
Positive Correlation of Lodging Resistance and Soybean Yield under the Influence of Uniconazole


Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Field Experimental Design
2.3. Sampling and Measurements
2.3.1. Growth and Physiological Parameter Determination
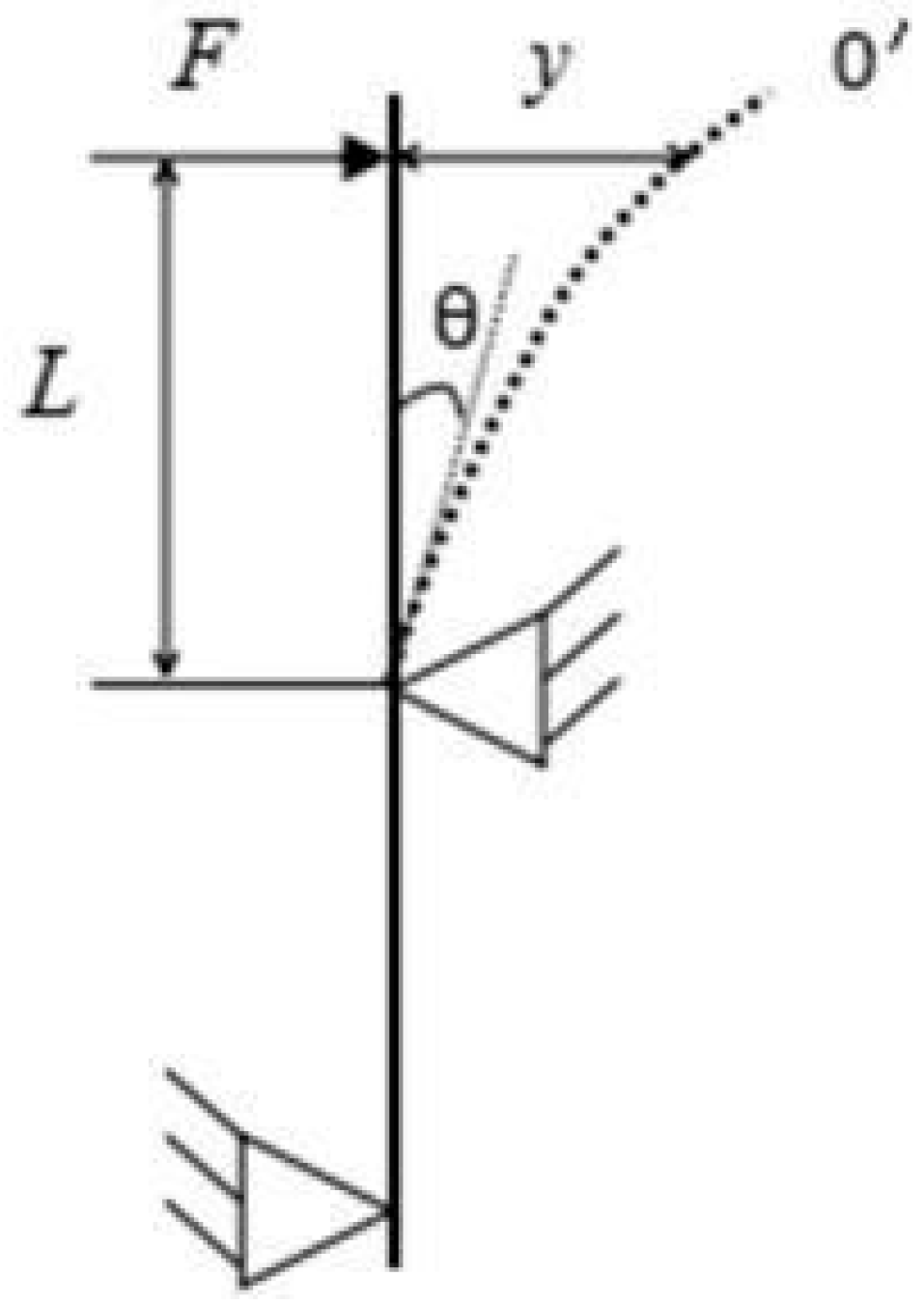
2.3.2. Breaking Force and Stem Lodging Resistance Coefficient Determination
2.3.3. Yield Determination
2.4. Statistical Analysis
3. Results
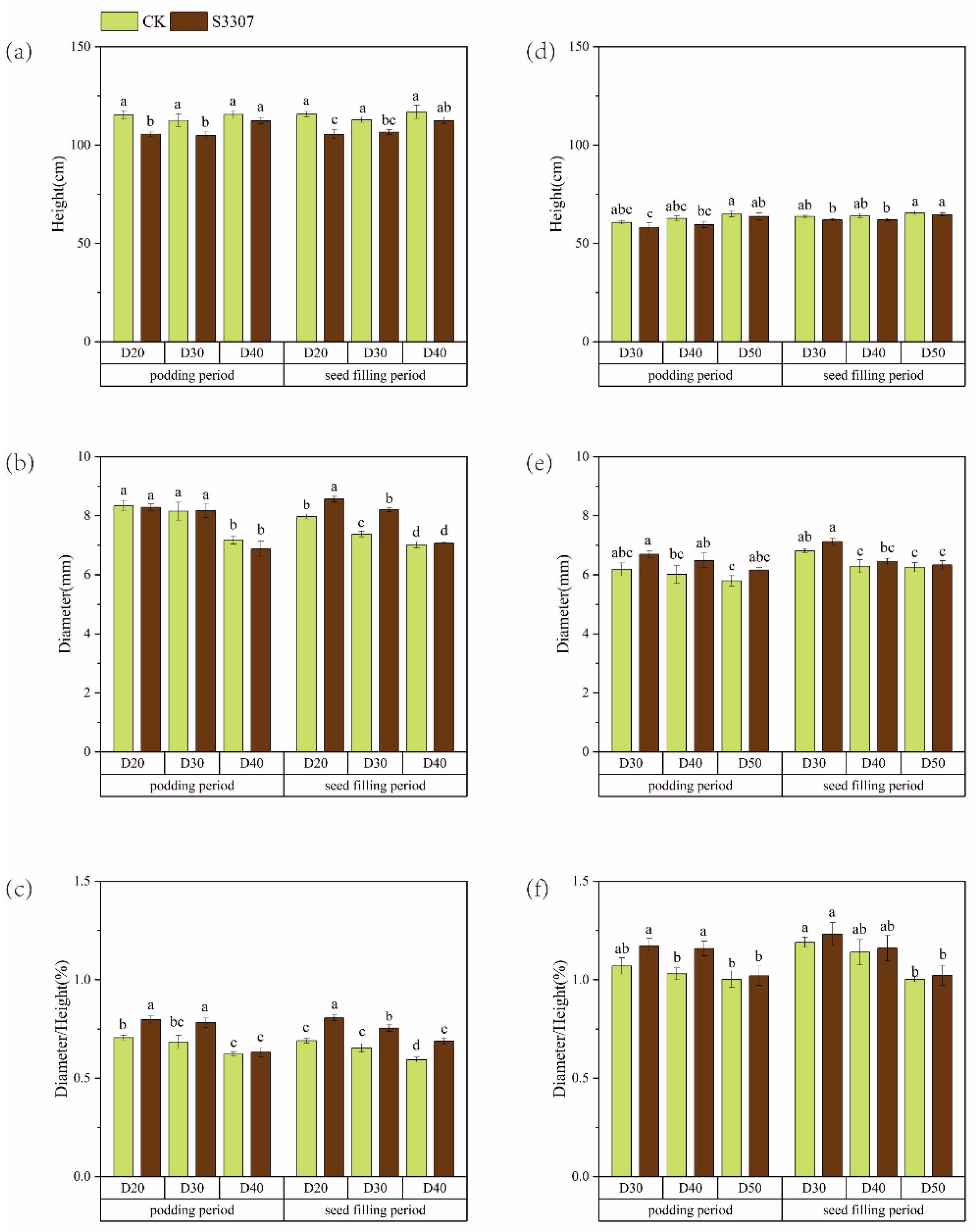
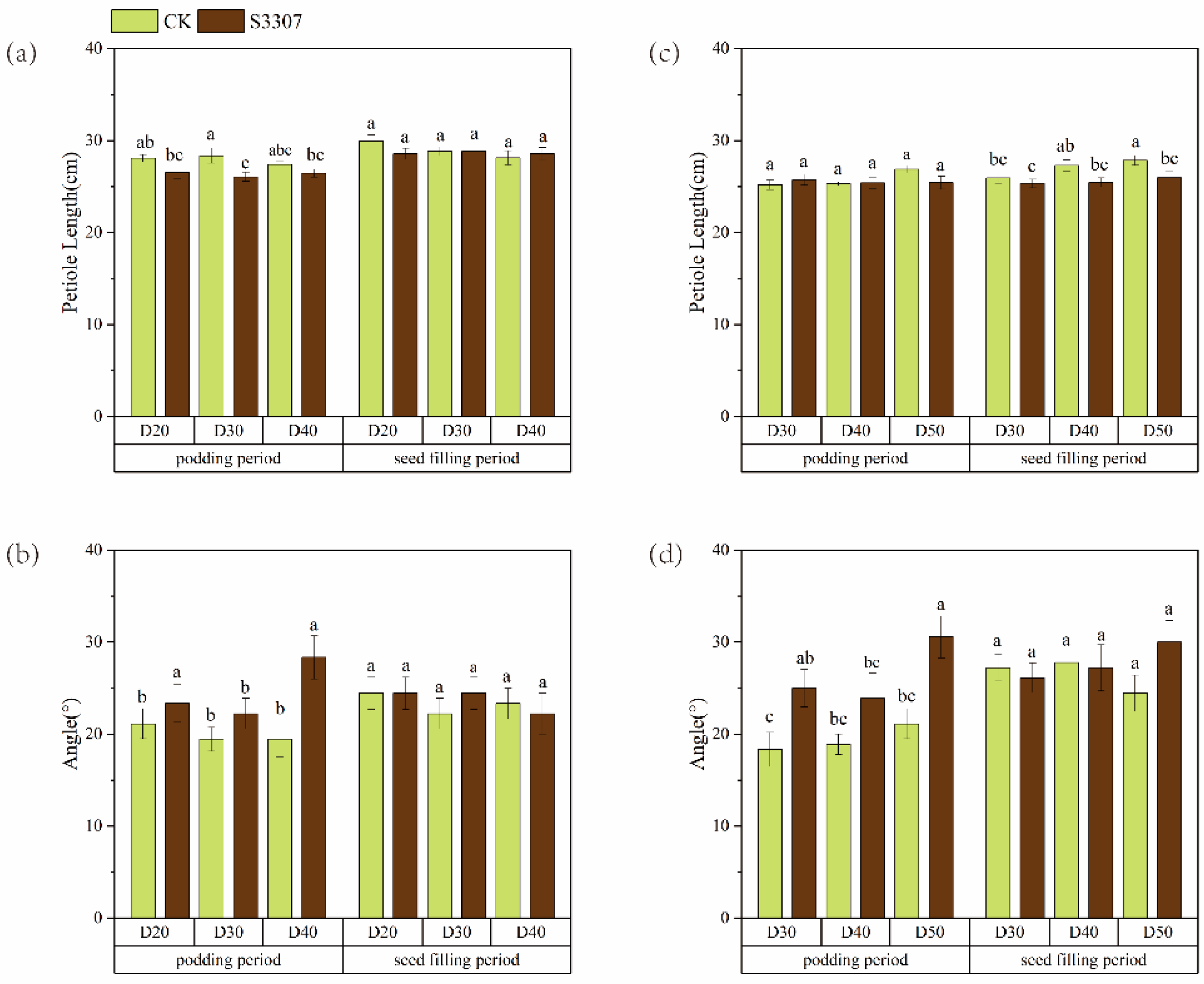
3.1. Morphological Characteristics of Soybean Plants
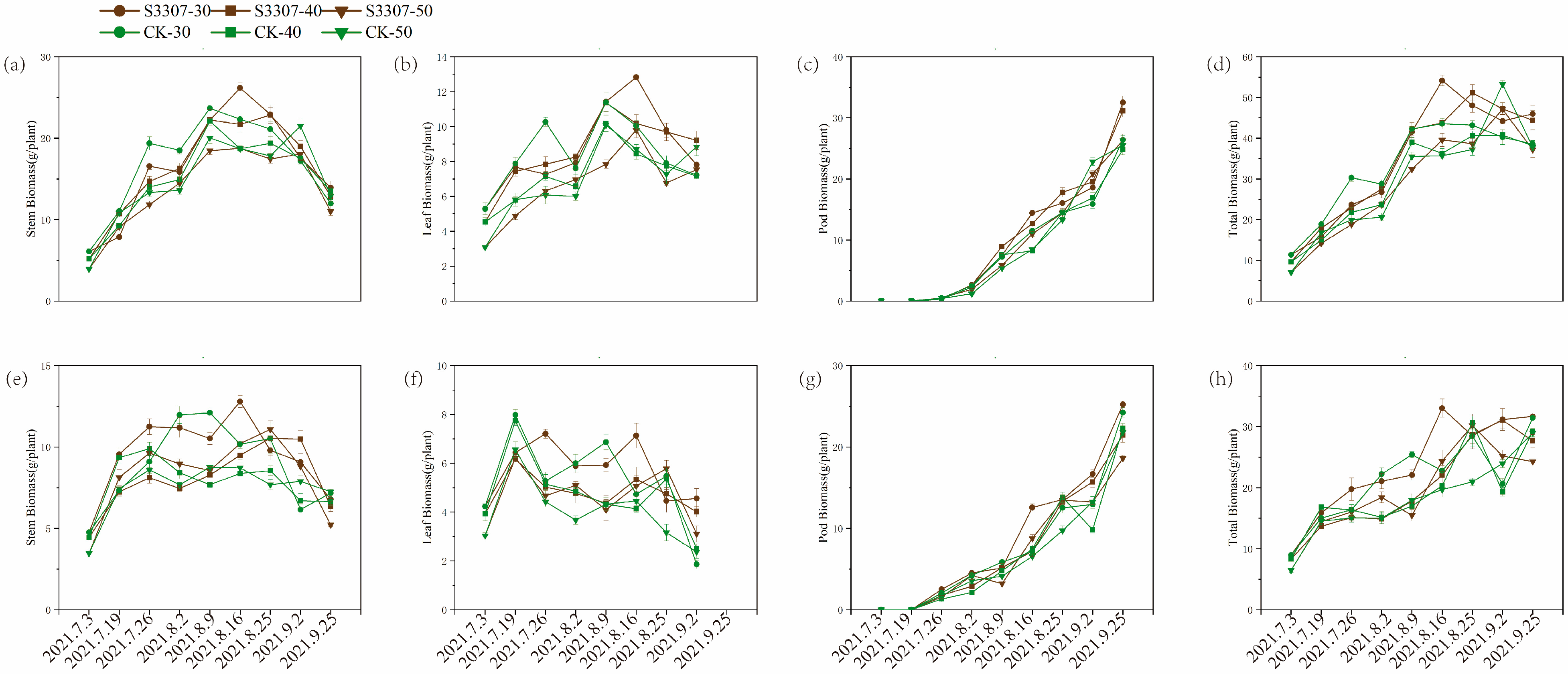
3.2. Plant Dry Weight
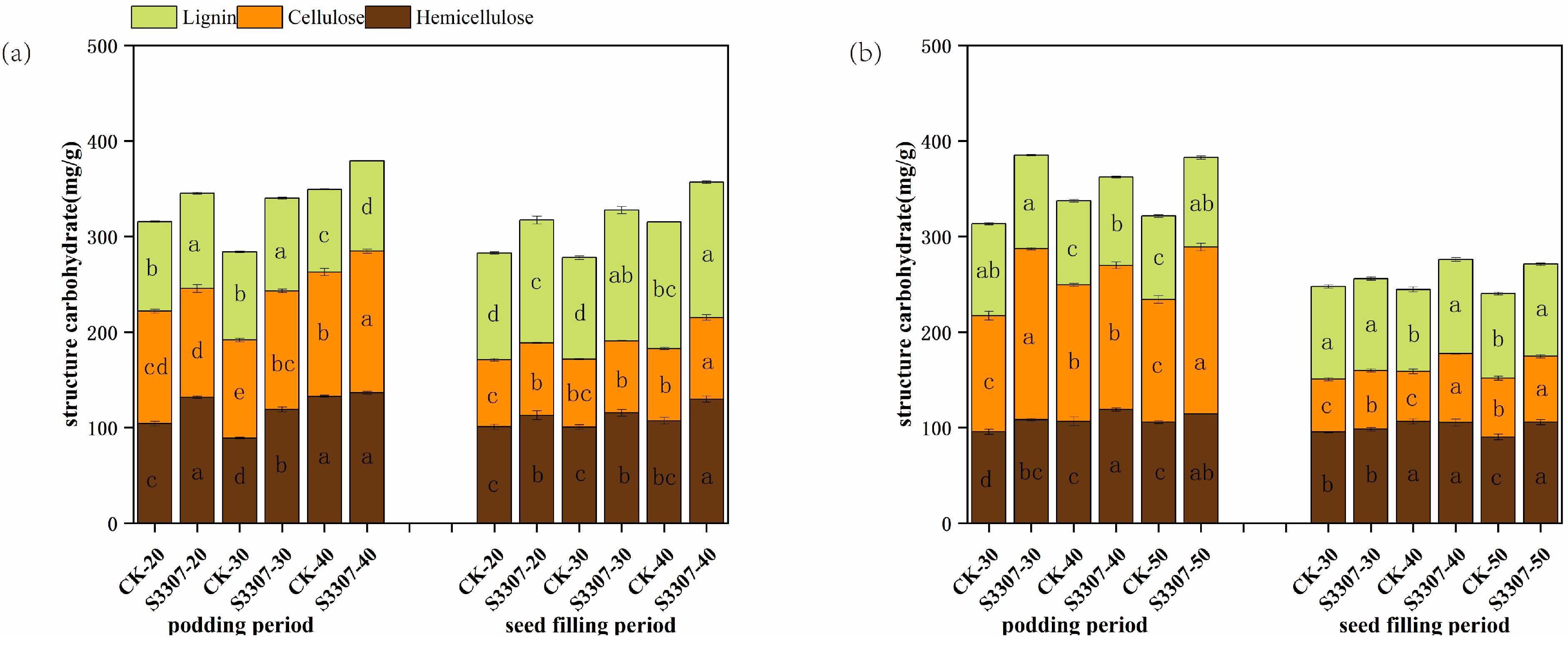
3.3. Carbohydrates Contents
3.4. Breaking Force and Lodging Resistance Coefficient
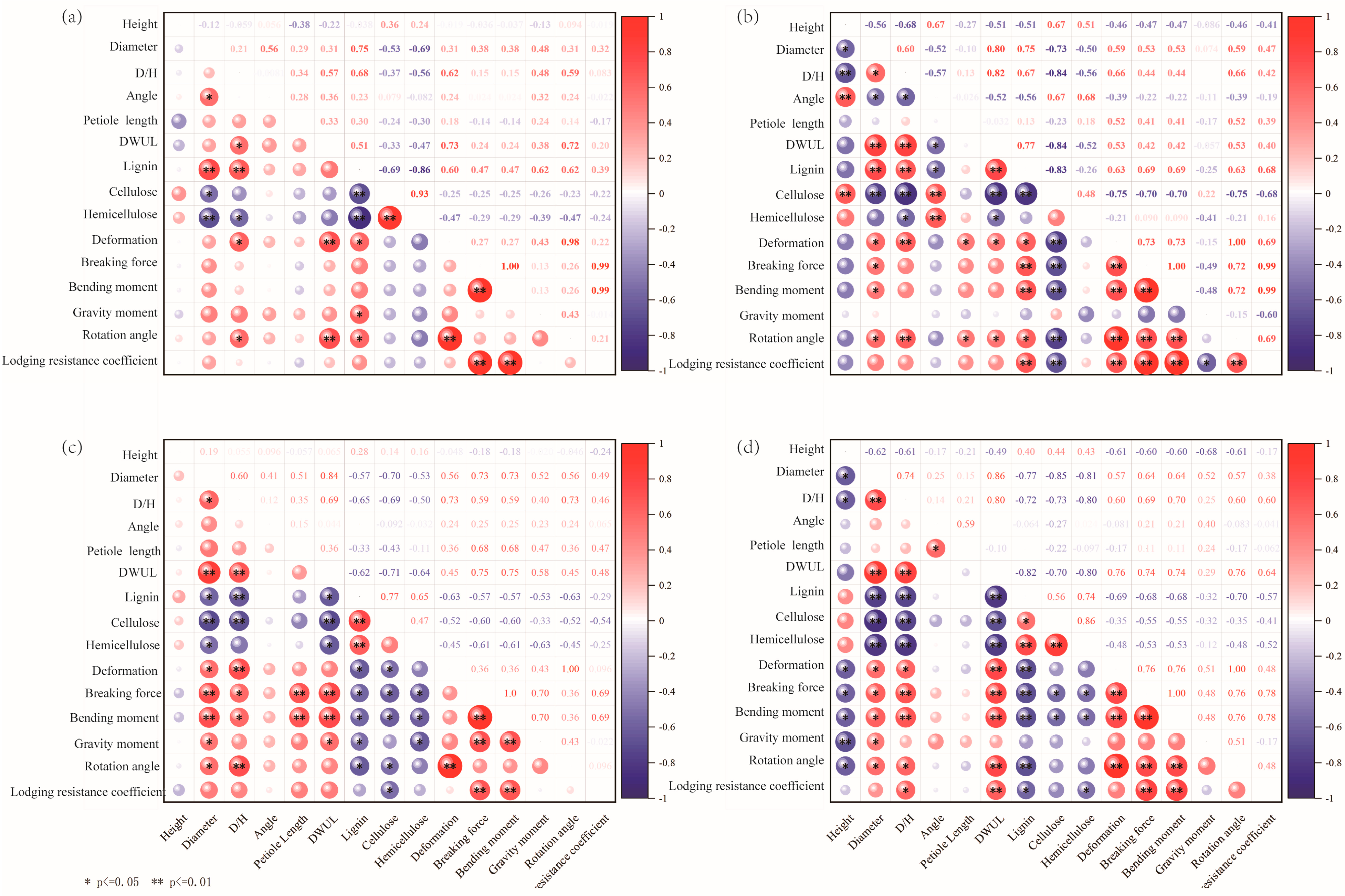
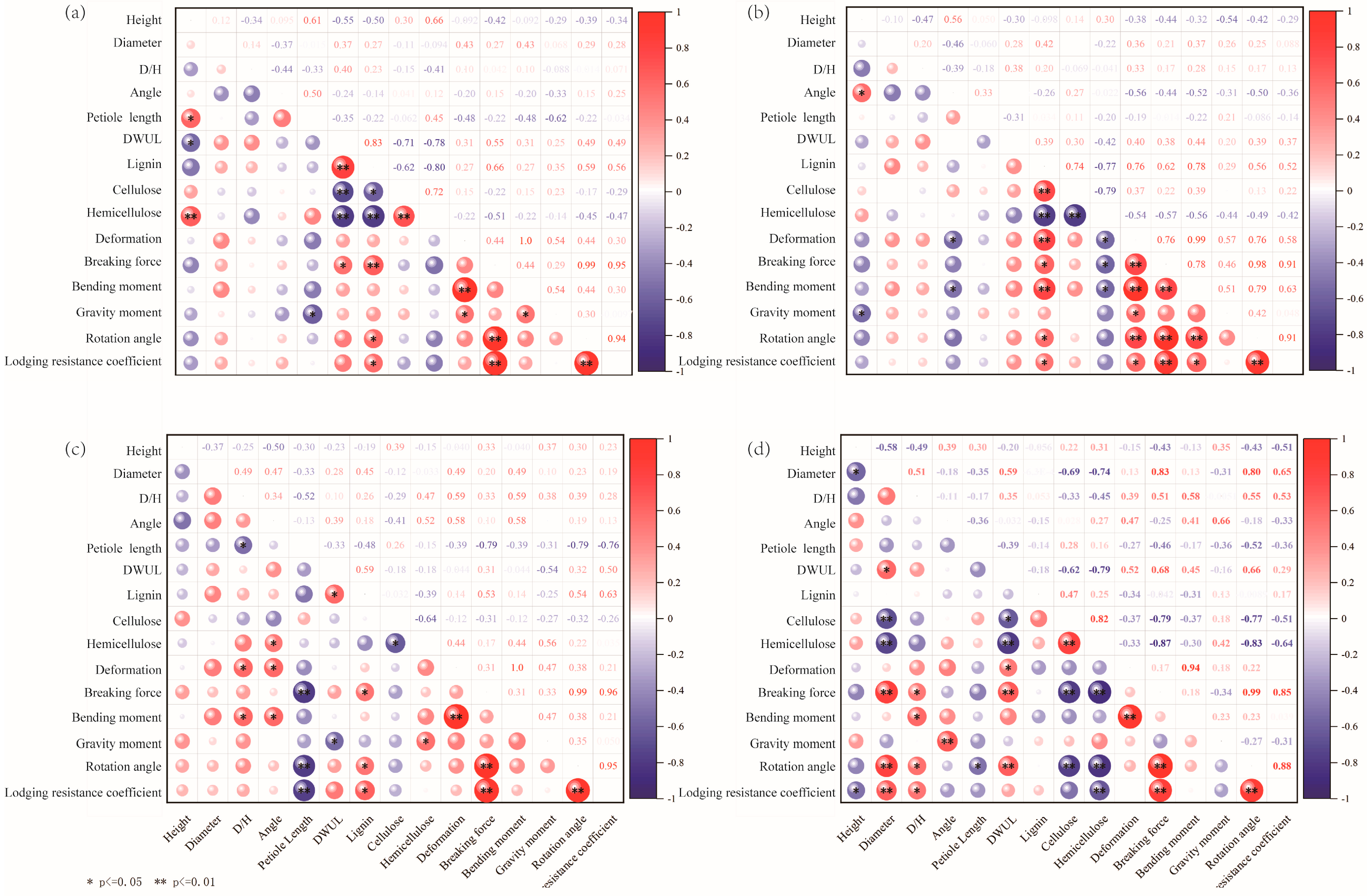
3.5. Correlation Analysis on the Mechanical Properties of Soybean Plants
3.6. Grain Yields and Yield Components
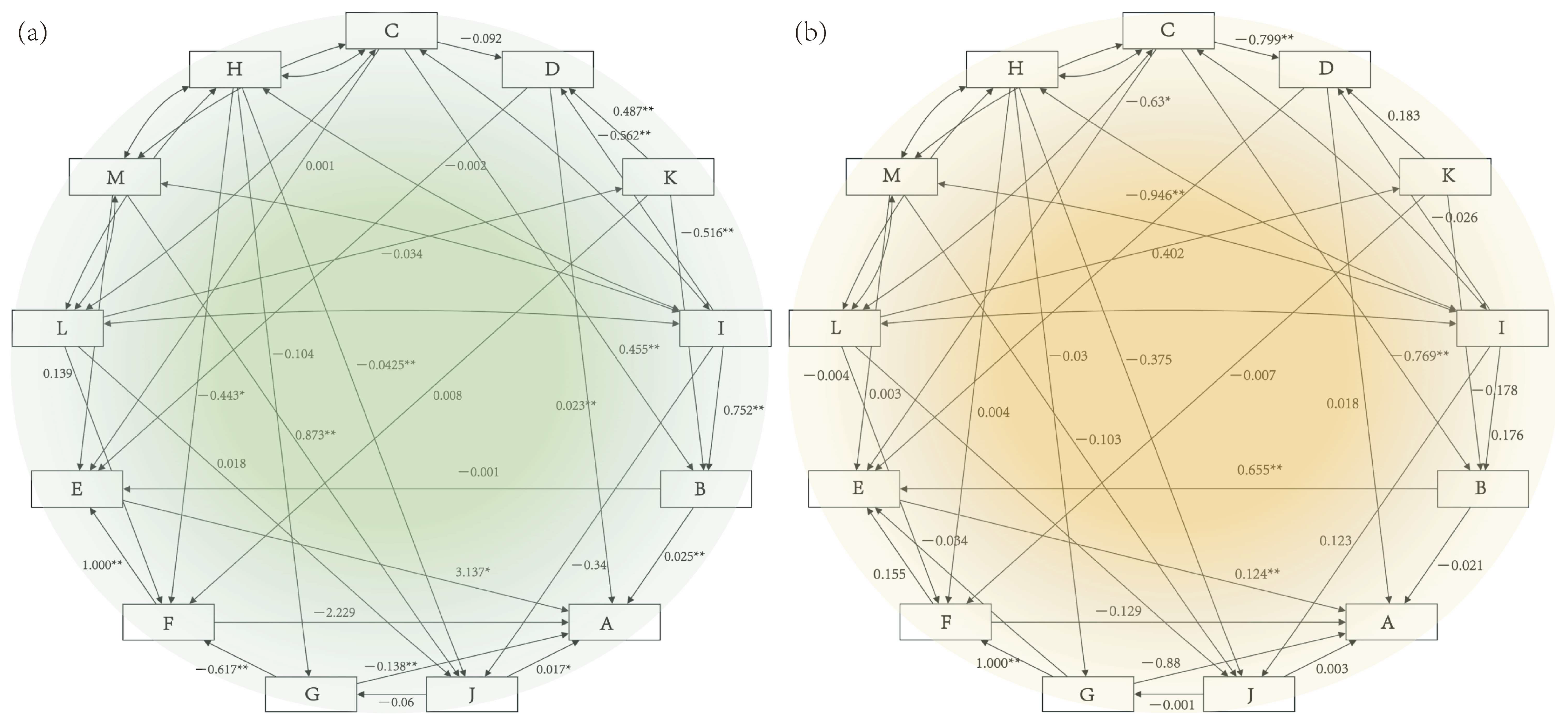
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Le, D.T.; Nishiyama, R.; Watanabe, Y.; Tanaka, M.; Seki, M.; Ham, L.H.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.P.; Shiu, S. Differential Gene Expression in Soybean Leaf Tissues at Late Developmental Stages under Drought Stress Revealed by Genome-Wide Transcriptome Analysis. PLoS ONE 2012, 7, e49522. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Shui, Z.W.; Xu, L.; Yang, Y.H.; Li, Y.; Yuan, X.Q.; Shang, J.; Muhammad, A.A.; Wu, X.L.; Yu, L.; et al. Gibberellins modulate shade-induced soybean hypocotyl elongation downstream of the mutual promotion of auxin and brassinosteroids. Plant Physiol. Biochem. 2020, 150, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Le, D.T.; Nishiyama, R.; Watanabe, Y.; Mochida, K.; YamaguchiShinozaki, K.; Shinozaki, K.; Tran, L.S. Genome-Wide Survey and Expression Analysis of the Plant-Specific NAC Transcription Factor Family in Soybean during Development and Dehydration Stress. DNA Res. 2011, 18, 263–276. [Google Scholar] [CrossRef]
- Xie, Y.R.; Liu, Y.; Wang, H.; Ma, X.J.; Wang, B.B.; Wu, G.X.; Wang, H.Y. Phytochrome-interacting factors directly suppress MIR156 expressionto enhance shade-avoidance syndrome in Arabidopsis. Nat. Commun. 2017, 8, 348. [Google Scholar] [CrossRef]
- Yang, F.; Wang, X.C.; Liao, D.P.; Lu, F.Z.; Gao, R.C.; Liu, W.G.; Yong, T.W.; Wu, X.L.; Du, J.B.; Liu, J.; et al. Yield response to different planting geometries in maize-soybean relay strip intercropping systems. Agron. J. 2015, 107, 296–304. [Google Scholar] [CrossRef]
- Purcell, L.C.; Ball, R.A.; Reaper, J.D.; Vories, E.D. Radiation use efficiency and biomass production in soybean at different plant population densities. Crop. Sci. 2002, 42, 172–177. [Google Scholar] [CrossRef]
- Echezona, B.C. Corn-stalk lodging and borer damage as influenced by varying corn densities and planting geometry with soybean (Glycine max. L. Merrill). Int. Agrophys. 2007, 21, 133–143. [Google Scholar] [CrossRef]
- Khan, S.; Anwar, S.; Kuai, J.; Ullah, S.; Fahad, S.; Zhou, G.S. Optimization of Nitrogen Rate and Planting Density for Improving Yield, Nitrogen Use Efficiency, and Lodging Resistance in Oilseed Rape. Front. Plant Sci. 2017, 8, 532. [Google Scholar] [CrossRef]
- Reza, Y.; Morteza, S.; Hamidreza, M.; Dastan, S.; Alireza, N. Effect of Plant Density on Morphologic Characteristics Related to Lodging and Yield Components in Different Rice Varieties (Oriza sativa L.). J. Agric. Sci. 2011, 4, 31–38. [Google Scholar] [CrossRef][Green Version]
- Zheng, M.J.; Chen, J.; Shi, Y.H.; Li, Y.X.; Yin, Y.P.; Yang, D.Q.; Luo, Y.L.; Pang, D.W.; Xu, X.; Li, W.Q.; et al. Manipulation of lignin metabolism by plant densities and its relationship with lodging resistance in wheat. Sci. Rep. 2016, 7, 19–26. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, J.; Lin, Q.Q.; Li, X.J.; Teng, N.J.; Li, Z.S.; Li, B.; Zhang, A.; Lin, J.X. Effects of stem structure and cell wall components on bending strength in wheat. Chin. Sci. Bull. 2006, 51, 815–823. [Google Scholar] [CrossRef]
- Berg, E.V.D.; Labuschagne, T. The interaction of stem strength with plant density and nitrogen application in wheat progeny from parents with varying stem strength. Acta Agric. Scand. Sect. B Soil. Plant Sci. 2012, 62, 251. [Google Scholar] [CrossRef]
- Liang, S.J.; Li, Z.Q.; Li, X.J.; Xie, H.G.; Zhu, R.S.; Lin, J.X.; Xie, H.A.; Wu, H. Effects of stem structural characters and silicon content on lodging resistance in rice (Oryza satva L.). Res. Crops 2013, 14, 621–636. [Google Scholar]
- Kong, E.; Liu, D.C.; Guo, X.L.; Yang, W.L.; Sun, J.Z.; Li, X.; Zhan, K.H.; Cui, D.Q.; Lin, J.X.; Zhang, A.M. Anatomical and chemical characteristics associated with lodging resistance in wheat. Crop. J. 2013, 1, 43–49. [Google Scholar] [CrossRef]
- Zuber, U.; Winzeler, H.; Messmer, M.M.; Keller, M.; Keller, B.; Schmid, J.E.; Stamp, P. Morphological Traits Associated with Lodging Resistance of Spring Wheat (Triticum aestivum L.). J. Agron. Crop. Sci. 2010, 182, 17–24. [Google Scholar] [CrossRef]
- Jing, B.; Shah, F.; Xiao, E.; Coulter, J.A.; Wu, W. Sprinkler irrigation increases grain yield of sunflower without enhancing the risk of root lodging in a dry semi-humid region. Agric. Water Manag. 2020, 239, 106270. [Google Scholar] [CrossRef]
- Li, C.; Li, C.; Ma, B.L.; Wu, W. The role of ridge-furrow with plastic film mulching system on stem lodging resistance of winter wheat in a dry semi-humid region. Agron. J. 2020, 112, 885–898. [Google Scholar] [CrossRef]
- Shah, A.; Tanveer, M.; Rehman, A.; Anjum, A.; Iqbal, J.; Ahmad, R. Lodging stress in cereal-effects and management: An overview. Environ. Sci. Pollut. Res. 2017, 24, 5222–5237. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhang, R.; Hou, Z.F.; Yan, C.; Xia, X.; Ma, C.M.; Dong, S.K.; Gong, Z.P. Mechanical properties of soybean plants under various plant densities. Crop. Pasture Sci. 2020, 7, 249–259. [Google Scholar] [CrossRef]
- Beeck, C.P.; Wroth, J.; Cowling, W.A. Genetic variation in stem strength in field pea (Pisum sativum L.) and its association with compressed stem thickness. Aust. J. Agr. Res. 2006, 57, 193–199. [Google Scholar] [CrossRef]
- Xiang, D.B.; Yu, X.B.; Wan, Y.; Guo, K.; Yang, W.Y.; Gong, W.Z.; Cui, L. Responses of soybean lodging and lodging-related traits to potassium under shading by maize in relay strip intercropping system. Afr. J. Agr. Res. 2013, 8, 6499–6508. [Google Scholar]
- Xia, H.Y.; Wang, Z.G.; Zhao, J.H.; Sun, J.H.; Bao, X.G.; Christie, P.; Zhang, F.S.; Li, L. Contribution of interspecific interactions and phosphorus application to sustainable and productive intercropping systems. Field Crops Res. 2013, 154, 53–64. [Google Scholar] [CrossRef]
- Yang, S.M.; Xie, L.; Zheng, S.L.; Li, J.; Yuan, J.C. Effects of nitrogen rate and transplanting density on physical and chemical characteristis and lodging resistance of culms in hybrid rice. Acta Agron. Sin. 2009, 35, 93–103. [Google Scholar] [CrossRef]
- Wu, Q.; Wang, Z.; Yang, W. Seedling shading affects morphogenesis and substance accumulatiom of stem in soybean. Soybean Sci. 2007, 26, 868–872. [Google Scholar]
- Jun, X.; Zhao, Y.S.; Gou, L.; Shi, Z.G.; Yao, M.N.; Zhang, W.F. How High Plant Density of Maize Affects Basal Internode Development and Strength Formation. Crop. Sci. 2016, 56, 3295. [Google Scholar] [CrossRef]
- Zhou, W.J.; Leul, M. Uniconazole-induced alleviation of freezing injury in relation to changes in hormonal balance, enzyme activities and lipid peroxidation in winter rape. Plant Growth Regul. 1998, 26, 41–47. [Google Scholar] [CrossRef]
- Han, L.P.; Wang, X.L.; Guo, X.Q.; Rao, M.S.; Steinberger, Y.; Cheng, X.; Xie, G.H. Effects of plant regulators on growth, yield and lodging of sweet sorghum. Res. Crop. 2011, 12, 372–382. [Google Scholar] [CrossRef]
- Yan, Y.H.; Gong, W.Z.; Yang, W.Y.; Wan, Y.; Chen, X.L.; Chen, Z.Q.; Wang, L.Y. Seed treatment with uniconazole powder improves soybean seedling growth under shading by corn in relay strip intercropping system. Plant Prod. Sci. 2010, 13, 367–374. [Google Scholar] [CrossRef]
- Fang, X.; Liu, X.; Zhang, Y.; Huang, K.; Zhang, Y.; Li, Y.; Nie, J.; She, H.; Ruan, R.; Yuan, X.; et al. Effects of uniconazole or gibberellic acid application on the lignin metabolism in relation to lodging resistance of culm in common buckwheat (Fagopyrum esculentum M.). J. Agron. Crop. Sci. 2018, 204, 4. [Google Scholar] [CrossRef]
- Ahmad, I.; Kamran, M.; Guo, Z.Y.; Meng, X.P.; Ali, S.; Zhang, P.; Liu, T.N.; Cai, T.; Han, Q.F. Effects of uniconazole or ethephon foliar application on culm mechanical strength and lignin metabolism, and their relationship with lodging resistance in winter wheat. Crop. Pasture Sci. 2020, 71, 12–22. [Google Scholar] [CrossRef]
- Han, Y.Q.; Shi, Y.; Gao, Y.M.; Zheng, D.F.; Du, J.D.; Zhang, Y.X.; Feng, N.J. Effects of gibberellins and uniconazole on morphology, photosynthetic physiology and yield of soybean. Chin. J. Oil Crop. Sci. 2018, 40, 820–882. [Google Scholar]
- Wan, Y.; Luo, Q.M.; Yan, Y.H.; Yang, W.Y.; Cao, X.N. Response of morphological characters of soybean to application of growth retardant (uniconazole) at third trifoliate stage. Res. Crop. 2013, 14, 792–797. [Google Scholar]
- Zhao, J.J.; Feng, N.F.; Wang, X.X.; Cai, G.R.; Cao, M.Y.; Zheng, D.F.; Zhu, H.D. Uniconazole confers chilling stress tolerance in soybean (Glycine max L.) by modulating photosynthesis, photoinhibition, and activating oxygen metabolism system. Photosynthetica 2019, 57, 446–457. [Google Scholar] [CrossRef]
- Zhang, M.C.; Duan, L.S.; Tian, X.L.; He, Z.P.; Li, J.M.; Wang, B.M.; Li, Z.H. Uniconazole-induced tolerance of soybean to water deficit stress in relation to changes in photosynthesis, hormones and antioxidant system. J. Plant Physiol. 2007, 164, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Feng, N.J.; Liu, C.J.; Zheng, D.F.; Gong, X.W. Effect of uniconazole treatment on the drought tolerance of soybean seedlings. Pak. J. Bot. 2020, 52, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Shan, F.X.; Sun, K.X.; Gong, S.D.; Wang, C.; Ma, C.M.; Zhang, R.; Chao, Y. Effects of Shading on the Internode Critical for Soybean (Glycine max) Lodging. Agronomy 2022, 12, 492. [Google Scholar] [CrossRef]
- Wang, C.Y.; Dai, X.L.; Shi, Y.H.; Wang, Z.L.; Chen, X.G.; He, M.R. Effects of nitrogen application rate and plant density on lodging resistance in winter wheat. Acta Agron. Sin. 2012, 38, 121–128. [Google Scholar] [CrossRef]
- Berry, P.M.; Griffin, J.M.; Sylvester-Bradley, R.; Scott, R.K.; Spink, J.H.; Baker, C.J.; Clare, R.W. Controlling plant form through husbandry to minimize lodging in wheat. Field Crop. Res. 2000, 67, 59–81. [Google Scholar] [CrossRef]
- Hall, A.J.; Sposaro, M.M.; Chimenti, C.A. Stem lodging in sunflower: Variations in stem failure moment of force and structure across Crop. population densities and post–anthesis developmental stages in two genotypes of contrasting susceptibility to lodging. Field Crops Res. 2010, 116, 46–51. [Google Scholar] [CrossRef]
- Iqbal, M.; Ashraf, M. Gibberellic acid mediated induction of salt tolerance in wheat plants: Growth, ionic partitioning, photosynthesis, yield and hormonal homeostasis. Environ. Exp. Bot. 2013, 86, 76–85. [Google Scholar] [CrossRef]
- Wang, C.H.; Liu, D.; Xu, R.N.; Yang, Y.Q.; Liao, H. Mapping of QTLs for leafstalk angle in soybean. Acta Agron. Sin. 2020, 46, 9–19. [Google Scholar]
- Liang, L.; Guo, Y. Correlation study of biomechanical properties and morphological characteristics of Crop. stalks. Trans. Chin. Soc. Agric. Eng. 2008, 24, 1–6. [Google Scholar]
- Luo, M.; Tian, C.; Li, X.; Ling, J. Relationship between morpho-anatomical traits together with chemical components and lodging resistance of stem in rice. Acta Bot. Boreali Occid. Sin. 2007, 27, 2346–2353. [Google Scholar] [CrossRef]
- Wang, Z.K.; Wang, B.; Kuai, J.; Li, Z.; Bai, R.; Zhou, G.S. Planting density and variety intercropping improve organ biomass distribution of rapeseed to alleviate the trade–off between yield and lodging resistance. Crop. Sci. 2021, 61, 4. [Google Scholar] [CrossRef]
- Hussain, S.; Iqbal, N.; Rahman, T.; Liu, T.; Brestic, M.; Safdar, M.E.; Asghar, M.A.; Farooq, M.U.; Shafiq, I.; Ali, A.; et al. Shade effect on carbohydrates dynamics and stem strength of soybean genotypes. Environ. Exp. Bot. 2019, 162, 374–382. [Google Scholar] [CrossRef]
- Kuai, J.; Sun, Y.; Zhou, M.; Zhang, P.; Zuo, Q.; Wu, J.; Zhou, G. The effect of nitrogen application and planting density on the radiation use efficiency and the stem lignin metabolism in rapeseed (Brassica napus L.). Field Crops Res. 2016, 199, 89–98. [Google Scholar] [CrossRef]
- Shao, Y.J.; Shen, Y.S.; He, F.F.; Li, Z.Y. QTL identification for stem fiber, strength and rot resistance in a DH population from an alien introgression of Brassica napus. Plants 2022, 11, 373. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, S.C.; Sayre, K.D. Growth and morphology of spring wheat (Triticum aestivum L.) culms and their association with lodging: Effects of genotypes, N levels and ethephon. Field Crops Res. 2003, 84, 271–290. [Google Scholar] [CrossRef]
- Hu, D.; Liu, X.B.; Wang, C.; Yang, H.; Li, H.X.; Ruan, R.W.; Yuan, X.H.; Yi, Z.L. Expression Analysis of Key Enzyme Genes in Lignin Synthesis of Culm Among Different Lodging Resistances of Common Buckwheat (Fagopyrum esculentum Moench). Sci. Agric. Sin. 2015, 48, 1864–1872. [Google Scholar]
- Chai, L.; Li, H.J.; Zhang, J.F.; Wu, L.T.; Zheng, B.C.; Cui, C.; Jiang, J.; Zuo, S.Q.; Jiang, L.C. Rapid identification of a genomic region conferring dwarfism in rapeseed (Brassica napus L.) YA2016-12. Agronomy 2019, 9, 129. [Google Scholar] [CrossRef]
- Liu, C.; Wang, J.L.; Huang, T.D.; Wang, F.; Yuan, F.; Cheng, X.M.; Zhang, Y.; Shi, S.W.; Wu, J.S.; Liu, K.D. A missense mutation in the VHYNP motif of a DELLA protein causes a semi-dwarf mutant phenotype in Brassica napus. Theor. Appl. Genet. 2010, 121, 249–258. [Google Scholar] [CrossRef]
- Sanchez, P.; Nehlin, L.; Greb, T. From thin to thick: Major transitions during stem development. Trends Plant Sci. 2012, 17, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Pinera–Chavez, F.J.; Berry, P.M.; Foulkes, M.J.; Reynolds, M.P. Avoiding lodging in irrigated spring wheat. II. Genetic variation of stem and root structural properties. Field Crop. Res. 2016, 196, 64–74. [Google Scholar] [CrossRef]
- Wu, W.; Ma, B.L. Erect–leaf posture promotes lodging resistance in oat plants under high plant population. Eur. J. Agron. 2019, 103, 175–187. [Google Scholar] [CrossRef]
- Zhao, B.; Wang, B.; Li, Z.H.; Guo, T.; Zhao, J.W.; Guan, Z.L.; Liu, K.D. Identification and characterization of a new dwarf locus DS-4 encoding an Aux/IAA7 protein in Brassica napus. Theor. Appl. Genet. 2019, 132, 1435–1449. [Google Scholar] [CrossRef]
- Li, S.C.; Sun, Z.H.; Sang, Q.; Qin, C.; Kong, L.P.; Huang, X.; Liu, H.; Su, T.; Li, H.Y.; He, M.L.; et al. Soybean reduced internode 1 determines internode length and improves grain yield at dense planting. Nat. Commun. 2023, 14, 7939. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Period | Treatment | DWUL (g/cm) | Deformation (cm) | Breaking Force (N) | Bending Moment (Nm) | Gravity Moment (Nm) | Rotation Angle (°) | Lodging Resistance Coefficient | |
|---|---|---|---|---|---|---|---|---|---|
| podding period | D20 | CK | 0.20 ± 0.01 ab | 1.54 ± 0.05 ab | 58.41 ± 5.75 b | 4.67 ± 0.46 b | 0.78 ± 0.01 a | 10.9 ± 0.35 ab | 6.02 ± 0.65 b |
| S3307 | 0.22 ± 0 a | 1.68 ± 0.05 a | 70.96 ± 0.65 a | 5.68 ± 0.05 a | 0.74 ± 0 b | 11.86 ± 0.34 a | 7.68 ± 0.05 a | ||
| D30 | CK | 0.20 ± 0.01 ab | 1.42 ± 0.09 bc | 48.33 ± 3.25 bc | 3.87 ± 0.26 bc | 0.76 ± 0.01 ab | 10.06 ± 0.6 ab | 5.09 ± 0.38 bc | |
| S3307 | 0.21 ± 0.01 a | 1.5 ± 0.04 ab | 50.64 ± 4.14 bc | 4.05 ± 0.33 bc | 0.78 ± 0.01 a | 10.62 ± 0.31 bc | 5.15 ± 0.4 bc | ||
| D40 | CK | 0.17 ± 0.01 bc | 1.24 ± 0.07 c | 43 ± 1.97 c | 3.44 ± 0.16 c | 0.74 ± 0.01 b | 8.81 ± 0.52 c | 4.64 ± 0.2 c | |
| S3307 | 0.16 ± 0 c | 1.3 ± 0.07 c | 47.93 ± 2.12 bc | 3.83 ± 0.17 bc | 0.76 ± 0.01 ab | 9.23 ± 0.49 c | 5.05 ± 0.27 bc | ||
| seed filling period | D20 | CK | 0.19 ± 0 b | 1.54 ± 0.02 b | 63.44 ± 2.68 a | 4.44 ± 0.19 a | 1.09 ± 0.05 a | 12.41 ± 0.19 b | 4.09 ± 0.15 ab |
| S3307 | 0.25 ± 0 a | 1.72 ± 0.05 a | 68.2 ± 2.75 a | 4.78 ± 0.19 a | 1.1 ± 0.05 a | 13.8 ± 0.38 a | 4.35 ± 0.19 a | ||
| D30 | CK | 0.17 ± 0.01 c | 1.5 ± 0.04 b | 56.05 ± 1.44 bc | 3.92 ± 0.1 bc | 1.08 ± 0.04 a | 12.09 ± 0.35 b | 3.63 ± 0.07 c | |
| S3307 | 0.20 ± 0.01 b | 1.5 ± 0.03 b | 56.77 ± 1.56 b | 3.97 ± 0.11 b | 1.07 ± 0.03 a | 12.09 ± 0.25 b | 3.71 ± 0.14 bc | ||
| D40 | CK | 0.15 ± 0 d | 1.36 ± 0.05 c | 50.2 ± 1.71 c | 3.51 ± 0.12 bc | 0.99 ± 0.01 a | 10.99 ± 0.4 c | 3.56 ± 0.12 c | |
| S3307 | 0.17 ± 0 c | 1.48 ± 0.04 b | 53.09 ± 1.4 bc | 3.72 ± 0.1 bc | 1.02 ± 0.02 a | 11.94 ± 0.29 b | 3.66 ± 0.15 bc | ||
| Analysis of variance | |||||||||
| Density(D) | 60.883 ** | 25.931 ** | 38.37 ** | 36.724 ** | 4.289 * | 26.438 ** | 25.097 ** | ||
| S3307(S) | 33.879 ** | 9.611 ** | 8.464 ** | 8.422 ** | 0.13 ns | 10.088 ** | 6.846 * | ||
| Period(P) | 0.611 ns | 5.04 * | 8.646 ** | 2.548 ns | 358.404 ** | 77.356 ** | 116.624 ** | ||
| D × S | 6.981 ** | 1.246 ns | 1.689 ns | 1.688 ns | 0.518 ns | 1.349 ns | 2.773 ns | ||
| D × P | 5.864 ** | 1.68 ns | 1.27 ns | 2.156 ns | 2.476 ns | 1.099 ns | 7.019 ** | ||
| S × P | 15.283 ** | 0.011 ns | 1.39 ns | 1.754 ns | 0.13 ns | 0.088 ns | 2.954 ns | ||
| D × S × P | 0.475 ns | 0.491 ns | 0.381 ns | 0.452 ns | 0.623 ns | 0.599 ns | 1.692 ns | ||
| Period | Treatment | DWUL (g/cm) | Deformation (cm) | Breaking Force (N) | Bending Moment (Nm) | Gravity Moment (Nm) | Rotation Angle (°) | Lodging Resistance Coefficient | |
|---|---|---|---|---|---|---|---|---|---|
| podding period | D30 | CK | 0.22 ± 0.01 a | 1.72 ± 0.07 b | 29.07 ± 0.56 ab | 0.12 ± 0 ab | 0.21 ± 0 ab | 76.44 ± 0.25 ab | 9.68 ± 0.31 a |
| S3307 | 0.18 ± 0.01 b | 2.08 ± 0.05 a | 31.78 ± 1.46 a | 0.14 ± 0 a | 0.22 ± 0 a | 77.48 ± 0.57 a | 10.21 ± 0.49 a | ||
| D40 | CK | 0.14 ± 0.01 c | 1.62 ± 0.13 b | 25.82 ± 1.61 bc | 0.12 ± 0.01 b | 0.21 ± 0 ab | 74.6 ± 1 abc | 8.64 ± 0.58 a | |
| S3307 | 0.15 ± 0.02 bc | 1.7 ± 0.09 b | 26.55 ± 0.59 bc | 0.12 ± 0.01 b | 0.21 ± 0.01 ab | 75.2 ± 0.31 abc | 8.93 ± 0.23 a | ||
| D50 | CK | 0.14 ± 0.01 c | 1.24 ± 0.2 c | 23.32 ± 2.12 c | 0.09 ± 0.01 c | 0.19 ± 0 c | 72.72 ± 1.74 c | 8.4 ± 0.78 a | |
| S3307 | 0.14 ± 0.01 c | 1.56 ± 0.1 bc | 24.26 ± 1.66 c | 0.11 ± 0.01 bc | 0.2 ± 0.01 bc | 73.64 ± 1.07 bc | 8.6 ± 0.73 a | ||
| seed filling period | D30 | CK | 0.17 ± 0.02 b | 1.76 ± 0.02 a | 37.76 ± 2.83 b | 0.14 ± 0 a | 0.32 ± 0.01 a | 77.78 ± 0.91 b | 9.45 ± 0.72 a |
| S3307 | 0.25 ± 0.02 a | 1.8 ± 0.07 a | 51.65 ± 1.65 a | 0.14 ± 0.01 a | 0.31 ± 0 a | 81.16 ± 0.28 a | 9.7 ± 0.32 a | ||
| D40 | CK | 0.14 ± 0.01 b | 1.72 ± 0.1 a | 32.07 ± 2.61 bcd | 0.14 ± 0.01 a | 0.33 ± 0.01 a | 75.7 ± 0.96 bcd | 7.72 ± 0.52 bc | |
| S3307 | 0.16 ± 0.02 b | 1.74 ± 0.05 a | 35.15 ± 1.54 bc | 0.14 ± 0 a | 0.32 ± 0.01 a | 77.09 ± 0.51 bc | 8.74 ± 0.33 ab | ||
| D50 | CK | 0.14 ± 0.01 b | 1.62 ± 0.08 a | 28.23 ± 1.37 d | 0.13 ± 0.01 a | 0.31 ± 0 a | 74.05 ± 0.7 d | 7.19 ± 0.33 c | |
| S3307 | 0.15 ± 0.01 b | 1.7 ± 0.03 a | 30.82 ± 1.06 cd | 0.14 ± 0 a | 0.32 ± 0.01 a | 75.39 ± 0.45 cd | 7.55 ± 0.14 bc | ||
| Analysis of variance | |||||||||
| Density(D) | 22.166 ** | 10.853 ** | 42.366 ** | 9.629 ** | 2.682 ns | 26.256 ** | 13.896 ** | ||
| S3307(S) | 3.66 ns | 7.613 ** | 16.127 ** | 6.989 * | 0.038 ns | 8.91 ** | 2.368 ns | ||
| Period(P) | 0.479 ns | 1.658 ns | 84.789 ** | 30.727 ** | 1070.104 ** | 14.614 ** | 5.625 * | ||
| D×S | 0.171 ns | 0.846 ns | 4.705 * | 0.902 ns | 1.261 ns | 0.631 ns | 0.147 ns | ||
| D × P | 0.022 ns | 4.071 * | 6.926 ** | 3.695 * | 3.953 * | 0.468 ns | 0.626 ns | ||
| S × P | 6.577 * | 3.613 ns | 6.486 * | 1.636 ns | 1.858 ns | 1.504 ns | 0.131 ns | ||
| D × S × P | 4.598 * | 0.5 ns | 2.376 ns | 0.611 ns | 1.204 ns | 0.371 ns | 0.255 ns | ||
| Cultivar | Treatment | Pod Number per Plant | The Internode of the First Pod | Number of Internodes | Grain Number per Plant | 100-Grain Weight (g) | Grain Yield (kg/hm2) | |
|---|---|---|---|---|---|---|---|---|
| HN84 | D20 | CK | 38.71 ± 0.71 bc | 4.14 ± 0.26 ab | 16.86 ± 0.26 a | 94.29 ± 4.07 b | 19.65 ± 0.5 ab | 2906.00 ± 65.65 cd |
| S3307 | 49.57 ± 1.65 a | 3.71 ± 0.18 b | 16.43 ± 0.37 a | 117.14 ± 4.83 a | 19.95 ± 0.27 ab | 2981.00 ± 60.20 c | ||
| D30 | CK | 36.00 ± 1.05 cd | 4.14 ± 0.26 ab | 17.00 ± 0.31 a | 88.57 ± 2.20 b | 20.42 ± 0.16 a | 3076.94 ± 63.85 bc | |
| S3307 | 46.29 ± 1.96 a | 4.00 ± 0.22 ab | 16.71 ± 0.36 a | 110.43 ± 4.33 a | 20.15 ± 0.41 ab | 3333.35 ± 110.60 a | ||
| D40 | CK | 33.43 ± 0.84 d | 4.57 ± 0.2 a | 16.71 ± 0.29 a | 86.86 ± 1.24 b | 20.29 ± 0.10 a | 2735.06 ± 60.44 d | |
| S3307 | 40.14 ± 0.91 b | 4.00 ± 0.22 ab | 17.14 ± 0.26 a | 95.43 ± 4.47 b | 19.21 ± 0.35 b | 3247.88 ± 45.90 ab | ||
| Analysis of variance | ||||||||
| Density(D) | 16.941 ** | 1.267ns | 0.141 ns | 7.547 ** | 1.621 ns | 12.934 ** | ||
| S3307(S) | 80.053 ** | 4.267 * | 0.459 ns | 33.402 ** | 1.743 ns | 39.593 ** | ||
| D × S | 1.560 ns | 0.467 ns | 1.094 ns | 2.244 ns | 2.201 ns | 8.064 ** | ||
| HN60 | D30 | CK | 36.57 ± 1.69 cd | 1.86 ± 0.14 ab | 12.14 ± 0.26 a | 101.00 ± 0.82 bc | 15.43 ± 0.23 b | 2906.00 ± 55.15 c |
| S3307 | 43.71 ± 1.21 a | 1.57 ± 0.20 b | 12.29 ± 0.29 a | 109.29 ± 2.63 a | 16.40 ± 0.15 a | 3333.35 ± 110.60 b | ||
| D40 | CK | 35.43 ± 1.09 cd | 2.29 ± 0.29 ab | 11.86 ± 0.14 a | 95.29 ± 4.33 c | 16.47 ± 0.35 a | 3247.88 ± 65.658 b | |
| S3307 | 41.00 ± 1.27 ab | 2.14 ± 0.26 ab | 11.86 ± 0.14 a | 106.14 ± 1.81 ab | 15.70 ± 0.36 ab | 3615.33 ± 44.20 a | ||
| D50 | CK | 33.86 ± 1.01 d | 2.43 ± 0.30 ab | 11.86 ± 0.34 a | 82.14 ± 2.01 d | 15.87 ± 0.25 ab | 3162.41 ± 64.68 b | |
| S3307 | 38.00 ± 1.45 bc | 2.14 ± 0.34 a | 11.71 ± 0.18 a | 99.00 ± 2.23 bc | 16.26 ± 0.19 ab | 3675.23 ± 50.16 a | ||
| Analysis of variance | ||||||||
| Density(D) | 5.220 ** | 2.803 ns | 1.86 ns | 17.355 ** | 0.231 ns | 21.121 ** | ||
| S3307(S) | 27.774 ** | 1.230 ns | 0.00 ns | 33.588 ** | 0.796 ns | 96.585 ** | ||
| D × S | 0.660 ns | 0.49 ns | 0.18 ns | 1.504 ns | 5.551 * | 0.905 ns | ||
| Cultivar | Treatment | Number of Internodes | Grain Number per Plant | 100-Grain Weight (g) | Grain Yield (kg/hm2) | |
|---|---|---|---|---|---|---|
| HN84 | D20 | CK | 19.25 ± 0.16 a | 116.94 ± 1.33 bc | 18.94 ± 0.30 a | 2539.00 ± 77.96 c |
| S3307 | 19.38 ± 0.26 a | 128.13 ± 3.35 a | 19.03 ± 0.26 a | 2563.11 ± 38.56 c | ||
| D30 | CK | 19.13 ± 0.30 a | 113.94 ± 4.69 bc | 19.10 ± 0.27 a | 2805.26 ± 36.26 b | |
| S3307 | 19.25 ± 0.31 a | 122.81 ± 4.86 ab | 19.38 ± 0.25 a | 2973.24 ± 69.55 a | ||
| D40 | CK | 19.13 ± 0.23 a | 111.00 ± 3.92 c | 18.69 ± 0.32 a | 2475.44 ± 25.04 c | |
| S3307 | 19.50 ± 0.19 a | 114.81 ± 1.36 bc | 19.09 ± 0.27 a | 2576.44 ± 23.81 c | ||
| Analysis of variance | ||||||
| Density(D) | 0.170 ns | 3.689 * | 0.849 ns | 33.128 ** | ||
| S3307(S) | 1.061 ns | 7.519 ** | 1.268 ns | 5.761 * | ||
| D × S | 0.170 ns | 0.563 ns | 0.159 ns | 1.042 ns | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, C.; Shan, F.; Wang, C.; Lyu, X.; Wu, Y.; Yan, S.; Ma, C. Positive Correlation of Lodging Resistance and Soybean Yield under the Influence of Uniconazole. Agronomy 2024, 14, 754. https://doi.org/10.3390/agronomy14040754
Yan C, Shan F, Wang C, Lyu X, Wu Y, Yan S, Ma C. Positive Correlation of Lodging Resistance and Soybean Yield under the Influence of Uniconazole. Agronomy. 2024; 14(4):754. https://doi.org/10.3390/agronomy14040754
Chicago/Turabian StyleYan, Chao, Fuxin Shan, Chang Wang, Xiaochen Lyu, Yuanyi Wu, Shuangshuang Yan, and Chunmei Ma. 2024. "Positive Correlation of Lodging Resistance and Soybean Yield under the Influence of Uniconazole" Agronomy 14, no. 4: 754. https://doi.org/10.3390/agronomy14040754
APA StyleYan, C., Shan, F., Wang, C., Lyu, X., Wu, Y., Yan, S., & Ma, C. (2024). Positive Correlation of Lodging Resistance and Soybean Yield under the Influence of Uniconazole. Agronomy, 14(4), 754. https://doi.org/10.3390/agronomy14040754