high poly (butylene succinate-co-adipate) load using high-resolution molecular technique. Environ. Sci. Eur. 2021, 33, 1–17. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, Y.; Liu, J.L.; Liu, X.W.; Dong, Y.Y.; Huang, X.; Zhen, Z.C.; Lv, J.; He, W.Q. Degradability and properties of PBAT-based biodegradable mulch films in field and their effects on cotton planting. Polymers 2022, 14, 3157. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Min, J.; Xue, T.; Jiang, P.; Liu, X.; Peng, R.; Huang, J.; Qu, Y.; Li, X.; Ma, N. Complete bio-degradation of poly (butylene adipate-co-terephthalate) via engineered cutinases. Nat. Commun. 2023, 14, 1645. [Google Scholar] [CrossRef] [PubMed]
- Sreejata, B.; González, J.; Henderson, K.; Anunciado, M.; Hayes, D.; Debruyn, J. Soil microbial communities associated with biodegradable plastic mulch films. Front. Microbiol. 2020, 11, 587074. [Google Scholar]
- Qiu, Y.; Zhou, S.; Zhang, C.; Chen, L.; Qin, W.; Zhang, Q. Vertical distribution and weathering characteristic of microplastics in soil profile of different land use types. Sci. Total Environ. 2023, 905, 166902. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, X.; Hao, X.; Wang, J.; Zhang, Y. Distribution of low-density microplastics in the mollisol farmlands of northeast China. Sci. Total Environ. 2020, 708, 135091. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hou, P.; Liu, K.; Hayat, K.; Liu, W. Depth distribution of nano-and microplastics and their contribution to carbon storage in Chinese agricultural soils. Sci. Total Environ. 2024, 913, 169709. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xie, Y.; Li, H.; Dong, H.; Li, B.; Guo, Y.; Wang, Y.; Guo, X.; Yin, T.; Liu, X.; et al. Responses of lettuce (Lactuca sativa L.) growth and soil properties to conventional non-biodegradable and new biodegradable microplastics. Environ. Pollut. 2024, 341, 122897. [Google Scholar] [CrossRef] [PubMed]
- de Souza Machado, A.A.; Lau, C.W.; Till, J.; Kloas, W.; Lehmann, A.; Becker, R.; Rillig, M.C. Impacts of microplastics on the soil biophysical environment. Environ. Sci. Technol. 2018, 52, 9656–9665. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, P.; Wang, Y.; Zhang, Y.; Xu, T.; Zhang, Y.; Xi, J.; Hou, L.; Li, L.; Zhang, Z. Negative effects of poly (butylene adipate-co-terephthalate) microplastics on Arabidopsis and its root-associated microbiome. J. Hazard. Mater. 2022, 437, 129294. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.D. Agricultural and Chemistry Analysis of Soil; Agriculture Press: Beijing, China, 2005. [Google Scholar]
- Reinsch, T.G.; Grossman, R.B. A method to predict bulk density of tilled Ap horizons. Soil Tillage Res. 1995, 34, 95–104. [Google Scholar] [CrossRef]
- Hillel, D. Environmental Soil Physics; Academic Press. San Diego, CA, USA, 1998.
- Elliott, E. Aggregate structure and carbon, nitrogen, and phosphorus in native and cultivated soils. Soil Sci. Soc. Am. J. 1986, 50, 627–633. [Google Scholar] [CrossRef]
- Bavel, C. Mean weight-diameter of soil aggregates as a statistical index of aggregation. Proc. Soil Sci. Soc. Am. J. 1950, 14, 20–23. [Google Scholar] [CrossRef]
- Ginestet, C. ggplot2: Elegant Graphics for Data Analysis; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- Oksanen, J.; Kindt, R.; Legendre, P.; O Hara, B.; Stevens, M.H.H.; Oksanen, M.J.; Suggests, M. The vegan package. Community Ecol. 2007, 10, 719. [Google Scholar]
- Wen, T.; Xie, P.; Yang, S.; Niu, G.; Liu, X.; Ding, Z.; Xue, C.; Liu, Y.X.; Shen, Q.; Yuan, J. ggClusterNet: An R package for microbiome network analysis and modularity-based multiple network layouts. iMeta 2022, 1, e32. [Google Scholar] [CrossRef]
- Yang, X.; Song, W.; Yang, X.; Bao, W.; Li, J.; Zhong, S.; Sun, W. Microbial network structure, not plant and microbial community diversity, regulates multifunctionality under increased precipitation in a cold steppe. Front. Microbiol. 2024, 14, 1349747. [Google Scholar] [CrossRef] [PubMed]
- Abad-Valle, P.; Iglesias-Jiménez, E.; álvarez-Ayuso, E. A comparative study on the influence of different organic amendments on trace element mobility and microbial functionality of a polluted mine soil. J. Environ. Manag. 2017, 188, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Palansooriya, K.N.; Sang, M.K.; El-Naggar, A.; Shi, L.; Chang, S.X.; Sung, J.; Zhang, W.; Ok, Y.S. Low-density polyethylene microplastics alter chemical properties and microbial communities in agricultural soil. Sci. Rep. 2023, 13, 16276. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Cui, Q.; Li, Y.; Zhang, K.; Lu, X.; Zhang, Y. Effect of LDPE and biodegradable PBAT primary microplastics on bacterial community after four months of soil incubation. J. Hazard. Mater. 2022, 429, 128353. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.A.; Hasan, F.; Hameed, A.; Ahmed, S. Biological degradation of plastics: A comprehensive review. Biotechnol. Adv. 2008, 26, 246–265. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Gong, J.; Qin, M.; Qin, L.; Fang, S.; Li, J.; Tang, S.; Tan, Z.; Li, X.; Chen, Z. Biodegradable Microplastics Affect the Wheatgrass Traits, Fe Plaque Development Involved in Sb Accumulation, and Microbial Community Functions in Antimony-Contaminated Riparian Wetlands. ACS Sustain. Chem. Eng. 2022, 10, 5847–5858. [Google Scholar] [CrossRef]
- Xi, B.; Wang, B.; Chen, M.; Lee, X.; Zhang, X.; Wang, S.; Yu, Z.; Wu, P. Environmental behaviors and degradation methods of microplastics in different environmental media. Chemosphere 2022, 299, 134354. [Google Scholar] [CrossRef]
- Zhang, X.; Xia, X.; Dai, M.; Cen, J.; Zhou, L.; Xie, J. Microplastic pollution and its relationship with the bacterial community in coastal sediments near Guangdong Province, South China. Sci. Total Environ. 2021, 760, 144091. [Google Scholar] [CrossRef]
- Xiao, S.; Zhang, Y.; Wu, Y.; Li, J.; Dai, W.; Pang, K.; Liu, Y.; Wu, R. Bacterial community succession and the enrichment of antibiotic resistance genes on microplastics in an oyster farm. Mar. Pollut. Bull. 2023, 194, 115402. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Xu, L.; Bai, X.; Zhang, G.; Zhang, M.; Huang, Y. Differential fungal assemblages and functions between the plastisphere of biodegradable and conventional microplastics in farmland. Sci. Total Environ. 2024, 906, 167478. [Google Scholar] [CrossRef] [PubMed]
- Rüthi, J.; Bölsterli, D.; Pardi-Comensoli, L.; Brunner, I.; Frey, B. The “plastisphere” of biodegradable plastics is characterized by specific microbial taxa of alpine and arctic soils. Front. Environ. Sci. 2020, 8, 562263. [Google Scholar] [CrossRef]
- Li, K.; Jia, W.; Xu, L.; Zhang, M.; Huang, Y. The plastisphere of biodegradable and conventional microplastics from residues exhibit distinct microbial structure, network and function in plastic-mulching farmland. J. Hazard. Mater. 2023, 442, 130011. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Cao, M.; Zhang, W.; Chen, X.; Liu, H.; Ning, Z.; Peng, L.; Fan, C.; Wu, D.; Zhang, M. Effect of biodegradable PBAT microplastics on the C and N accumulation of functional organic pools in tropical latosol. Environ. Int. 2024, 183, 108393. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Hu, Z.; Wu, F.; Guo, K.; Gu, F.; Cao, M. The use and recycling of agricultural plastic mulch in China: A review. Sustainability 2023, 15, 15096. [Google Scholar] [CrossRef]
- Kanwal, A.; Zhang, M.; Sharaf, F.; Li, C. Polymer pollution and its solutions with special emphasis on Poly (butylene adipate terephthalate (PBAT)). Polym. Bull. 2022, 79, 9303–9330. [Google Scholar] [CrossRef]
- Li, C.; Li, Z.; Cui, Q.; Hassan, A.; Zhang, K.; Lu, X.; Zhang, Y. Effect of different additions of low-density polyethylene and microplastics polyadipate/butylene terephthalate on soil bacterial community structure. Environ. Sci. Pollut. Res. 2023, 30, 55649–55661. [Google Scholar] [CrossRef]
- Nie, Z.; Wang, L.; Lin, Y.; Xiao, N.; Zhao, J.; Wan, X.; Hu, J. Effects of polylactic acid (PLA) and polybutylene adipate-co-terephthalate (PBAT) biodegradable microplastics on the abundance and diversity of denitrifying and anammox bacteria in freshwater sediment. Environ. Pollut. 2022, 315, 120343. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zhao, X.; Wu, D.; Peng, L.; Fan, C.; Zhang, W.; Li, Q.; Ge, C. Addition of biodegradable microplastics alters the quantity and chemodiversity of dissolved organic matter in latosol. Sci. Total Environ. 2022, 816, 151960. [Google Scholar] [CrossRef] [PubMed]
- Nelson, T.F.; Baumgartner, R.; Jaggi, M.; Bernasconi, S.M.; Battagliarin, G.; Sinkel, C.; Künkel, A.; Kohler, H.E.; Mcneill, K.; Sander, M. Biodegradation of poly (butylene succinate) in soil laboratory incubations assessed by stable carbon isotope labelling. Nat. Commun. 2022, 13, 5691. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Wei, W.; Liu, G.; Zhu, B.; Cui, J.; Yin, T. Effects of Conventional Non-Biodegradable Film-Derived Microplastics and New Biodegradable Film-Derived Microplastics on Soil Properties and Microorganisms after Entering Sub-Surface Soil. Agronomy 2024, 14, 753. https://doi.org/10.3390/agronomy14040753
Liu X, Wei W, Liu G, Zhu B, Cui J, Yin T. Effects of Conventional Non-Biodegradable Film-Derived Microplastics and New Biodegradable Film-Derived Microplastics on Soil Properties and Microorganisms after Entering Sub-Surface Soil. Agronomy. 2024; 14(4):753. https://doi.org/10.3390/agronomy14040753
Chicago/Turabian StyleLiu, Xiaowei, Wenliang Wei, Guocheng Liu, Bo Zhu, Jie Cui, and Tao Yin. 2024. "Effects of Conventional Non-Biodegradable Film-Derived Microplastics and New Biodegradable Film-Derived Microplastics on Soil Properties and Microorganisms after Entering Sub-Surface Soil" Agronomy 14, no. 4: 753. https://doi.org/10.3390/agronomy14040753
APA StyleLiu, X., Wei, W., Liu, G., Zhu, B., Cui, J., & Yin, T. (2024). Effects of Conventional Non-Biodegradable Film-Derived Microplastics and New Biodegradable Film-Derived Microplastics on Soil Properties and Microorganisms after Entering Sub-Surface Soil. Agronomy, 14(4), 753. https://doi.org/10.3390/agronomy14040753