
Characterization and Transcriptome Analysis Reveal Exogenous GA3 Inhibited Rosette Branching via Altering Auxin Approach in Flowering Chinese Cabbage

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Exogenous GA3 Spraying Treatment
2.2. RNA Extraction and Transcriptome Sequencing
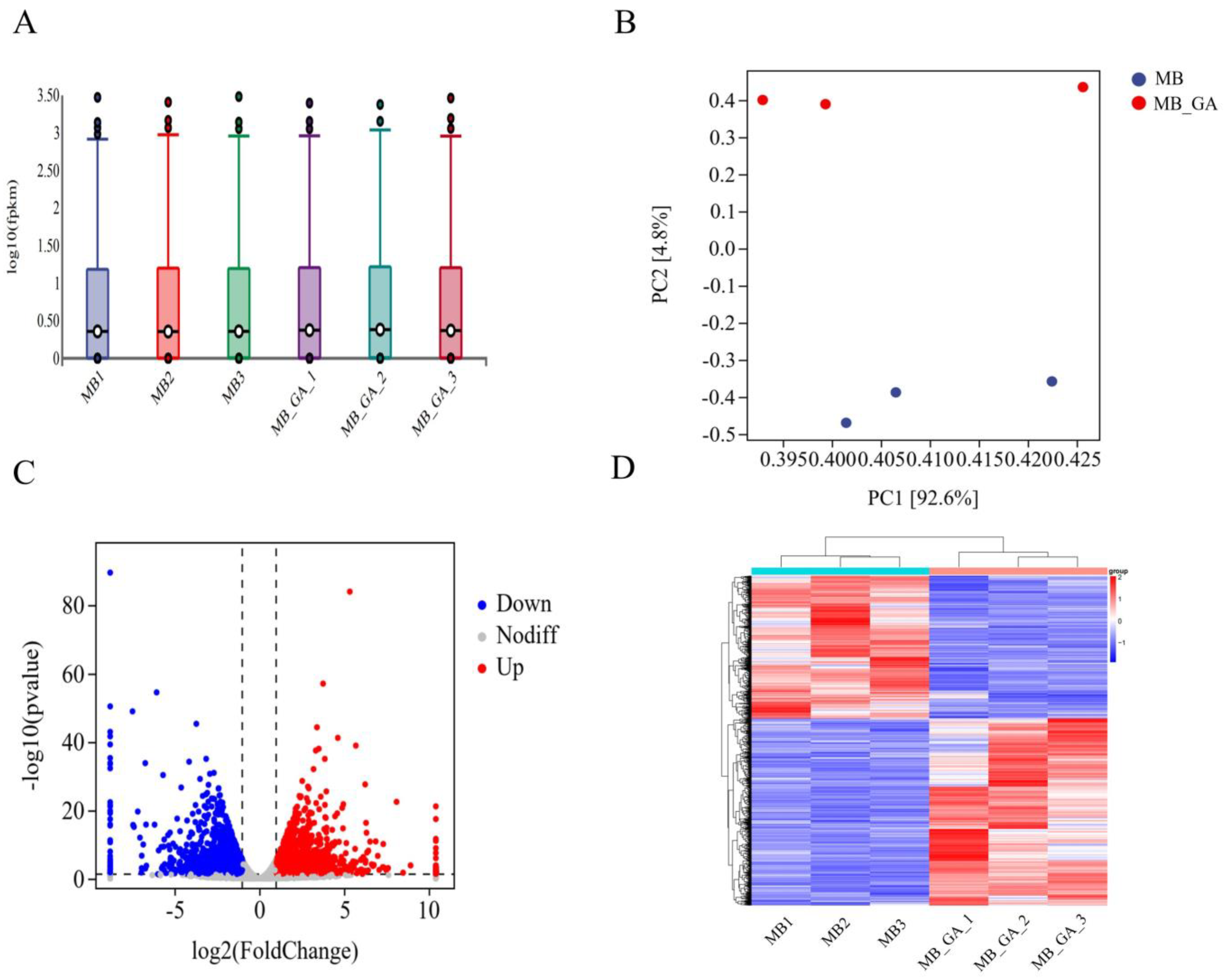
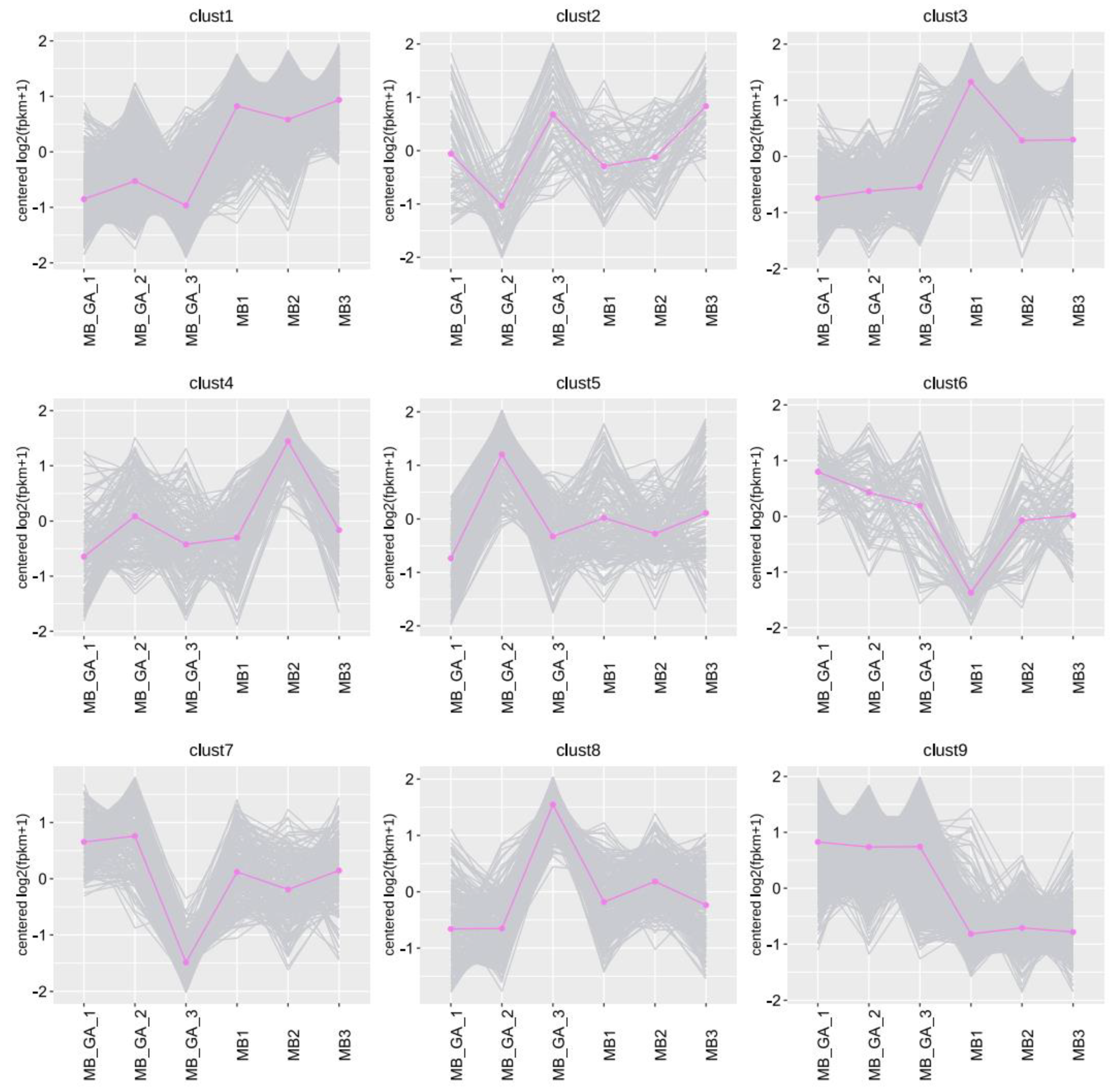
2.3. Transcriptome Analysis
2.4. Metabolite Profiling of Hormone Content Using UPLC-MS/MS
2.5. Real-Time Quantitative PCR Analysis (qRT-PCR)
2.6. Statistical Analysis
3. Results
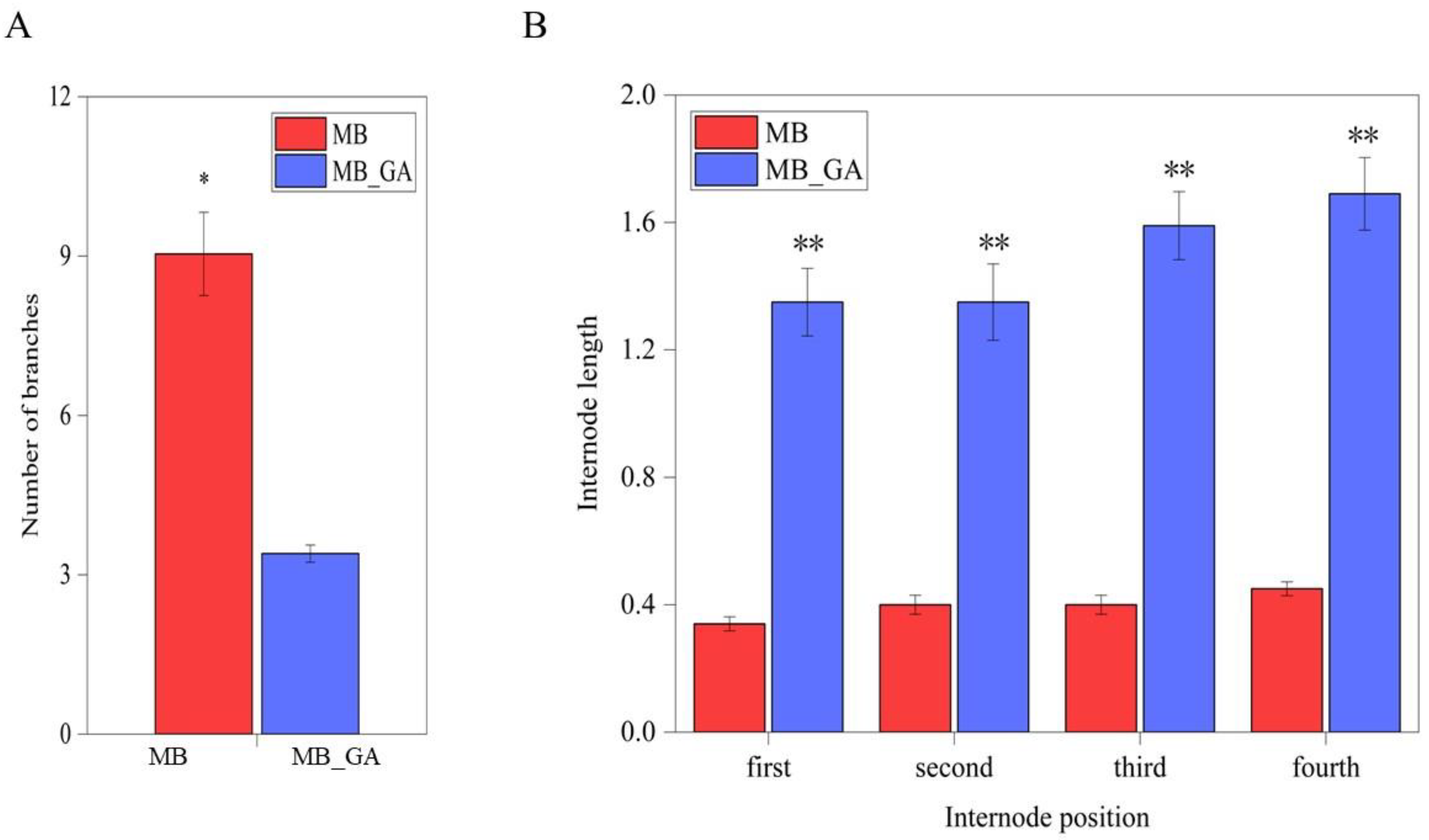
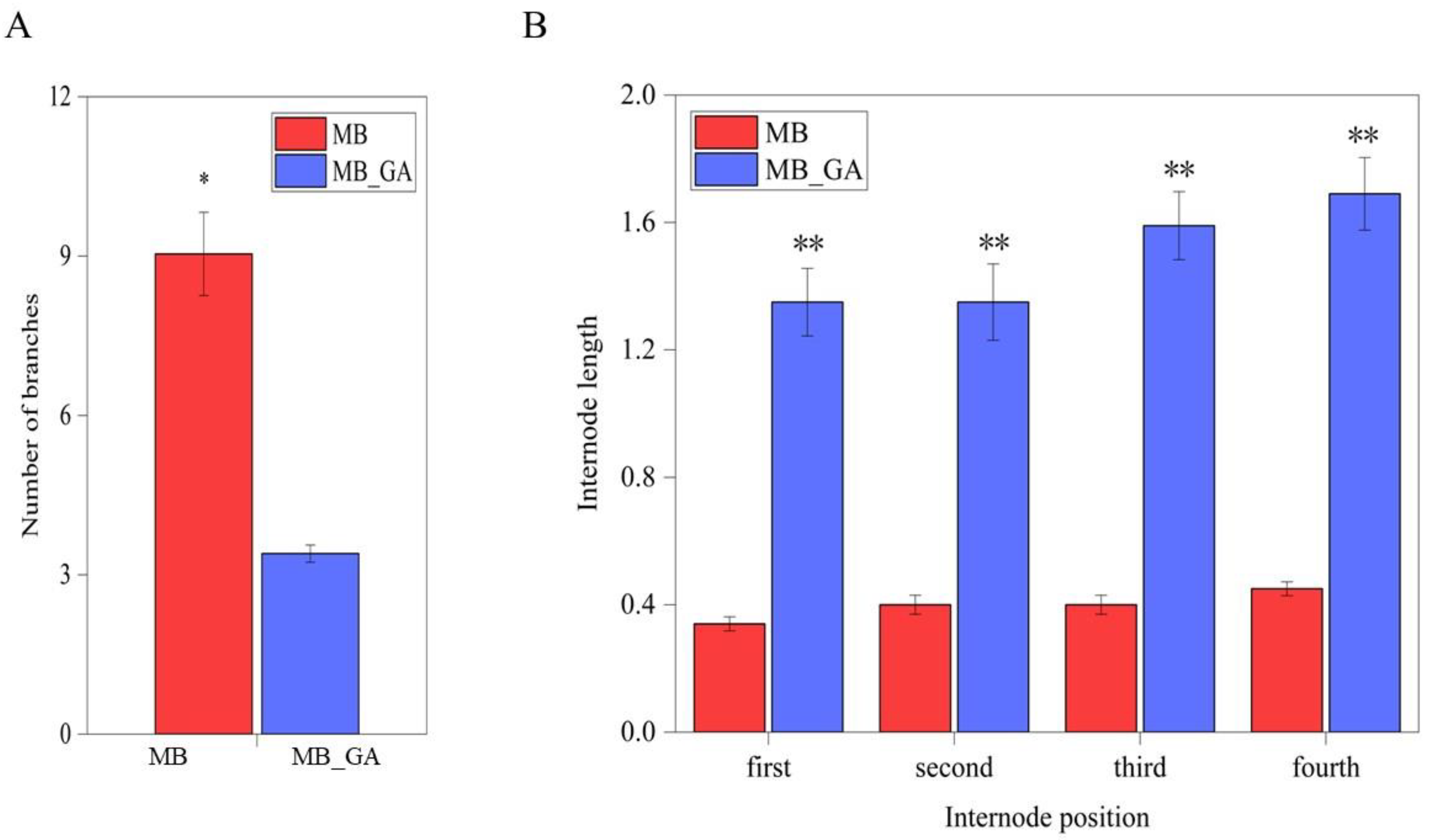
3.1. Exogenous GA3 Obviously Inhibited Rosette Branching
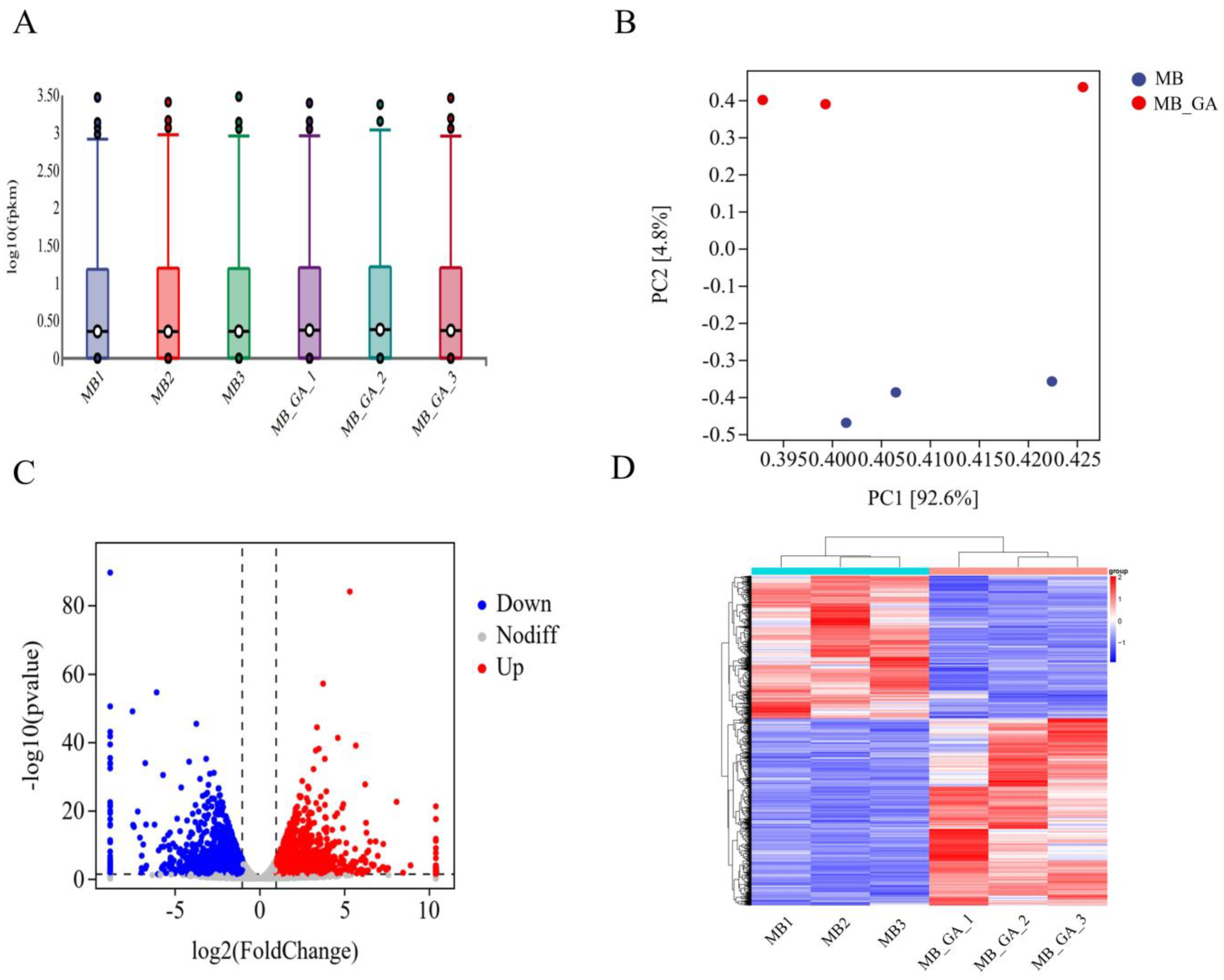
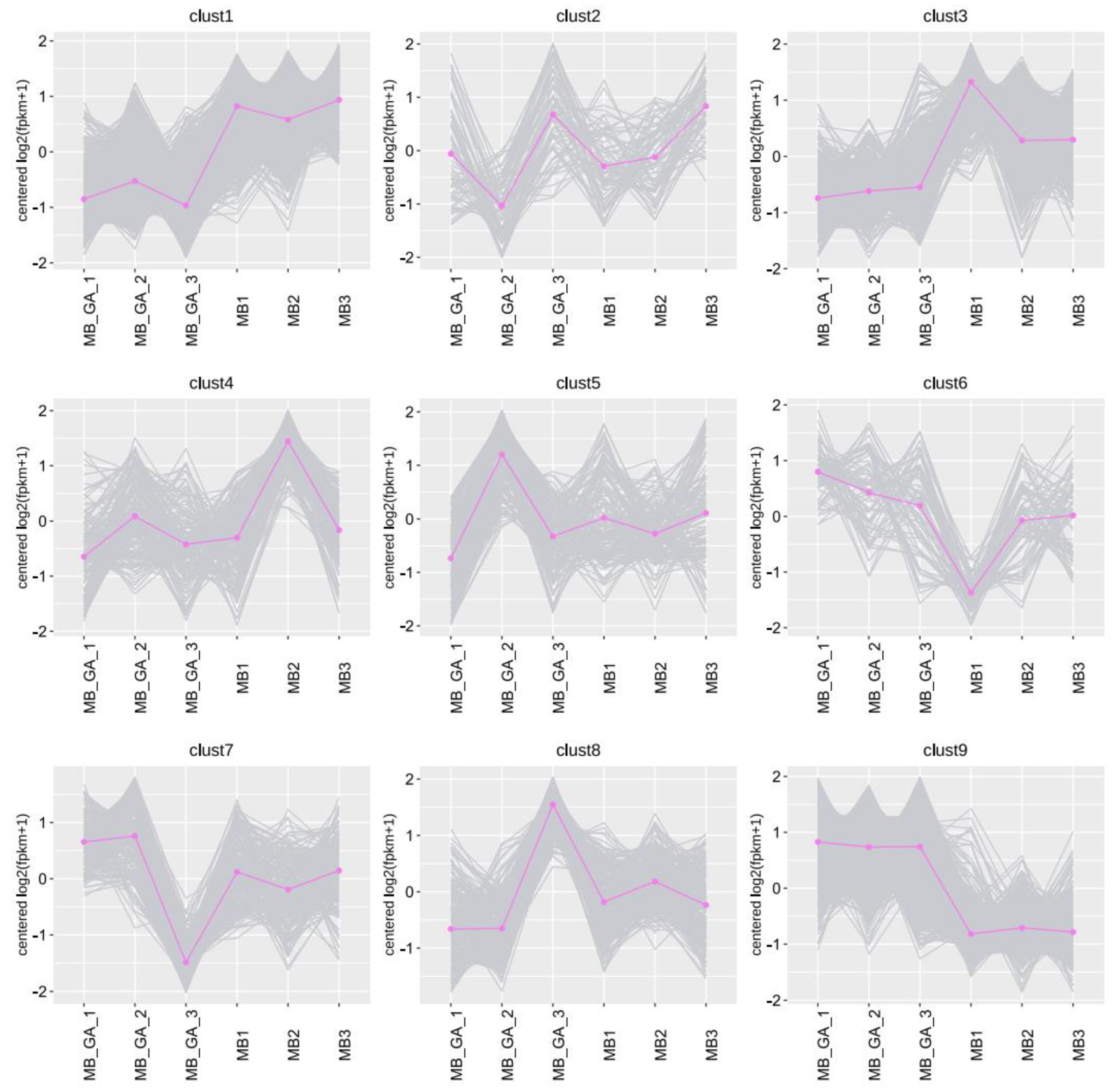
3.2. Influences of Exogenous GA3 on Gene Expressions
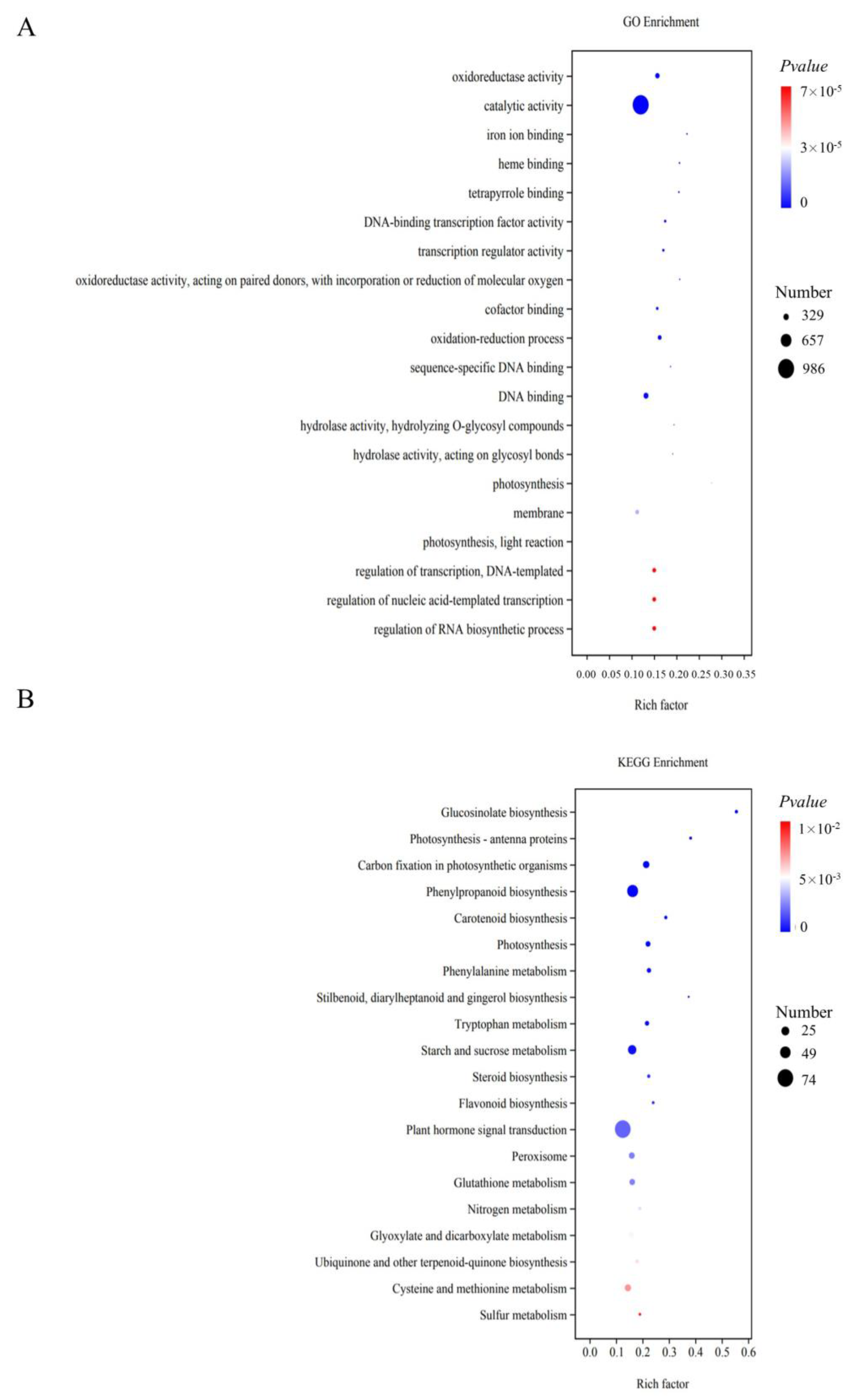
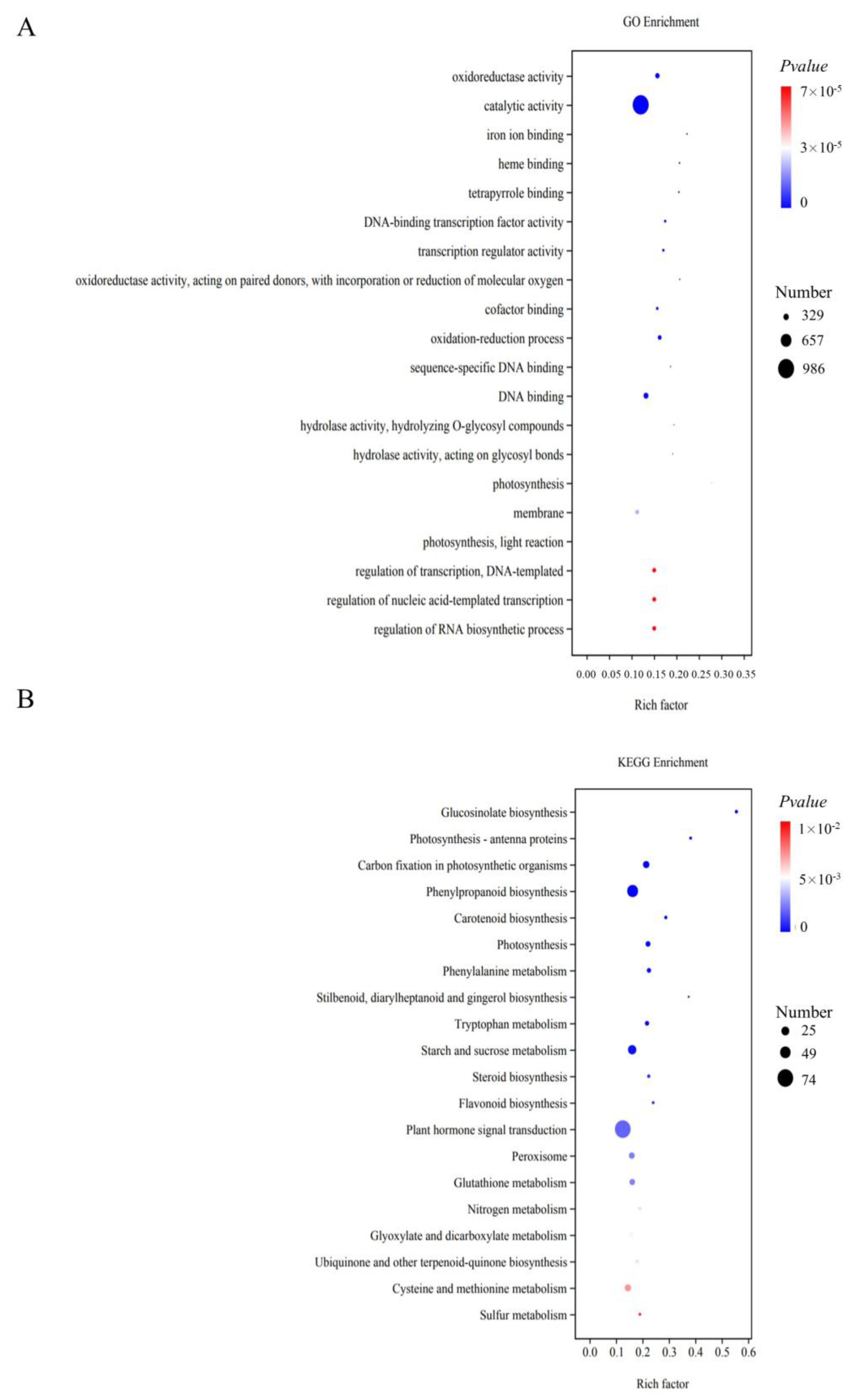
3.3. Enrichment Analysis for the DEGs
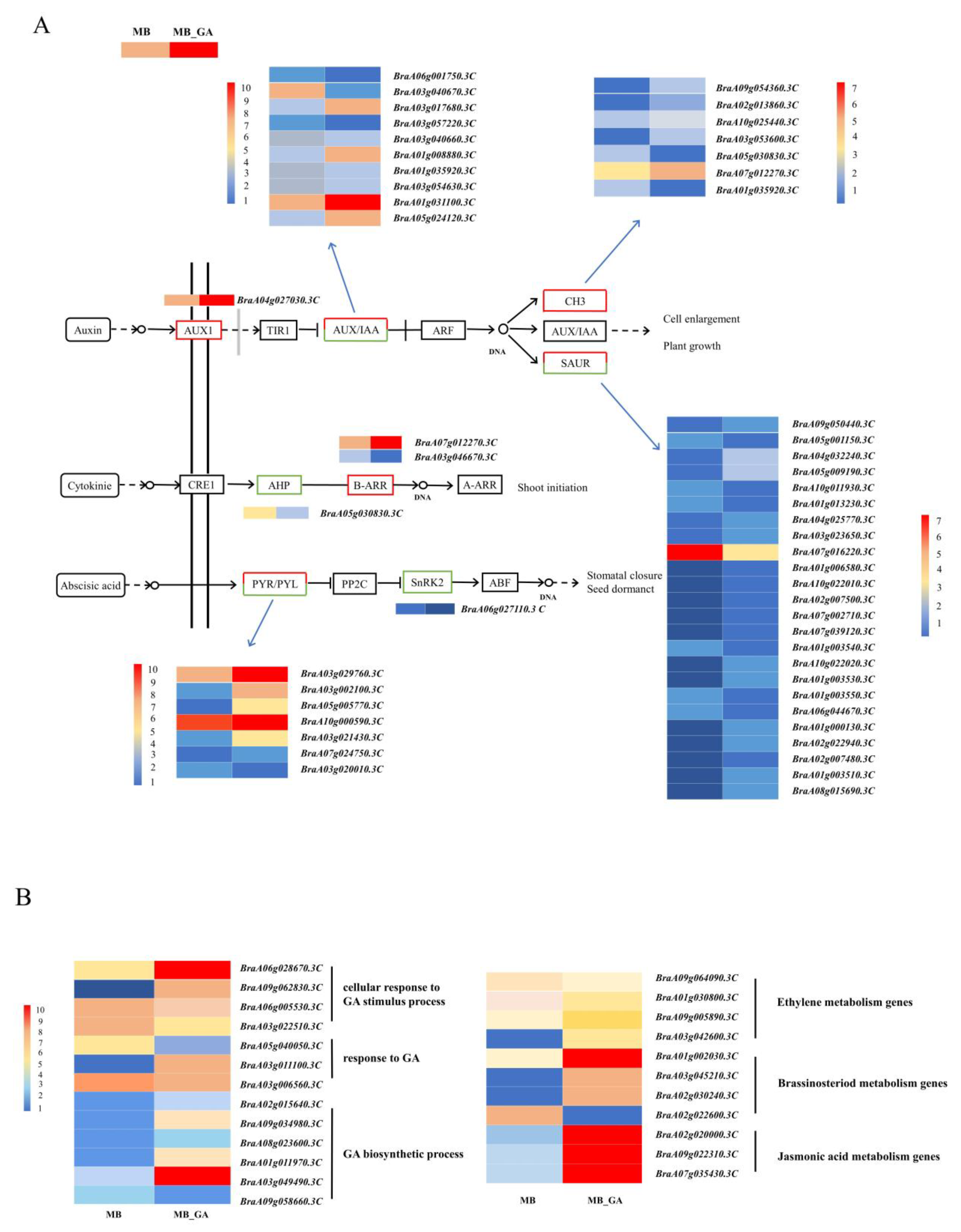
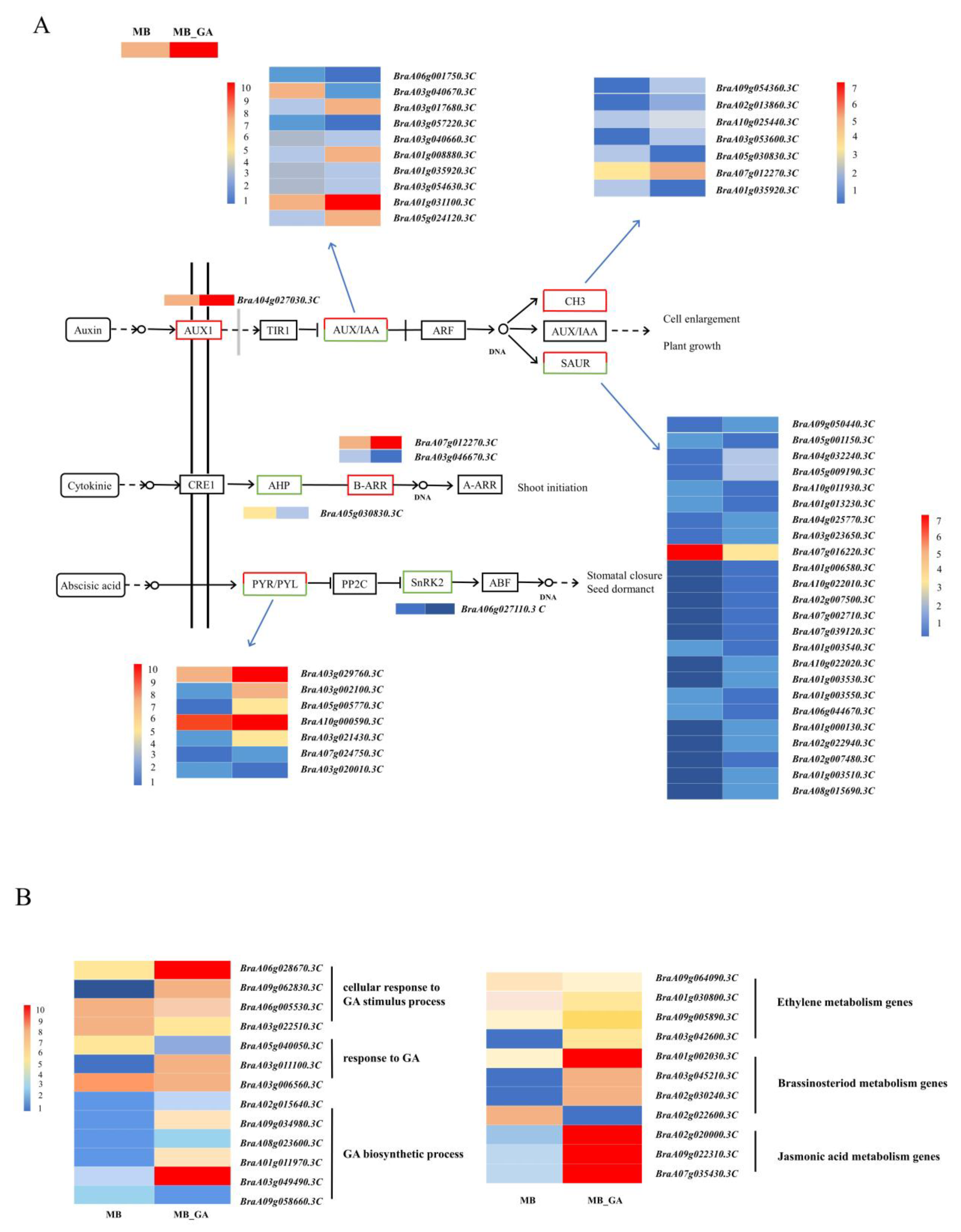
3.4. DEGs in Plant Hormones Related to Branching
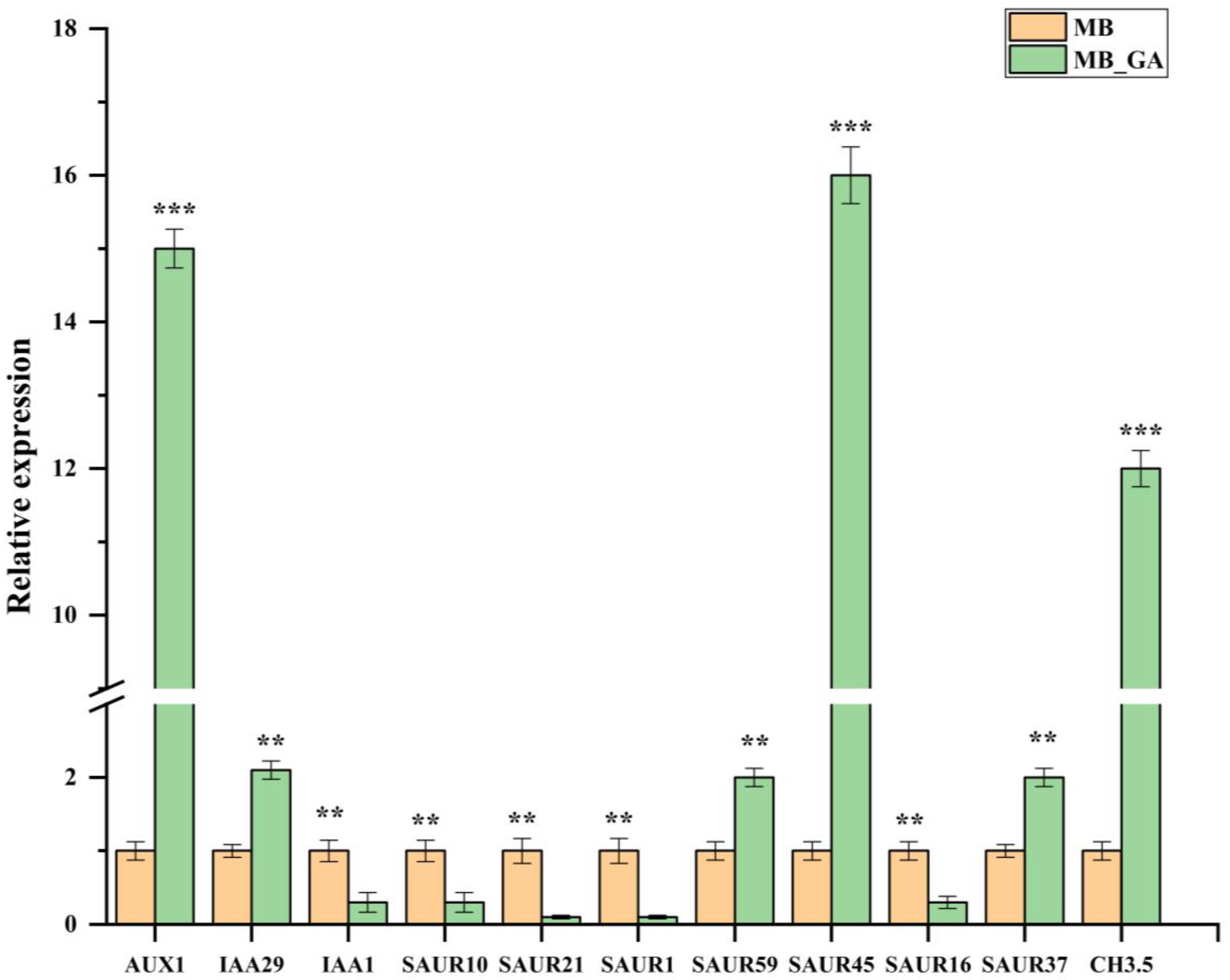
3.5. qRT-PCR Analysis on DEGs Related to Auxin
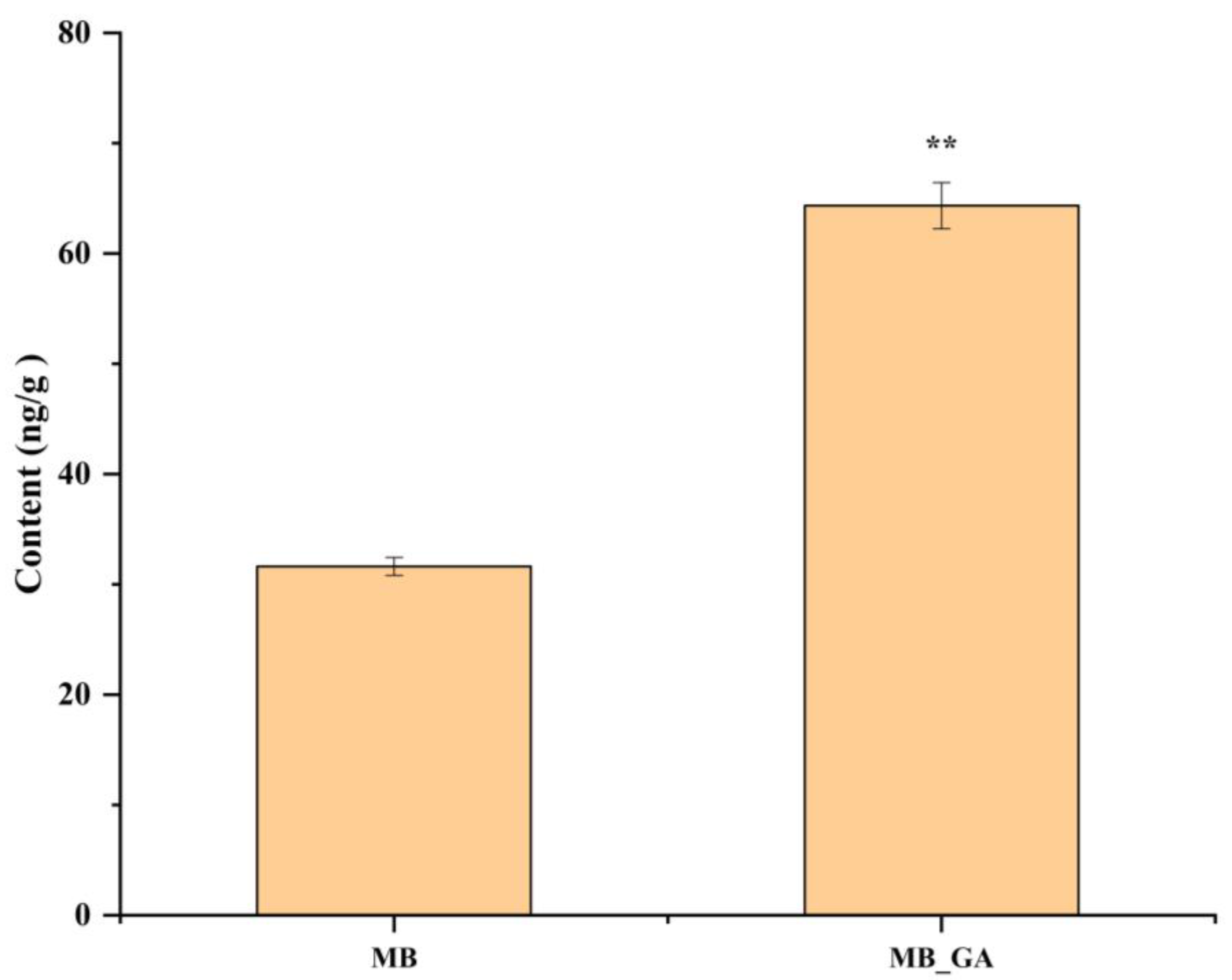
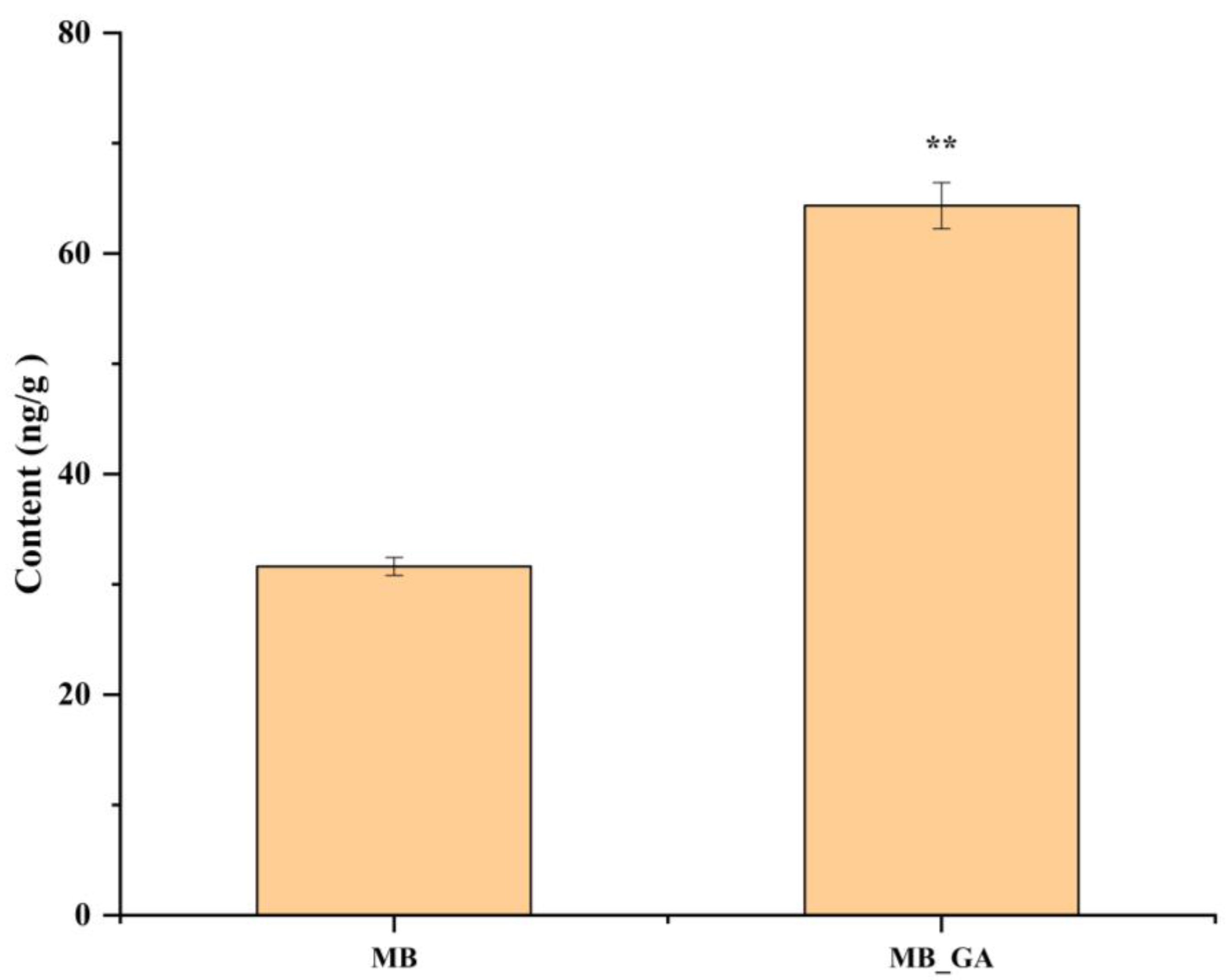
3.6. Exogenous GA3 Altered Auxin Contents in the Rosette Shoot
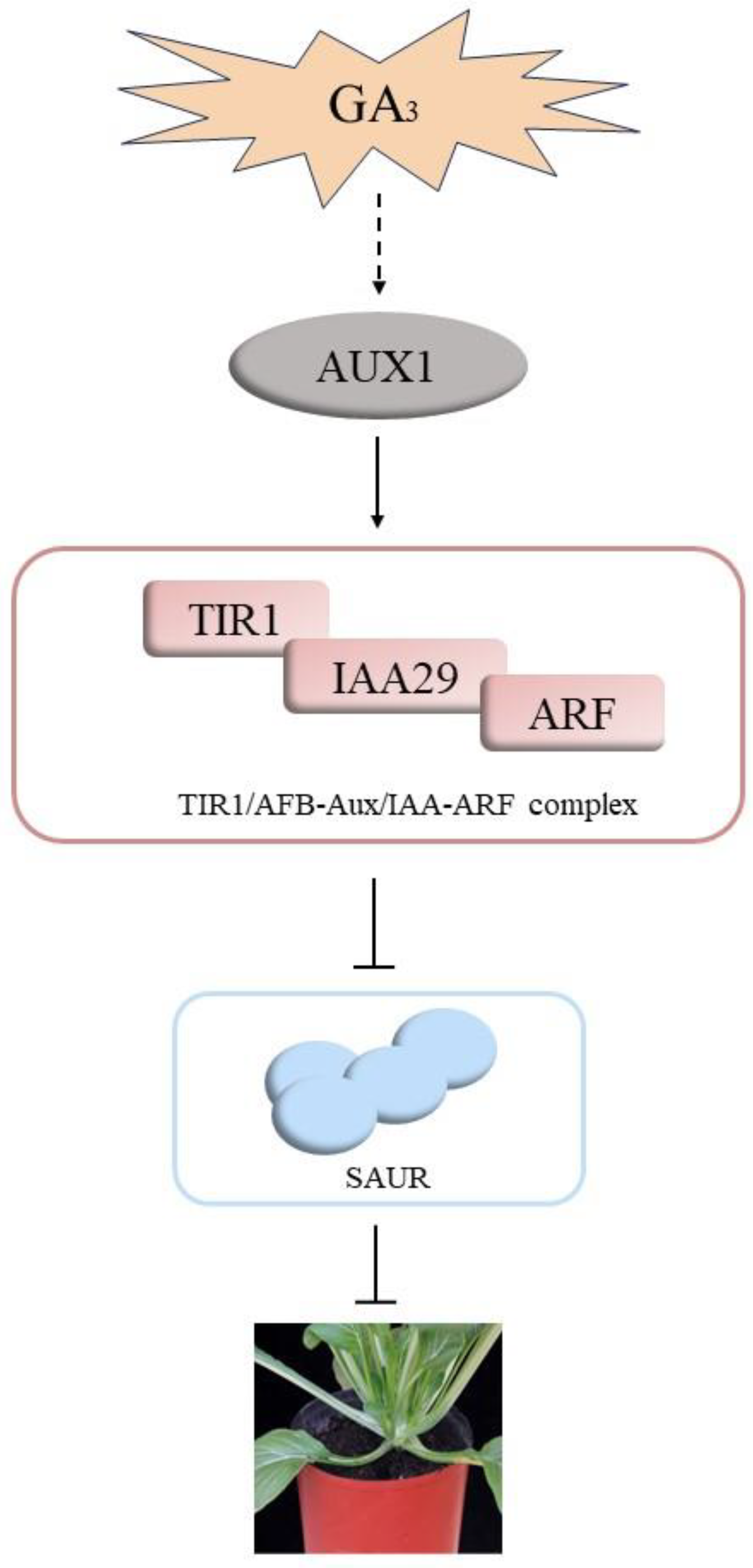
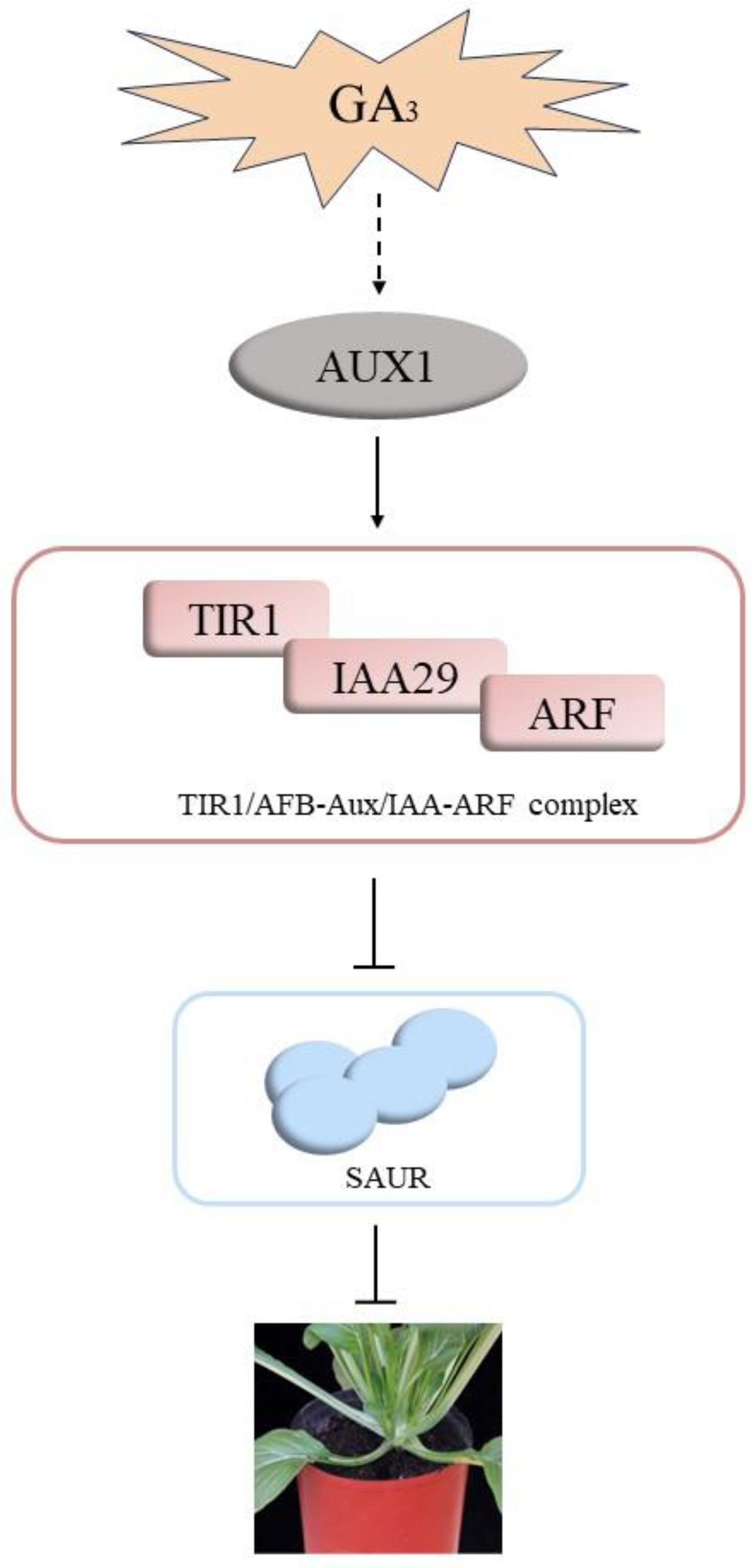
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lin, L.Z.; Harnly, J.M. Phenolic component profiles of mustard greens, yu choy, and 15 other brassica vegetables. J. Agric. Food Chem. 2010, 58, 6850–6857. [Google Scholar] [CrossRef] [PubMed]
- Teichmann, T.; Muhr, M. Shaping plant architecture. Front. Plant Sci. 2015, 6, 233. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Moigne, M.L.; Bertheloot, J.; Crespel, L.; Perez-Garcia, M.D.; Ogé, L.; Demotes-Mainard, S.; Hamama, L.; Davière, J.M.; Sakr, S. BRANCHED1: A Key Hub of Shoot Branching. Front. Plant Sci. 2019, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Nicolas, M.; Zhang, J.; Yu, H.; Guo, D.; Yuan, R.; Zhang, T.; Yang, J.; Cubas, P.; Qin, G. The TIE1 transcriptional repressor controls shoot branching by directly repressing BRANCHED1 in Arabidopsis. PLoS Genet. 2018, 14, e1007296. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.; Lee, D.K.; Choi, Y.D.; Kim, J.K. OsIAA6, a member of the rice Aux/IAA gene family, is involved in drought tolerance and tiller outgrowth. Plant Sci. 2015, 236, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Bainbridge, K.; Guyomarc’h, S.; Bayer, E.; Swarup, R.; Bennett, M.; Mandel, T.; Kuhlemeier, C. Auxin influx carriers stabilize phyllotactic patterning. Genes Dev. 2008, 22, 810–823. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, J.; Ding, Y.; Wang, Q.; Li, G.; Wang, S. Auxin inhibits the outgrowth of tiller buds in rice (Oryza sativa L.) by downregulating OsIPT expression and cytokinin biosynthesis in nodes. Am. J. Crop Sci. 2011, 5, 169–174. [Google Scholar]
- Reinhardt, D.; Pesce, E.R.; Stieger, P.; Mandel, T.; Baltensperger, K.; Bennett, M.; Traas, J.; Friml, J.; Kuhlemeier, C. Regulation of phyllotaxis by polar auxin transport. Nature 2003, 426, 255–260. [Google Scholar] [CrossRef]
- Vernoux, T.; Besnard, F.; Traas, J. Auxin at the shoot apical meristem. Cold Spring Harb. Perspect. Biol. 2010, 2, a001487. [Google Scholar] [CrossRef]
- Barbier, F.F.; Dun, E.A.; Kerr, S.C.; Chabikwa, T.G.; Beveridge, C.A. An Update on the Signals Controlling Shoot Branching. Trends Plant Sci. 2019, 24, 220–236. [Google Scholar] [CrossRef]
- Dierck, R.; Keyser, E.D.; Riek, J.D.; Dhooghe, E.; Huylenbroeck, J.V.; Prinsen, E.; Straeten, D.V.D. Change in Auxin and Cytokinin Levels Coincides with Altered Expression of Branching Genes during Axillary Bud Outgrowth in Chrysanthemum. PLoS ONE 2016, 11, e0161732. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.B.; Zhang, Z.; Wu, F.Q.; Feng, M.; Sun, Y.; Chen, W.W.; Cheng, Z.J.; Zhang, X.; Ren, Y.L.; Lei, C.l.; et al. The APC/CTE E3 Ubiquitin Ligase Complex Mediates the Antagonistic Regulation of Root Growth and Tillering by ABA and GA. Plant Cell 2020, 32, 1973–1987. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, S.; Zhu, W.; Jia, K.; Yang, H.; Wang, X. Strigolactone/MAX2-induced degradation of brassinoste-roid transcriptional effector BES1 regulates shoot branching. Dev. Cell 2013, 27, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S. Gibberellin metabolism and its regulation. Annu. Rev. Plant Biol. 2008, 59, 225–251. [Google Scholar] [CrossRef] [PubMed]
- Hedden, P.; Phillips, A.L. Gibberellin metabolism: New insights revealed by the genes. Trends Plant Sci. 2000, 5, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Eiichi, T.; Naohiko, Y.; Yoshio, M. Effect of Gibberellic Acid on Dwarf and Normal Pea Plants. Physiol. Plant. 1967, 20, 291–298. [Google Scholar] [CrossRef]
- Tan, M.; Li, G.; Liu, X.; Cheng, F.; Ma, J.; Zhao, C.; Zhang, D.; Han, M. Exogenous application of GA3 inactively regulates axillary bud outgrowth by influencing of branching-inhibitors and bud-regulating hormones in apple (Malus domestica Borkh.). Mol. Genet. Genomics 2018, 293, 1547–1563. [Google Scholar] [CrossRef]
- Tian, Y.T.; Wang, J.N.; Guo, H.P.; Qu, K.; Xu, D.; Hou, L.L.; Li, J.H. Transcriptome Analysis of Active Axillary Buds from Narrow-crown and Broad-crown Poplars Provides Insight into the Phytohormone Regulatory Network for Branching Angle. Plant Mol. Biol. Report. 2021, 39, 595–606. [Google Scholar] [CrossRef]
- Zhang, Q.Q.; Wang, J.G.; Wang, L.Y.; Wang, J.F.; Wang, Q.; Yu, P.; Bai, M.Y.; Fan, M. Gibberellin repression of axillary bud formation in Arabidopsis by modulation of DELLA-SPL9 complex activity. J. Integr. Plant Biol. 2020, 62, 421–432. [Google Scholar] [CrossRef]
- Lo, S.F.; Yang, S.Y.; Chen, K.T.; Hsing, Y.I.; Zeevaart, J.A.; Chen, L.J.; Yu, S.M. A novel class of gibberellin 2-oxidases control semidwarfism, tillering, and root development in rice. Plant Cell 2008, 20, 2603–2618. [Google Scholar] [CrossRef]
- Liao, Z.; Yu, H.; Duan, J.; Yuan, K.; Yu, C.; Meng, X.; Kou, L.; Chen, M.; Jing, Y.; Liu, G.; et al. SLR1 inhibits MOC1 degradation to coordinate tiller number and plant height in rice. Nat. Commun. 2019, 10, 2738. [Google Scholar] [CrossRef]
- Mauriat, M.; Sandberg, L.G.; Moritz, M. Proper gibberellin localization in vascular tissue is required to control auxin-dependent leaf development and bud outgrowth in hybrid aspen. Plant J. 2011, 67, 805–816. [Google Scholar] [CrossRef]
- Agharkar, M.; Lomba, P.; Altpeter, F.; Zhang, H.; Kenworthy, K.L.T. Stable expression of AtGA2ox1 in a low-input turfgrass (Paspalum notatum Flugge) reduces bioactive gibberellin levels and improves turf quality under field conditions. Plant Biotechnol. J. 2007, 5, 791–801. [Google Scholar] [CrossRef]
- Caruana, J.C.; Sittmann, J.W.; Wang, W.P.; Liu, Z.C. Suppressor of Runnerless encodes a DELLA protein that controls runner formation for asexual reproduction in strawberry. Mol. Plant 2018, 11, 230–233. [Google Scholar] [CrossRef]
- O’Neill, D.P.; Ross, J.J. Auxin regulation of the gibberellin pathway in pea. Plant Physiol. 2000, 130, 1974–1982. [Google Scholar] [CrossRef]
- Tenreira, T.; Lange, M.J.P.; Lange, T.; Bres, C.; Labadie, M.; Monfort, A.; Hernould, M.; Rothan, C.; Denoyes, B. A Specific Gibberellin 20-Oxidase Dictates the Flowering-Runnering Decision in Diploid Strawberry. Plant Cell 2017, 29, 2168–2182. [Google Scholar] [CrossRef]
- Elfving, D.; Visser, D.; Henry, J. Gibberellins stimulate lateral branch development in young sweet cherry trees in the orchard. Int. J. Fruit Sci. 2011, 11, 41–54. [Google Scholar] [CrossRef]
- Choubane, D.; Rabot, A.; Mortreau, E.; Legourrierec, J.; Péron, T.; Foucher, F.; Ahcène, Y.; Pelleschi-Trvier, S.; Leduc, N.; Hamama, L. Photocontrol of bud burst involves gibberellin biosynthesis in Rosa sp. J. Plant Physiol. 2012, 169, 1271–1280. [Google Scholar] [CrossRef]
- Katyayini, N.U.; Rinne, P.L.H.; Tarkowská, D.; Strnad, M.; van der Schoot, C. Dual Role of Gibberellin in Perennial Shoot Branching: Inhibition and Activation. Front. Plant Sci. 2020, 11, 529247. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.; Li, J.; Yao, Q.; Liu, Z.; Feng, H.; Zhang, Y. Identification of two tandem genes associated with primary rosette branching in flowering Chinese cabbage. Front. Plant Sci. 2022, 19, 1083528. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.P.; Tseng, E.; Salamov, A.; Zhang, J.; Meng, X.; Zhao, Z.; Wang, Z. Widespread Polycistronic Transcripts in Fungi Revealed by Single-Molecule mRNA Sequencing. PLoS ONE 2015, 10, e0132628. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Rosa, M.; Hilal, M.; González, J.A.; Prado, F.E. Low-temperature effect on enzyme activities involved in sucrose-starch partitioning in salt-stressed and salt-acclimated cotyledons of quinoa (Chenopodium quinoa Willd.) seedlings. Plant Physiol. Biochem. 2019, 47, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR. Methods 2002, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Benjamini, Y.; Yekutieli, D. The Control of the False Discovery Rate in Multiple Testing under Dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar] [CrossRef]
- Abdi, H. Bonferroni and Šidák corrections for multiple comparisons. In Encyclopedia of Measurement and Statistics; Salkind, N., Ed.; CiteSeerX: Princeton, NJ, USA, 2007; pp. 103–107. [Google Scholar]
- Yuan, C.; Xi, L.; Kou, Y.; Zhao, Y.; Zhao, L. Current perspectives on shoot branching regulation. Front. Agric. Sci. Eng. 2015, 2, 38–52. [Google Scholar] [CrossRef]
- Lee, M.S.; An, J.H.; Cho, H.T. Biological and molecular functions of two EAR motifs of Arabidopsis IAA7. J. Plant Biol. 2016, 59, 24–32. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Q.; Ding, Y.; Li, G.H.; Xu, J.X.; Wang, S.H. Effects of external ABA, GA3 and NAA on the tiller bud outgrowth of rice is related to changes in endogenous hormones. Plant Growth Regul. 2011, 65, 247–254. [Google Scholar] [CrossRef]
- Cui, D.; Neill, S.J.; Tang, Z.; Cai, W. Gibberellin-regulated XET is differentially induced by auxin in rice leaf sheath bases during gravitropic bending. J. Exp. Bot. 2005, 56, 1327–1334. [Google Scholar] [CrossRef]
- Fukazawa, J.; Ito, T.; Kamiya, Y.; Yamaguchi, S.; Takahashi, Y. Binding of GID1 to DELLAs promotes dissociation of GAF1 from DELLA in GA dependent manner. Plant Signal. Behav. 2015, 10, e1052923. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Li, L.; Zhou, Q.Y.; Liu, W.Z.; Zhang, H.X.; Chen, W.Z.; Ding, Y.F. Expression of wheat gibberellins 2-oxidase gene induced dwarf or semi-dwarf phenotype in rice. Cereal Res. Commun. 2019, 47, 239–249. [Google Scholar] [CrossRef]
- Nadeau, C.D.; Ozga, J.A.; Kurepin, L.V.; Jin, A.; Pharis, R.P.; Reinecke, D.M. Tissue-Specific Regulation of Gibberellin Biosynthesis in Developing Pea Seeds. Plant Physiol. 2011, 156, 897–912. [Google Scholar] [CrossRef] [PubMed]
- Worley, C.K.; Zenser, N.; Ramos, J.; Rouse, D.; Leyser, O.; Theologis, A.; Callis, J. Degradation of AUX⁄IAA proteins is essential for normal auxinsignalling. Plant J. 2000, 21, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.B.; Wang, X.J.; Hagen, G.; Guilfoyle, T.J. AUX⁄IAA proteins areactive repressors, and their stability and activity are modulated by auxin. Plant Cell 2001, 13, 2809–2822. [Google Scholar] [CrossRef] [PubMed]
- Dharmasiri, N.; Dharmasiri, S.; Estelle, M. The F-box protein TIR1 is anauxin receptor. Nature 2005, 435, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Gray, W.M.; Kepinski, S.; Rouse, D.; Leyser, O.; Estelle, M. Auxin regulates SCF(TIR1)-dependent degradation of AUX⁄IAA proteins. Nature 2001, 414, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Nijhawan, A.; Arora, R.; Agarwal, P.; Ray, S.; Sharma, P.; Kapoor, S.; Tyagi, A.K.; Khurana, J.P. F-box proteins in rice. Genome-wide analysis, classification, temporal and spatial gene expression during panicle and seeddevelopment, and regulation by light and abiotic stress. Plant Physiol. 2007, 143, 1467–1483. [Google Scholar] [CrossRef]
- Trenner, J.; Poeschl, Y.; Grau, J.; Gogol-Döring, A.; Quint, M.; Delker, C. Auxin-induced expression divergence between Arabidopsis species may originate within the TIR1/AFB-AUX/IAA-ARF module. J. Exp. Bot. 2017, 68, 539–552. [Google Scholar] [CrossRef]
- Spartz, A.K.; Lee, S.H.; Wenger, J.P.; Gonzalez, N.; Itoh, H.; Inzé, D.; Peer, W.A.; Murphy, A.S.; Overvoorde, P.J.; Gray, W.M. The SAUR19 subfamily of SMALL AUXIN UP RNA genes promote cell expansion. Plant J. 2012, 70, 978–1068. [Google Scholar] [CrossRef]
- Lee, M.S.; Choi, H.-S.; Cho, H.-T. Branching the auxin signaling; Multiple players and diverse interactions. J. Plant Biol. 2013, 56, 130–137. [Google Scholar] [CrossRef]
- Pattison, R.J.; Catala, C. Evaluating auxin distribution in tomato (Solanum lycopersicum) through an analysis of the PIN and AUX/LAX gene families. Plant J. 2012, 70, 585–598. [Google Scholar] [CrossRef] [PubMed]
- Vanneste, S.; Friml, J. Auxin: A trigger for change in plant development. Cell 2009, 36, 005–016. [Google Scholar] [CrossRef] [PubMed]
- Salanenka, Y.; Verstraeten, I.; Löfke, C.; Tabata, K.; Naramoto, S.; Glanc, M.; Friml, J. Gibberellin DELLA signaling targets the retromer complex to redirect protein trafficking to the plasma membrane. Proc. Natl. Acad. Sci. USA 2018, 115, 3716–3721. [Google Scholar] [CrossRef] [PubMed]
- Harrison, B.R.; Masson, P.H. ARL2, ARG1 and PIN3 define a gravity signal transduction pathway in root statocytes. Plant J. 2008, 53, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Gou, J.; Fu, C.; Liu, S.; Tang, C.; Debnath, S.; Flanagan, A.; Ge, Y.; Tang, Y.; Jiang, Q.; Larson, P.R.; et al. The miR156-SPL4 module predominantly regulates aerial axillary bud formation and controls shoot architecture. New Phytol. 2017, 216, 829–840. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.H.; Chang, S.; Wang, H.M.; Chu, Z.L.; Xi, C.; Liu, J.; Zhao, H.P.; Han, S.C.; Wang, Y.D. Transcriptomic analysis reveals the contribution of auxin on the differentially developed caryopses on primary and secondary branches in rice. J. Plant Physiol. 2021, 256, 153310. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.; He, S.; Bian, S.; Han, X.; Liu, K.; Cong, P. Genome-wide identification and expression analysis of major latex protein (MLP) family genes in the apple (Malus domestica borkh.) genome. Gene 2020, 733, 144275. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.W. Auxin response factors. Plant Cell Environ. 2016, 39, 1014–1028. [Google Scholar] [CrossRef]
- Singh, K.B.; Foley, R.C.; Oñate-Sánchez, L. Transcription factors in plant defense and stress responses. Curr. Opin. Plant Biol. 2002, 5, 430–436. [Google Scholar] [CrossRef]
- Boter, M.; Ruiz-Rivero, O.; Abdeen, A.; Prat, S. Conserved MYC transcription factors play a key role in jasmonate signaling both in tomato and Arabidopsis. Genes 2004, 18, 1577–1591. [Google Scholar] [CrossRef]
- Wu, Q.; Bai, J.; Tao, X.; Mou, W.; Luo, Z.; Mao, L. Synergistic effect of abscisic acid and ethylene on color development in tomato (Solanum lycopersicum L.) fruit. Sci. Hortic. 2018, 235, 169–180. [Google Scholar] [CrossRef]
- Yang, C.; Shi, G.; Li, Y.; Luo, M.; Wang, H.; Wang, J.; Yuan, L.; Wang, Y.; Li, Y. Genome-Wide Identification of SnRK1 Catalytic α Subunit and FLZ Proteins in Glycyrrhiza inflata Bat. Highlights Their Potential Roles in Licorice Growth and Abiotic Stress Responses. Int. J. Mol. Sci. 2023, 24, 121. [Google Scholar] [CrossRef]
- Pan, W.; Zheng, P.; Zhang, C.; Wang, W.; Li, Y.; Fan, T. The effect of ABRE BINDING FACTOR 4-mediated FYVE1 on salt stress tolerance in arabidopsis. Plant Sci. 2020, 296, 110489. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, L.; Du, H.; Dong, Y.; Zhen, S.; Wang, C.; Wang, Q.; Yang, J.; Zhang, P.; Zheng, X.; et al. An ARF24-ZmArf2 module influences kernel size in different maize haplotypes. J. Integr. Plant Biol. 2023, 65, 1767–1781. [Google Scholar] [CrossRef]
- Keita, S.; Daisuke, Y. Two cis-acting elements necessary and sufficient for gibberellin-upregulated proteinase expression in rice seeds. Plant J. 2003, 34, 635–645. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway ID | Pathway | DEG (%) | Total DEGs | Adjust p Value |
|---|---|---|---|---|
| brp04075 | Plant hormone signal transduction | 74 (9.60%) | 771 | 1.73 × 10−2 |
| brp00940 | Phenylpropanoid biosynthesis | 51 (6.61%) | 771 | 6.20 × 10−4 |
| brp00500 | Starch and sucrose metabolism | 39 (5.06%) | 771 | 2.91 × 10−3 |
| brp04626 | Plant–pathogen interaction | 38 (4.93%) | 771 | 3.54 × 10−1 |
| brp04016 | MAPK signaling pathway—plant | 32 (4.15%) | 771 | 6.81 × 10−2 |
| brp00710 | Carbon fixation in photosynthetic organisms | 30 (3.90%) | 771 | 2.24 × 10−4 |
| brp00270 | Cysteine and methionine metabolism | 29 (3.76%) | 771 | 4.43 × 10−2 |
| brp04146 | Peroxisome | 27 (3.50%) | 771 | 1.95 × 10−2 |
| brp00480 | Glutathione metabolism | 26 (3.37%) | 771 | 1.95 × 10−2 |
| brp00630 | Glyoxylate and dicarboxylate metabolism | 24 (3.11%) | 771 | 3.32 × 10−2 |
| brp00230 | Purine metabolism | 24 (3.11%) | 771 | 3.19 × 10−1 |
| brp00195 | Photosynthesis | 23 (2.98%) | 771 | 8.07 × 10−4 |
| brp00260 | Glycine, serine, and threonine metabolism | 21 (2.72%) | 771 | 5.77 × 10−2 |
| brp00040 | Pentose and glucuronate interconversions | 21 (2.72%) | 771 | 4.72 × 10−1 |
| brp00360 | Phenylalanine metabolism | 20 (2.60%) | 771 | 1.71 × 10−3 |
| brp00380 | Tryptophan metabolism | 20 (2.60%) | 771 | 2.16 × 10−3 |
| brp00130 | Ubiquinone and other terpenoid–quinone biosynthesis | 16 (2.08%) | 771 | 3.71 × 10−2 |
| brp00561 | Glycerolipid metabolism | 16 (2.08%) | 771 | 2.49 × 10−1 |
| brp00966 | Glucosinolate biosynthesis | 15 (1.95%) | 771 | 1.41 × 10−7 |
| brp00906 | Carotenoid biosynthesis | 15 (1.95%) | 771 | 8.07 × 10−4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, X.; Zhao, Y.; Cai, N.; Guan, J.; Liu, Z.; Liu, Z.; Feng, H.; Zhang, Y. Characterization and Transcriptome Analysis Reveal Exogenous GA3 Inhibited Rosette Branching via Altering Auxin Approach in Flowering Chinese Cabbage. Agronomy 2024, 14, 762. https://doi.org/10.3390/agronomy14040762
Qi X, Zhao Y, Cai N, Guan J, Liu Z, Liu Z, Feng H, Zhang Y. Characterization and Transcriptome Analysis Reveal Exogenous GA3 Inhibited Rosette Branching via Altering Auxin Approach in Flowering Chinese Cabbage. Agronomy. 2024; 14(4):762. https://doi.org/10.3390/agronomy14040762
Chicago/Turabian StyleQi, Xinghua, Ying Zhao, Ningning Cai, Jian Guan, Zeji Liu, Zhiyong Liu, Hui Feng, and Yun Zhang. 2024. "Characterization and Transcriptome Analysis Reveal Exogenous GA3 Inhibited Rosette Branching via Altering Auxin Approach in Flowering Chinese Cabbage" Agronomy 14, no. 4: 762. https://doi.org/10.3390/agronomy14040762
APA StyleQi, X., Zhao, Y., Cai, N., Guan, J., Liu, Z., Liu, Z., Feng, H., & Zhang, Y. (2024). Characterization and Transcriptome Analysis Reveal Exogenous GA3 Inhibited Rosette Branching via Altering Auxin Approach in Flowering Chinese Cabbage. Agronomy, 14(4), 762. https://doi.org/10.3390/agronomy14040762