Final Seed Size in Soybean Is Determined during Mid-Seed Filling Stage


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Crop Husbandry
2.2. Experimental Design and Treatments
2.3. Observations
2.3.1. Temporal Seed Growth Dynamics under Control Conditions
2.3.2. Final Seed Weight, Pod Weight, Seed Filling Duration and Rate
2.3.3. Seed Composition
2.4. Statistical Analysis
3. Results
3.1. Climatic Conditions during Plant Growth
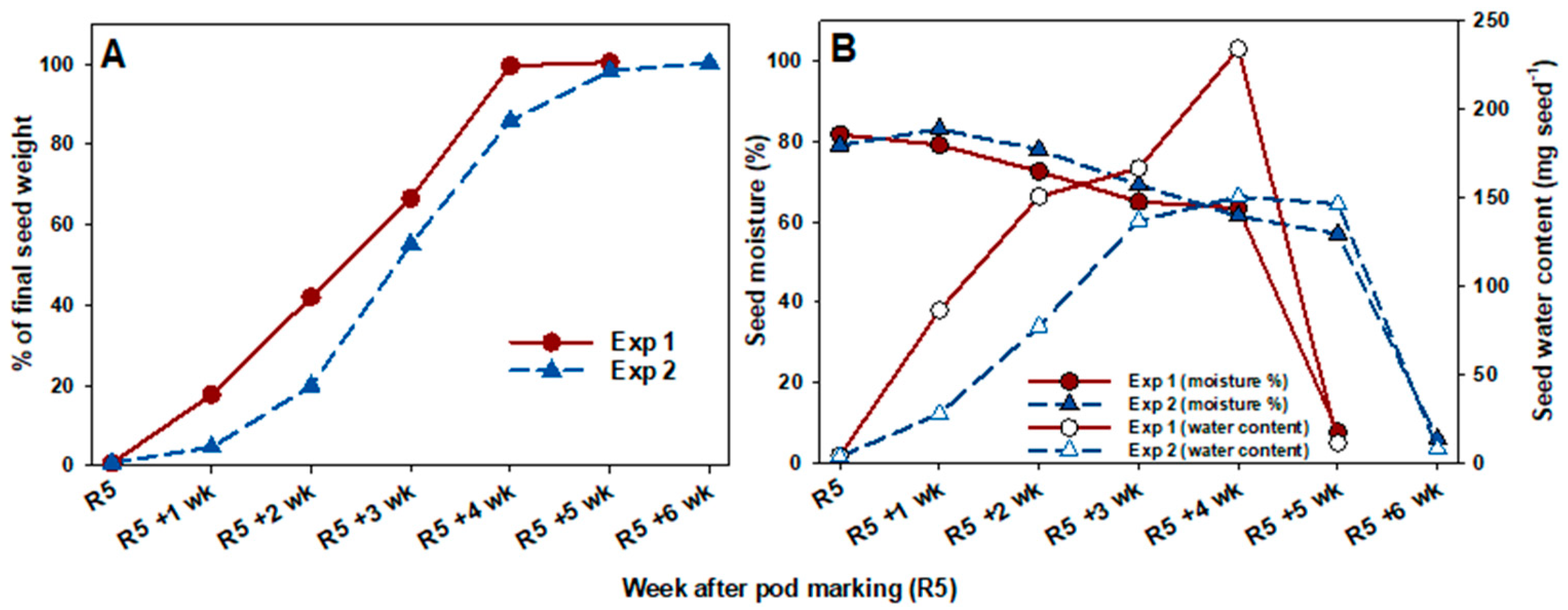
3.2. Seed Weight and Moisture Dynamics under Control
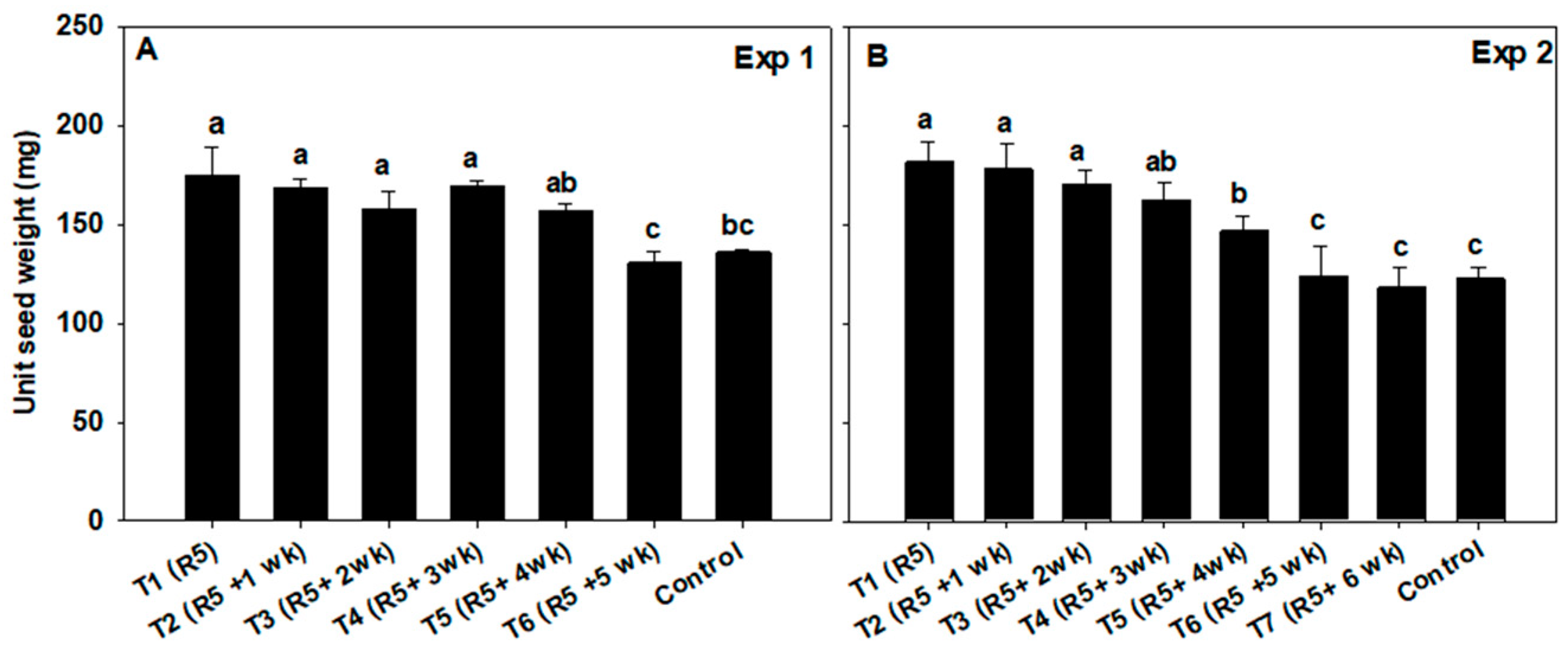
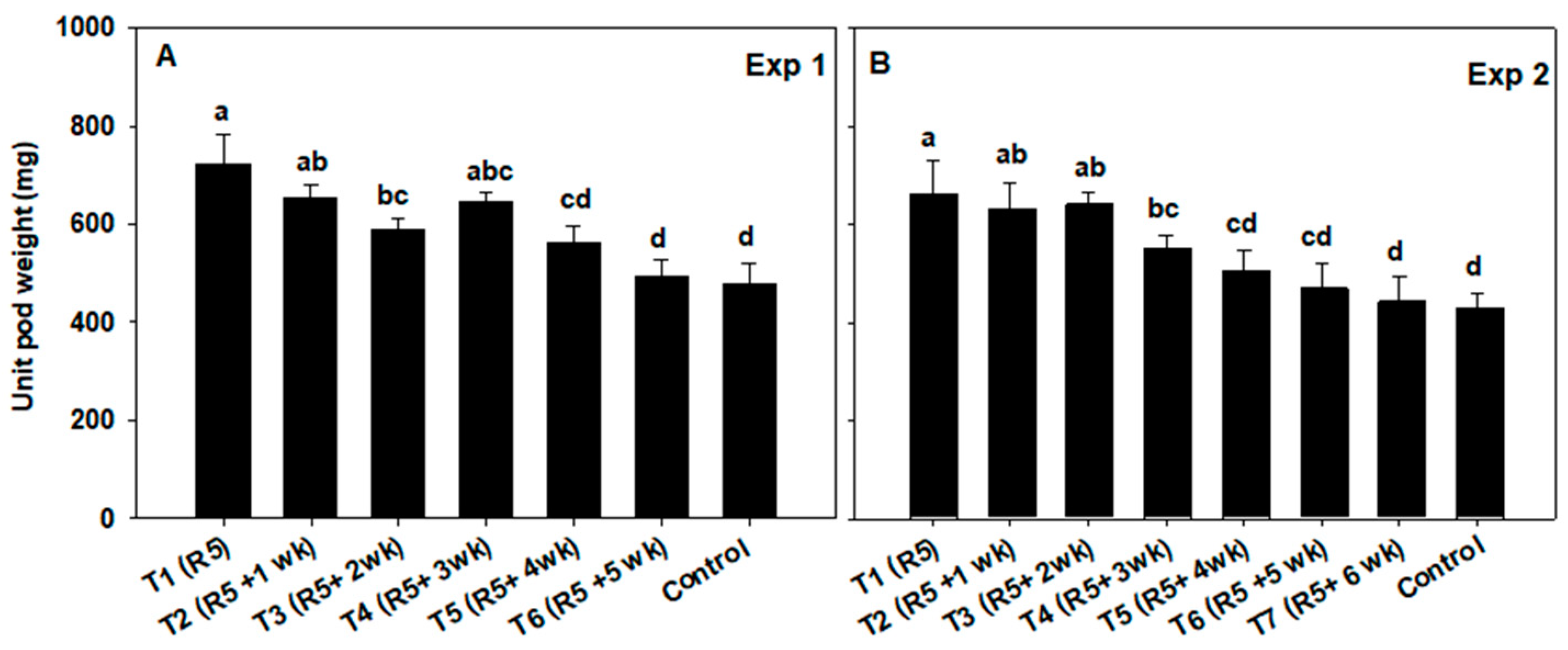
3.3. Seed and Pod Weight
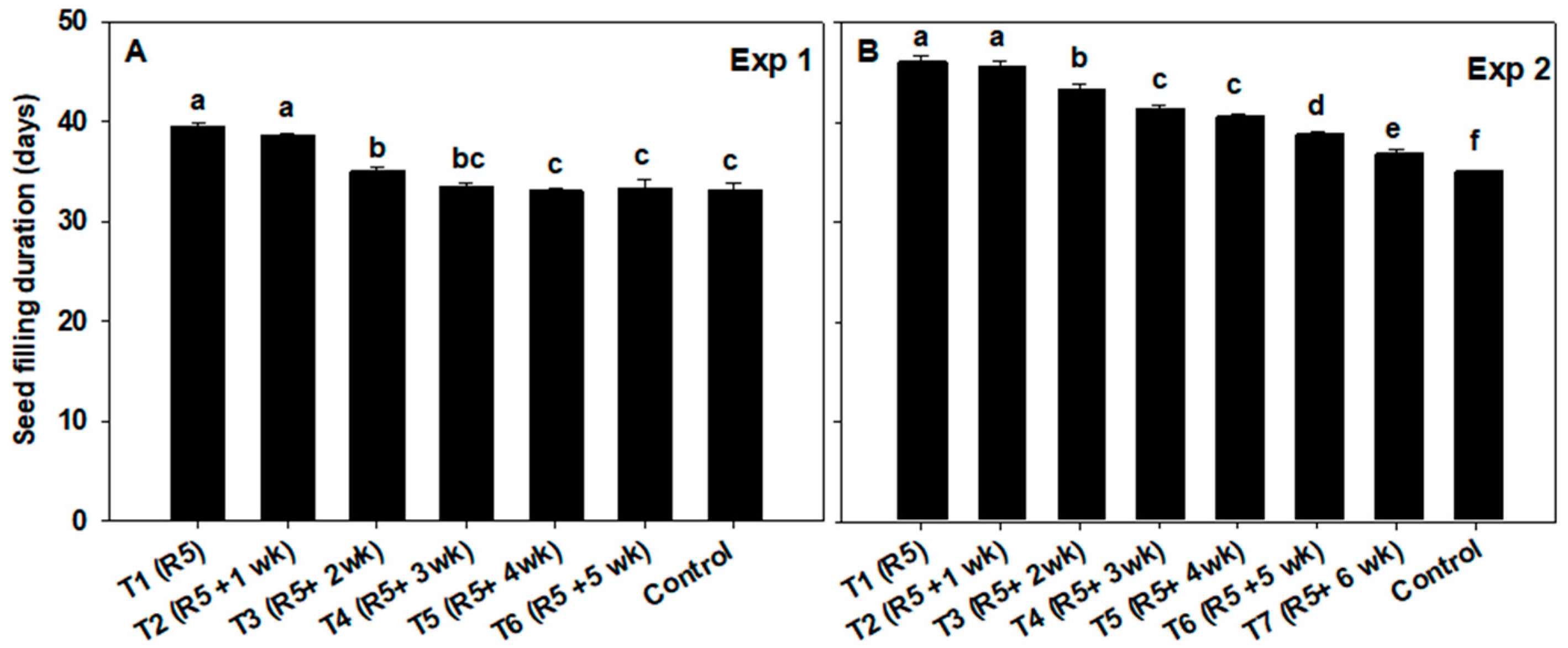
3.4. Seed Filling Duration
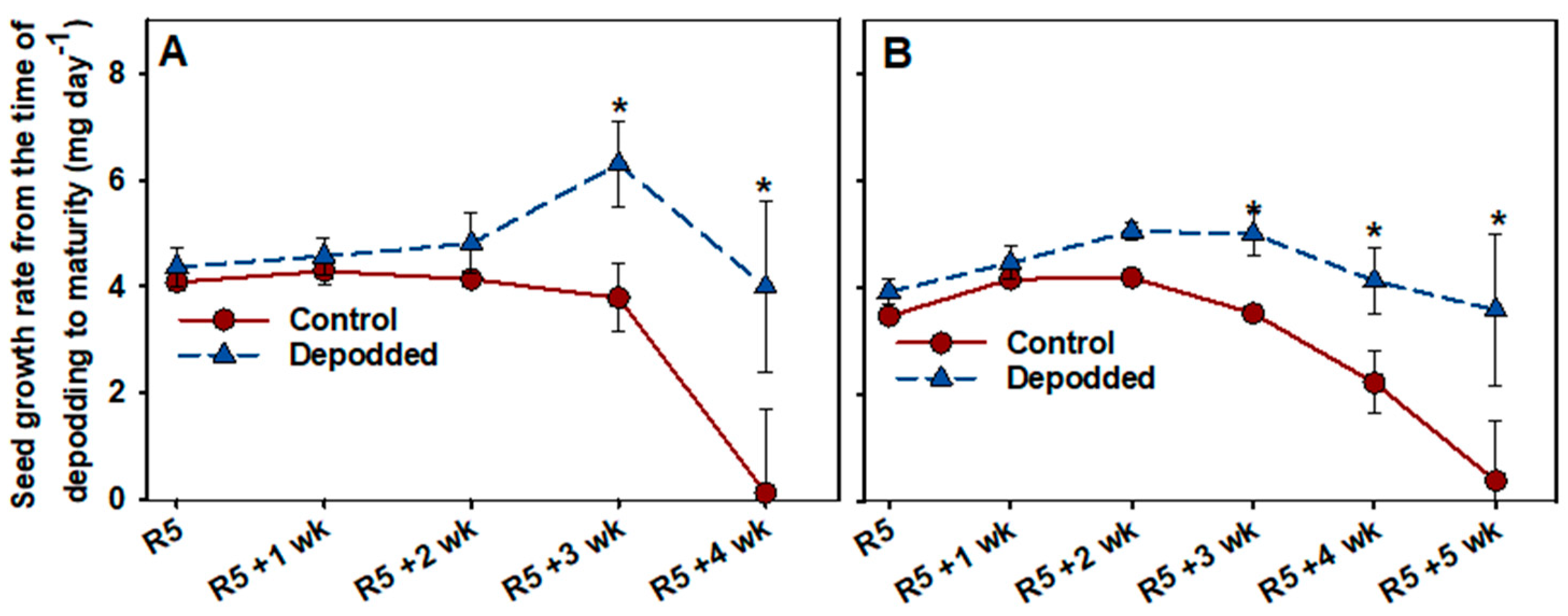
3.5. Seed Growth Rate
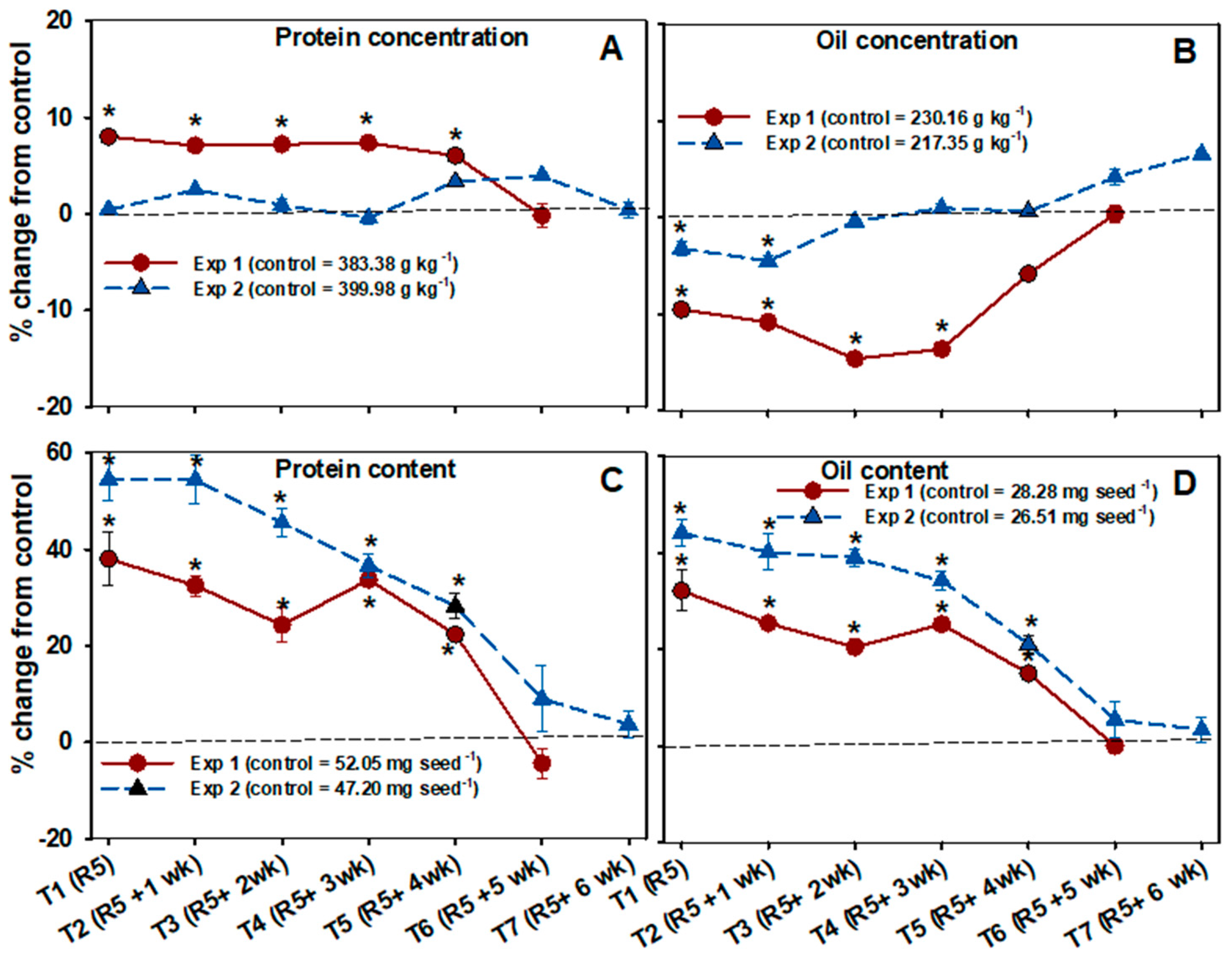
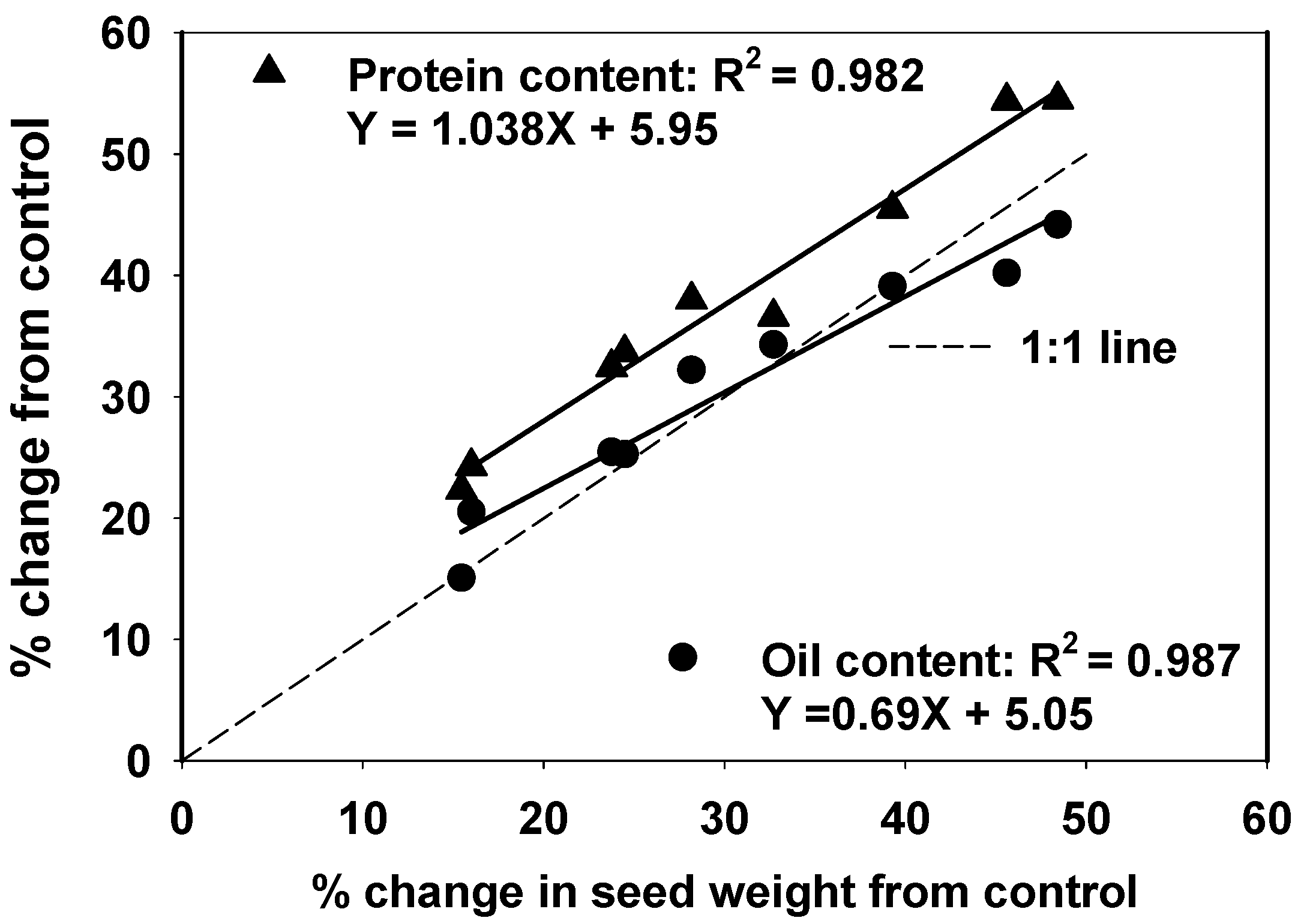
3.6. Seed Composition
4. Discussion
4.1. Source Limitation in Soybean and Its Implication
4.2. Soybean Respond to Increases in Assimilate Supply until Mid-Seed Filling Stage under Field Conditions
4.3. Increased Seed Filling Duration and Uniform Seed Filling Rate over Extended Period Associated with Higher Seed Weight under Increased Assimilate Supply
4.4. Protein Respond More Favorably under Increased Assimilate Supply Than Other Seed Constituents
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, M.R.; Rao, I.M.; Merchant, A. Source-Sink Relationships in Crop Plants and Their Influence on Yield Development and Nutritional Quality. Front Plant Sci 2018, 9, 420465. [Google Scholar] [CrossRef] [PubMed]
- Borrás, L.; Slafer, G.A.; Otegui, M.E. Seed dry weight response to source–sink manipulations in wheat, maize and soybean: A quantitative reappraisal. Field Crops Res. 2004, 86, 131–146. [Google Scholar] [CrossRef]
- Ordóñez, R.A.; Savin, R.; Cossani, C.M.; Slafer, G.A. Maize Grain Weight Sensitivity to Source–Sink Manipulations under a Wide Range of Field Conditions. Crop Sci. 2018, 58, 2542–2557. [Google Scholar] [CrossRef]
- Sheehy, J.E.; Dionora, M.J.A.; Mitchell, P.L. Spikelet numbers, sink size and potential yield in rice. Field Crops Res. 2001, 71, 77–85. [Google Scholar] [CrossRef]
- Yoshida, S. Fundamentals of Rice Crop Science; International Rice Research Institute: Manila, Phillipines, 1981. [Google Scholar]
- Yang, Z.; van Oosterom, E.J.; Jordan, D.R.; Hammer, G.L. Pre-anthesis ovary development determines genotypic differences in potential kernel weight in sorghum. J. Exp. Bot. 2009, 60, 1399–1408. [Google Scholar] [CrossRef]
- Fortescue, J.A.; Turner, D.W. Changes in seed size and oil accumulation in Brassica napus L. by manipulating the source–sink ratio and excluding light from the developing siliques. Aust. J. Agric. Res. 2007, 58, 413–424. [Google Scholar] [CrossRef]
- Faraji, A. Seed weight in canola as a function of assimilate supply and source-sink ratio during seed filling period. Int. J. Plant Prod. 2014, 8, 1735. [Google Scholar]
- Zhang, H.; Flottmann, S. Source-sink manipulations indicate seed yield in canola is limited by source availability. European J. Agron. 2018, 96, 70–76. [Google Scholar] [CrossRef]
- Howlader, M.H.K.; Bain, S.K.; Hasan, M.M.; Khan, A.; Biswas, S. Source-sink manipulation on yield contributing characters and yield of Sesame (Sesamum indicum L.). Progress. Agric. 2018, 29, 1–9. [Google Scholar] [CrossRef]
- Iglesias, F.M.; Miralles, D.J. Changes in seed weight in response to different sources: Sink ratio in oilseed rape. J. Agric. Res. Innov. Technol. 2014, 4, 44–52. [Google Scholar] [CrossRef]
- Alkio, M.; Schubert, A.; Diepenbrock, W.; Grimm, E. Effect of source-sink ratio on seed set and filling in sunflower (Helianthus annuus L.). Plant Cell Env. 2003, 26, 1609–1619. [Google Scholar] [CrossRef]
- Sandana, P.; Calderini, D.F. Comparative assessment of the critical period for grain yield determination of narrow-leafed lupin and pea. Eur. J. Agron. 2012, 40, 94–101. [Google Scholar] [CrossRef]
- Binnie, R.C.; Clifford, P.E. Effects of Some Defoliation and Decapitation Treatments on the Productivity of French Beans. Ann. Bot. 1980, 46, 811–813. [Google Scholar] [CrossRef]
- Clifford, P.E. Source Limitation of Sink Yield in Mung Beans. Ann. Bot. 1979, 43, 397–399. [Google Scholar] [CrossRef]
- Chapman, G.P.; Guest, H.L.; Peat, W.E. Top-removal in single stem plants of Vicia faba L. Z. Für Pflanzenphysiol. 1978, 89, 119–127. [Google Scholar] [CrossRef]
- McAlister, D.F.; Krober, O.A. Response of Soybeans to Leaf and Pod Removal. Agron. J. 1958, 50, 674–677. [Google Scholar] [CrossRef]
- Lawn, R.J.; Brun, W.A. Symbiotic Nitrogen Fixation in Soybeans. I. Effect of Photosynthetic Source-Sink Manipulations. Crop Sci. 1974, 14, 11–16. [Google Scholar] [CrossRef]
- Egli, D.B.; Leggett, J.E. Rate of Dry Matter Accumulation in Soybean Seeds with Varying Source-Sink Ratios. Agron. J. 1976, 68, 371–374. [Google Scholar] [CrossRef]
- Openshaw, S.J.; Hadley, H.H.; Brokoski, C.E. Effects of POD Removal Upon Seeds of Nodulating and Nonnodulating Soybean Line. Crop Sci. 1979, 19, 289. [Google Scholar] [CrossRef]
- Egli, D.B.; Guffy, R.D.; Meckel, L.W.; Leggett, J.E. The Effect of Source-Sink Alterations on Soybean Seed Growth. Ann. Bot. 1985, 55, 395–402. [Google Scholar] [CrossRef]
- Egli, D.B.; Ramseur, E.L.; Zhen-wen, Y.; Sullivan, C.H. Source-Sink Alterations Affect the Number of Cells in Soybean Cotyledons. Crop Sci. 1989, 29, 732. [Google Scholar] [CrossRef]
- Schonbeck, M.W.; Hsu, F.C.; Carlsen, T.M. Effect of Pod Number on Dry Matter and Nitrogen Accumulation and Distribution in Soybean. Crop Sci 1986, 26, 783. [Google Scholar] [CrossRef]
- Board, J.; Harville, B. Late-Planted Soybean Yield Response to Reproductive Source/Sink Stress. Crop Sci. 1998, 38, 763. [Google Scholar] [CrossRef]
- Munier-Jolain, N.G.; Munier-Jolain, N.M.; Roche, R.; Ney, B.; Duthion, C. Seed growth rate in grain legumes I. Effect of photoassimilate availability on seed growth rate. J. Exp. Bot. 1998, 49, 1963–1969. [Google Scholar] [CrossRef]
- Egli, D.B.; Bruening, W.P. Source-sink Relationships, Seed Sucrose Levels and Seed Growth Rates in Soybean. Ann. Bot. 2001, 88, 235–242. [Google Scholar] [CrossRef]
- Liu, X.; Herbert, S.J.; Baath, K.; Hashemi, A.M. Soybean (Glycine max) seed growth characteristics in response to light enrichment and shading. Plant Soil Environ. 2006, 52, 176. [Google Scholar] [CrossRef]
- Proulx, R.; Naeve, S. Pod Removal, Shade, and Defoliation Effects on Soybean Yield, Protein, and Oil. Agron. J. 2009, 101, 971. [Google Scholar] [CrossRef]
- Rotundo, J.L.; Borrás, L.; Westgate, M.E.; Orf, J.H. Relationship between assimilate supply per seed during seed filling and soybean seed composition. Field Crops Res. 2009, 112, 90–96. [Google Scholar] [CrossRef]
- Pereira Flores, M.; Justino, F.; Ruiz Vera, U.; Stordal, F.; Melo, A.; De, R.; Rodrigues, Á. Response of soybean yield components and allocation of dry matter to increased temperature and CO2 concentration. Aust. J. Crop Sci. 2016, 10, 808–818. [Google Scholar]
- Da Silva, A.J.; Magalhães Filho, J.R.; Sales, C.R.G.; de Pires, R.C.M.; Machado, E.C. Source-sink relationships in two soybean cultivars with indeterminate growth under water deficit. Bragantia 2018, 77, 23. [Google Scholar] [CrossRef]
- Fischer, R.A.; Laing, D.R. Yield potential in a dwarf spring wheat and response to crop thinning. J. Agric. Sci. 1976, 87, 113–122. [Google Scholar] [CrossRef]
- Prine, G.M. A Critical Period for Ear Development in Maize. Crop Sci. 1971, 11, 782. [Google Scholar] [CrossRef]
- Chiluwal, A.; Kawashima, T.; Salmeron, M. Soybean seed weight responds to increases in assimilate supply during late seed-fill phase. J. Crop Improv. 2022, 36, 222–238. [Google Scholar] [CrossRef]
- Liu, B.; Liu, X.-B.; Wang, C.; Jin, J.; Herbert, S.J.; Hashemi, M. Responses of soybean yield and yield components to light enrichment and planting density. Int. J. Plant Prod. 2010, 4, 1735–8043. [Google Scholar]
- Swank, J.C.; Egli, D.B.; Pfeiffer, T.W. Seed Growth Characteristics of Soybean Genotypes Differing in Duration of Seed Fill. Crop Sci. 1987, 27, 85. [Google Scholar] [CrossRef]
- Egli, D.B. Seed water relations and the regulation of the duration of seed growth in soybean. J. Exp. Bot. 1990, 41, 243–248. [Google Scholar] [CrossRef]
- Munier-Jolain, N.G.; Ney, B.; Duthion, C. Sequential Development of Flowers and Seeds on the Mainstem of an Indeterminate Soybean. Crop Sci. 1993, 33, 768. [Google Scholar] [CrossRef]
- Poeta, F.; Rotundo, J.; Borrás, L.; Westgate, M. Seed Water Concentration and Accumulation of Protein and Oil in Soybean Seeds. Crop Sci. 2014, 54, 2752. [Google Scholar] [CrossRef]
- Fraser, J.; Egli, D.B.; Leggett, J.E. Pod and Seed Development in Soybean Cultivars with Differences in Seed Size. Agron. J. 1982, 74, 81–85. [Google Scholar] [CrossRef]
- Tekrony, D.M.; Egli, D.B.; Balles, J.; Pfeiffer, T.; Fellows, R.J. Physiological Maturity in Soybean. Agron. J. 1979, 71, 771–775. [Google Scholar] [CrossRef]
- Andrade, F.H.; Ferreiro, M.A. Reproductive growth of maize, sunflower and soybean at different source levels during grain filling. Field Crops Res. 1996, 48, 155–165. [Google Scholar] [CrossRef]
- Rotundo, J.L.; Borrás, L.; Westgate, M.E. Linking assimilate supply and seed developmental processes that determine soybean seed composition. Eur. J. Agron. 2011, 35, 184–191. [Google Scholar] [CrossRef]
- Chiluwal, A.; Haramoto, E.R.; Hildebrand, D.; Naeve, S.; Poffenbarger, H.; Purcell, L.C.; Salmeron, M. Late-Season Nitrogen Applications Increase Soybean Yield and Seed Protein Concentration. Front Plant Sci. 2021, 12, 715940. [Google Scholar] [CrossRef] [PubMed]
- Specht, J.E.; Hume, D.J.; Kumudini, S. V Soybean Yield Potential—A Genetic and Physiological Perspective. Crop Sci. 1999, 39, 1560–1570. [Google Scholar] [CrossRef]
- Mahmoud, A.A.; Natarajan, S.S.; Bennett, J.O.; Mawhinney, T.P.; Wiebold, W.J.; Krishnan, H.B. Effect of Six Decades of Selective Breeding on Soybean Protein Composition and Quality: A Biochemical and Molecular Analysis. J. Agric. Food Chem. 2006, 54, 3916–3922. [Google Scholar] [CrossRef]
- Rowntree, S.C.; Suhre, J.J.; Weidenbenner, N.H.; Wilson, E.W.; Davis, V.M.; Naeve, S.L.; Casteel, S.N.; Diers, B.W.; Esker, P.D.; Specht, J.E.; et al. Genetic Gain × Management Interactions in Soybean: I. Planting Date. Crop Sci. 2013, 53, 1128–1138. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Average Daily Temperature (°C) | Total Precipitation (mm) | |||||
|---|---|---|---|---|---|---|
| * VE to R1 | R1 to R5 | R5 to R7 | VE to R1 | R1 to R5 | R5 to R7 | |
| Experiment 1 | 19.1 | 23.3 | 23.7 | 180.3 | 172.5 | 65.8 |
| Experiment 2 | 21.3 | 22.5 | 21.8 | 221.5 | 288.5 | 58.2 |
| Traits | p-Value | |
|---|---|---|
| Experiment 1 | Experiment 2 | |
| Unit seed weight (mg seed−1) | 0.0031 | <0.0001 |
| Unit pod weight (mg pod−1) | 0.0002 | <0.0001 |
| Seeds per pod | NS * | NS |
| Seed filling duration (days) | <0.0001 | <0.0001 |
| Seed filling rate (mg day−1) | 0.0001085 | 0.04278 |
| Protein concentration (g kg−1) | 0.0165 | NS |
| Oil concentration (g kg−1) | 0.0022 | 0.02003 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poudel, S.; Khatri, D.; Pun Magar, L.; KC, S.; Mukherjee, A.; Lucas, S.; Gebremedhin, M.; Chiluwal, A. Final Seed Size in Soybean Is Determined during Mid-Seed Filling Stage. Agronomy 2024, 14, 763. https://doi.org/10.3390/agronomy14040763
Poudel S, Khatri D, Pun Magar L, KC S, Mukherjee A, Lucas S, Gebremedhin M, Chiluwal A. Final Seed Size in Soybean Is Determined during Mid-Seed Filling Stage. Agronomy. 2024; 14(4):763. https://doi.org/10.3390/agronomy14040763
Chicago/Turabian StylePoudel, Sudip, Deepak Khatri, Lalit Pun Magar, Suraj KC, Atanu Mukherjee, Shawn Lucas, Maheteme Gebremedhin, and Anuj Chiluwal. 2024. "Final Seed Size in Soybean Is Determined during Mid-Seed Filling Stage" Agronomy 14, no. 4: 763. https://doi.org/10.3390/agronomy14040763
APA StylePoudel, S., Khatri, D., Pun Magar, L., KC, S., Mukherjee, A., Lucas, S., Gebremedhin, M., & Chiluwal, A. (2024). Final Seed Size in Soybean Is Determined during Mid-Seed Filling Stage. Agronomy, 14(4), 763. https://doi.org/10.3390/agronomy14040763