Abstract
One biostrategy to boost the sustainability of strawberry cultivation is the application of biostimulants to the growing substrate. Here, we investigated whether the use of biostimulants and their combinations affects the strawberry plants’ phyllochron, phenology, and fruit quality. We tested the absence (control) and presence of biostimulants (arbuscular mycorrhizal fungi (AMF), Ascophyllum nodosum (AN), Trichoderma harzianum (TH), AMF + AN, AMF + TH, AN + TH, and AMF + AN + TH). The experimental design used was in completely randomized blocks (four replications). AMF was represented by a multi-species on-farm inoculant; A. nodosum was represented by the commercial product Acadian®; and T. harzianum was represented by the commercial product Trichodermil®. The leaf emission rate, the occurrence and duration of phenological stages, and fruit quality were assessed. The greatest precocity in terms of harvesting the first fruit was observed in plants grown with AMF + TH, which also had the lowest phyllochron (77.52 °C day·leaf−1). Those treated only with AMF were the latest (144.93 °C day·leaf−1). More flavorful fruits were produced by plants grown with AMF + TH. Plants inoculated with the AMF community, whether or not associated with A. nodosum and T. harzianum, had more than 94% mycorrhizal colonization. We conclude that AMF, A. nodosum, T. harzianum, and their combinations influence the phenology, phyllochron, and fruit quality of strawberry plants. In the growing conditions of the Brazilian subtropics, the AMF and T. harzianum combination shortens the strawberry cycle, from transplanting the daughter plants to harvesting the first fruit, and improves the fruit flavor.
1. Introduction
In the Brazilian subtropics, the lay-flat bag culture system of strawberry (Fragaria × ananassa Duch.) is the most widely adopted [1]. This system in substrate is a response to increased yields and better ergonomics for producers. On the other hand, most producers who adopt this system use chemical inputs on a large scale, and this can lead to environmental contamination [2]. Thus, we need to establish better management practices for agroecosystems, which can be boosted by the use of biostimulants, e.g., arbuscular mycorrhizal fungi (AMF), Ascophyllum nodosum (L.) Le Jolis, and Trichoderma harzianum Rifai.
AMF establish symbiosis with most terrestrial plants. This mutual association is based on the two-way flow between host and fungus [3]. Previously, our research group demonstrated that AMF improve root and shoot growth [4], nutrient accumulation [5], and the synthesis of phytochemicals in leaves, roots [6], and fruit [4] in substrate-grown strawberry plants.
The application of macroalgal extracts in agriculture is related to their biodegradable, non-toxic, and environmentally friendly properties [7]. The beneficial effects of this bioinput include improvements in resistance to stresses in cultivated plants [8,9]. Extracts of the brown macroalgae A. nodosum can promote plant growth, improve protective activity against stresses, and increase nutrient absorption and phytochemical accumulation [10]. A. nodosum extracts improved the morphology of the strawberry root system [11] and increased fruit production [12]. In addition, A. nodosum reduced the progression of Podosphaera aphanis (syn. Sphaerotheca macularis) infection in strawberries [13].
Trichoderma is the main plant biostimulant. After the association between the plant and Trichoderma, this rhizofungus boosts the development of its plant partner [14]. Expression of the bgn13.1 gene of T. harzianum CECT 2413 encoded a β-1,3-glucanase, and this improved strawberry tolerance to Colletotrichum acutatum or Rosellinia necatrix [15]. In addition, this microorganism increased plant growth, improved strawberry yields, and increased the levels of cyanidin 3-O-glucoside, pelargonidin 3-O-rutinoside, and pelargonidin 3-O-acetyl-glucoside in fruit [16]. Anthocyanins are one of the most important health-related compounds found in strawberries. Up to 25 different anthocyanins have been identified [17]. However, only three major anthocyanins are commonly found in high concentrations in this fruit (cyanidin 3-O-glucoside, pelargonidin 3-O-acetyl-glucoside, and pelargonidin 3-O-rutinoside).
These growth-promoting and plant-defense-inducing activities highlight the potential use of these three biostimulants in strawberry plants. Therefore, scientists must intensify studies to improve the understanding of these biotechnologies and support their continued application in commercial strawberry crops. As previously reported, these three biostimulants are applied in isolation in field conditions [4,11,15]. However, as far as we know, there is no literature on the effects of the combined use of AMF, A. nodosum, and T. harzianum on strawberry plants.
The gap regarding the lack of information on the joint application of these bio-tools contributes to limiting their dissemination among producers, who could adopt new sustainable management techniques in strawberry crops through the exploration of microbial multifunctionality, with the potential to increase growth, development, and plant quality. In addition, we do not know the phyllochron, phenology, or fruit quality of this horticultural crop produced with these three biostimulants. Using mathematical models, one way of calculating the number of leaves corresponds to the phyllochron concept (time between the appearance of two successive leaves on the main plant crown) [18]. Phenology studies the morphological changes that occur during plant development.
Here, we investigated whether AMF, A. nodosum, T. harzianum, and their combinations interfere with the phyllochron, phenology, and fruit quality of strawberry plants. Our results will allow growers to use bio-tools aimed at sustainable strawberry cultivation in substrate, linked to decision-making regarding crop management based on phenology, phyllochron, and fruit quality.
2. Materials and Methods
2.1. Plant Material
Bare-root strawberry daughter plants, ‘San Andreas’ cultivar (neutral days), were sourced from the Llahuén nursery, Chile (33°50′15.41″ S; 70°40′03.06″ W). In the Llahuén, daughter plants were produced in fumigated soil, which prevented colonization by AMF and other fungal volunteers during their time in the nursery. The production period of the daughter plants in the Llahuén nursery was seven months, so they received a sufficient number of cold hours, aiming at the accumulation of carbohydrates, photoassimilates, and phytohormones. The daughter plants produced had a 6 to 8 mm diameter crown.
This work was carried out in Passo Fundo (28°15′41″ S; 52°24′45″ W), Rio Grande do Sul (RS), Brazil, from June to September 2023. The plants were grown in a greenhouse covered with low-density polyethylene (150 microns thick).
2.2. Experimental Design
Eight treatments were the absence (control) and presence of biostimulants: arbuscular mycorrhizal fungi (AMF), Ascophyllum nodosum (AN), Trichoderma harzianum (TH), AMF + AN, AMF + TH, AN + TH, and AMF + AN + TH. The experimental design used was in completely randomized blocks (four replications). We planted six plants in every container for each treatment, using four containers per treatment. To represent a replication of the experiment, we collected data from six plants in each container. Therefore, we used six plants per plot (24 plants per treatment; n = 192 plants).
AMF-based biostimulant was represented by a multi-species on-farm inoculant, adapted to strawberry cultivation [19], composed of eight species [20]: Acaulospora koskei Blaszk., Acaulospora rehmii Sieverding and Toro, Claroideoglomus aff. luteum, Claroideoglomus claroideum (N.C. Schenck and G.S. Sm.) C. Walker and A. Schüßler, Claroideoglomus etunicatum (W.N. Becker and Gerd.) C. Walker and A. Schüßler, Funneliformis aff. mosseae, Glomus aff. versiforme, and Glomus sp. (caesaris like). Inoculant was obtained by trap culture [21], with sorghum (Sorghum bicolor (L.) Moench) in sterilized sand (120 °C, 20 min). After five months of multiplication, an AMF-based inoculant was obtained and contained spores, root pieces, and sterile sand. Manually, on-farm inoculant was applied in the planting bed of the strawberry daughter plants.
The A. nodosum-based biostimulant was represented by the commercial product Acadian® (Koppert®, Piracicaba, São Paulo, Brazil), MAPA registration SP 002821-5.000004, made from seaweed harvested from the waters of the North Atlantic Ocean in Nova Scotia, Canada.
The T. harzianum-based biostimulant was represented by the commercial product Trichodermil® (Koppert®, Piracicaba, São Paulo, Brazil), MAPA registration 2007, consisting of T. harzianum (CEPA ESALQ 1306).
We selected Acadian® and Trichodermil® for the diffusion and benefits of both products in other agricultural crops belonging to Rosaceae, such as raspberry (Rubus idaeus L.), apple (Malus pumila Mill.), pear (Pyrus communis L.), and peach (Prunus persica (L.) Batsch).
2.3. Procedures
Daughter plants were transplanted in June 2023 into containers (1 m long × 0.5 m wide) filled with Dallemole® substrate (Vacaria, Rio Grande do Sul, Brazil). The substrate is made up of pine bark, rice husk, rice ash, and class A organic compost. Class A organic compost means that the material has been made from raw plant or animal materials or from agro-industry processing without the addition of sodium (Na+), heavy metals, or potentially toxic synthetic organic elements or compounds. The substrate was pasteurized (vaporized) by the manufacturer. The physical and chemical properties of Dallemole® are shown in Table 1.

Table 1.
Physical and chemical characterization of the Dallemole® substrate.
Treatments inoculated with AMF received 5 g of on-farm inoculant, added monthly to the planting bed of the daughter plants (June) at the time of transplanting and then around the plant crown (July, August, and September). The total amount of on-farm inoculant used, applied from June to September, was 20 g per plant, in accordance with our previous studies [4,5].
For the treatments receiving A. nodosum and T. harzianum monthly (from June to September), a solution of 2.5 mL·L−1 H2O (Acadian®) and 1.5 mL·L−1 H2O (Trichodermil®), respectively, was prepared. These doses were based on the specifications of each product. Subsequently, 10 mL of the Acadian® solution and 10 mL of the Trichodermil® solution were applied to each plant at the crown base in order to run it off into the substrate using a micropipette. The total amount used of Acadian® and Trichodermil®, applied from June to September, was 40 mL per plant for each of the biostimulants.
The irrigation used in the experiment was localized (drip strips) in an automated system (1.40 L·h−1 per dripper). The irrigation regime was activated five times a day, with a total wetting time of 10 min. Nutrient solutions [22] were supplied to the plants weekly, with a 50% reduction in the phosphorus supply [23].
The air temperature inside the greenhouse was monitored using a mini weather station (Figure 1). The absolute minimum and maximum temperatures of 0.10 °C (5 July 2023) and 40.40 °C (29 August 2023) were recorded, respectively (Figure 1). The overall average temperature was 18.24 °C (Figure 1). During the experiment, 26 days with temperatures below 7.0 °C were recorded (Figure 1).
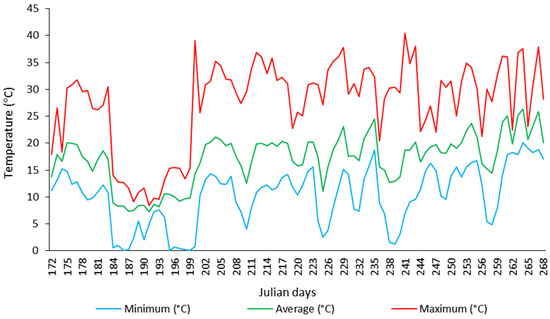
Figure 1.
Temperatures of the greenhouse (from June to September of 2023).
2.4. Mycorrhizal Colonization
Root portions of plants were prepared according to Phillips and Hayman [24]. They were then added to sodium hydroxide solution (NaOH, 10%) and kept in an autoclave (120 °C, 20 min). Afterwards, the roots were submerged in a hydrochloric acid solution (HCl, 1%) for five minutes. Finally, the roots were added to Trypan Blue solution (0.5%) and kept in an autoclave (120 °C, 20 min).
Mycorrhizal colonization (MC, %) was determined according to Trouvelot et al. [25]:
This percentage was evaluated based on the observation of 50 segments, approximately 1 cm (cm) of root, from each treatment. Subsequently, the roots were observed under a microscope to identify the presence of AMF structures.
2.5. Trichoderma–Plant Root Detection
To detect Trichoderma, the methodology proposed by Fernandez [26] was used. To this end, portions of the roots were removed from the substrate and washed in running water. Approximately 30 portions of roots (2 mm in diameter) from each treatment were cut and added to 70% alcohol for 30 s. Then, the roots were subjected to asepsis in hypochlorite + water (1:1 v/v for 2 min, rinsed with sterilized water, and dried on filter paper. Thus, they were transferred to potato dextrose agar culture medium and incubated in a growth chamber (25 °C, 12 h of light) for seven days.
2.6. Phyllochron
Phyllochron was assessed weekly by counting the number of leaves from the start of leaf emission (from the main crown) until the first fruit was harvested. A new leaf was considered to have emerged when it was visible and approximately 1 cm long [27].
The average daily temperature (ADT) was calculated as the arithmetic mean of the temperatures recorded by the mini weather station every hour:
The base temperature (BT), below which there is no leaf emergence, considered for the strawberry crop was 7.0 °C [28]. Daily thermal sum (DTS) was accumulated since the daughter plants were transplanted, resulting in accumulated thermal sum (ATS), that is, ATS (°C·day−1) = Σ DTS.
The DTS was calculated according to Gilmore Junior and Rogers [29] and Arnold [30]:
A regression analysis was carried out between the number of leaves and ATS. The angular coefficient of the linear regression was considered to be the leaf emission rate (leaves °C·day−1), and the phyllochron (°C day·leaf−1) was estimated by the inverse of the angular coefficient of the linear regression [31,32].
2.7. Phenology
Phenological evaluations consisted of weekly observations of five stages of development (Figure 2) according to the phenological scale [33] and the Biologische Bundesanstalt, Bundessortenamt und Chemische Industrie (BBCH) coding. After the first inflorescence, the primary flower was marked with a satin ribbon to make observations and notes on the subsequent stages: first set flowers at the bottom of the rosette (BBCH 55), first flowers open (primary or A-flower) (BBCH 60), receptacle protruding from the sepal whorl (BBCH 71), and main harvest, more colored fruits (BBCH 87).
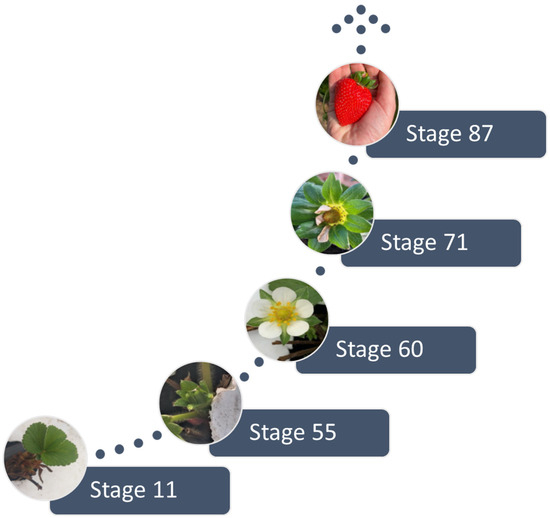
Figure 2.
The strawberry phenological stages [33] considered for evaluation.
The occurrence (O) date of each phenological stage was indicated in days after transplanting (DAT), and its duration (D) was expressed in days.
2.8. Fruit Quality
In September, total soluble solids (TSSs, %) and total titratable acidity (TTA, % of citric acid) were analyzed from 15 fruits from each treatment for each replication [34]. To evaluate the fruit flavor, the TSS/TTA ratio was determined.
2.9. Data Analysis
To estimate phyllochron, a linear regression was performed between the number of leaves and the ATS. These data were submitted to an analysis of variance (ANOVA), and the means of the treatments were compared using the Tukey test (5% probability of error) by Costat® software version 6.45 [35]. Phenology data were presented descriptively. The occurrence date of each phenological stage was defined based on the measure of central tendency referring to the mode [27].
Phenological data were also subjected to multivariate analysis using Genes® software version 1990.2019.91 [36]. Mahalanobis distance (D2) was used to verify the dissimilarity among the biostimulants, and, through this, the relative contribution of the phenological stages to the divergence among the treatments was obtained [37]. The Unweighted Pair Group Method with Arithmetic Mean (UPGMA) was used for the grouping analysis of the biostimulants. The dendrogram was cut using 45% dissimilarity, and the grouping was validated by the cophenetic correlation coefficient (CCC).
Fruit quality data were also submitted to an ANOVA, and the means of the treatments were compared using the Tukey test (5% probability of error) by Costat® software [35].
3. Results
3.1. Mycorrhizal Colonization
Plants inoculated with the AMF community, whether or not associated with A. nodosum and T. harzianum, had more than 94% mycorrhizal colonization (Figure 3). No contamination was observed in plants grown without AMF (Figure 3). The AMF structures observed in the roots were hyphae, vesicles, and arbuscules (Figure 4).
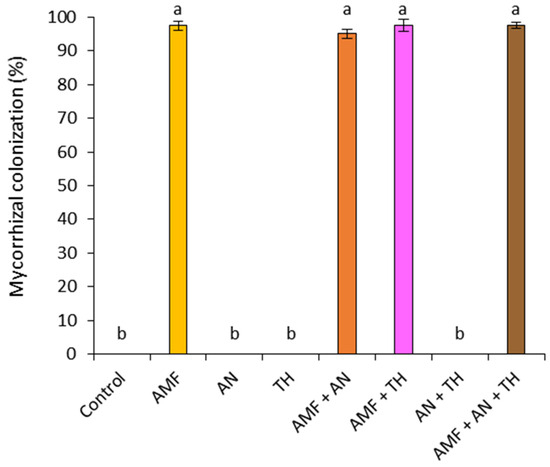
Figure 3.
Mycorrhizal colonization of roots in the absence and presence of biostimulants. Data are presented as mean ± standard deviation. Means followed by the same letter in the column do not differ significantly using the Tukey test (p ≤ 0.05). Control: absence of biostimulants; AMF: arbuscular mycorrhizal fungi; AN: A. nodosum; TH: T. harzianum.
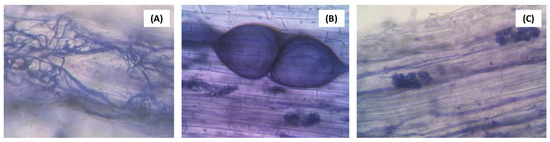
Figure 4.
AMF structures visualized in strawberry roots, ‘San Andreas’ cultivar, refer to hyphae (A), vesicles (B), and arbuscules (C). Optical microscope observation, with a magnification of 400×.
3.2. Trichoderma–Plant Root Detection
Plants cultivated with T. harzianum showed a 67% incidence (I) of this fungus in the roots (Figure 5). Compared to the double combinations (AMF + TH and AN + TH), the association of the three biostimulants (AMF + AN + TH) represented a 40% incidence of T. harzianum in strawberry roots (Figure 5). There was no contamination of T. harzianum in treatments without this rhizofungus (CONT, AMF, AN, and AMF + AN) (Figure 5).
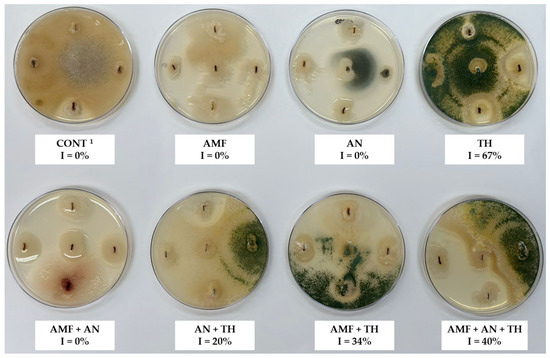
Figure 5.
Incidence of T. harzianum in strawberry roots, ‘San Andreas’ cultivar. 1 CONT: control (absence of biostimulants); AMF: arbuscular mycorrhizal fungi; AN: A. nodosum; TH: T. harzianum.
3.3. Phyllochron
In decreasing order, the leaf emission rate was 0.0129, 0.0121, 0.0115, 0.0113, 0.0101, 0.0097, 0.0092, and 0.0069 accumulated leaves at each °C·day−1 for AMF + TH, AMF + AN, TH, CONT, AN, AN + TH, AMF + AN + TH, and AMF, respectively (Figure 6), with phyllochrones of 77.52, 82.64, 86.96, 88.50, 99.01, 103.09, 108.70, and 144.93 °C day·leaf−1, in the same treatment order, to produce two consecutive leaves (Table 2).
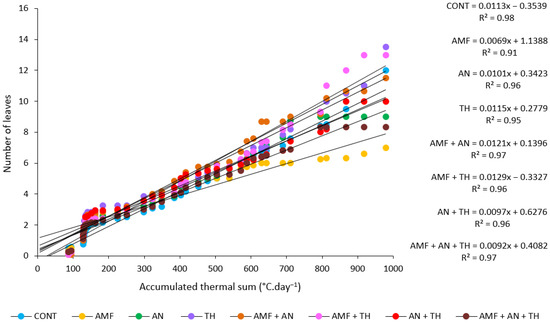
Figure 6.
Estimated phyllochron of strawberry plants, ‘San Andreas’ cultivar, in the absence and presence of biostimulants. CONT: control (absence of biostimulants); AMF: arbuscular mycorrhizal fungi; AN: A. nodosum; TH: T. harzianum.
Table 2.
Phyllochron of strawberry plants, ‘San Andreas’ cultivar, in the absence and presence of biostimulants.
Plants treated with AMF + TH were considered the earliest to start fruit harvest, as they had the lowest phyllochron value (77.52 °C day·leaf−1) (Figure 6 and Table 2), while plants treated with AMF alone were the latest, as they had the highest phyllochron value (144.93 °C day·leaf−1) (Figure 6 and Table 2). This difference between the highest (AMF) and lowest (AMF + TH) phyllochron values was 47% (Table 2). The day interval for a new leaf to emerge ranged from 6.9 (AMF + TH) to 10.9 (AMF).
3.4. Phenology
Plants grown without biostimulants (CONT) had a longer stage 11 duration (33 days), while plants treated with AN, TH, and AMF + TH only remained in this stage for 23 days before starting to flower (stage 55), which occurred 23 days after transplanting the daughter plants (Table 3).
Table 3.
Phenological dates of strawberry plants, ‘San Andreas’ cultivar, in the absence and presence of biostimulants.
The CONT, AMF, AN + TH, and AMF + AN + TH treatments were the latest to start flowering (Table 3). The start of flowering ranged from 35 days (AN, TH, and AMF + TH) to 40 days (CONT, AMF, AN + TH, and AMF + AN + TH) after transplanting the daughter plants (Table 3). AMF + TH showed the shortest cycle from the date of transplanting to the start of flowering (Table 3). This interval ranged from 26 days (AMF + TH) to 37 days (AN) (Table 3).
The inflorescence primary flower opening (stage 60) of the plants cultivated with AN, TH, and AMF + AN occurred in July, and that of the plants treated or not with the other biostimulants occurred in August (Table 3). The combination of the three biostimulants (AMF + AN + TH) increased the stage 60 duration, which represented an average duration 63% greater than the other treatments (Table 3).
The AN, TH, and AMF + AN treatments were the first to start fruiting (stage 71), while AMF + AN + TH was the last to reach this stage (Table 3). The longest duration of stage 71 was 30 days for AN and AMF + AN, while plants grown with AMF + TH only remained in this stage for 14 days (Table 3). This difference between the highest (AN and AMF + AN) and lowest (AMF + TH) stage 71 duration was 53% (Table 3).
The first fruit harvest (stage 87) began in the plants grown with AMF + TH (Table 3), confirming their precocity as indicated by the phyllochron analysis (Figure 6). This same biostimulant combination showed the shortest cycle from the date of transplanting to the harvest start (Table 3). This interval ranged from 61 days (AMF + TH) to 75 days (AMF) and represented a 19% difference between the two treatments (Table 3).
The accumulated heat sum from transplanting to the first fruit harvest for each treatment was 1814.34 °C·day−1 for CONT, 1982.01 °C·day−1 for AMF, 1638.99 °C·day−1 for AN, 1574.59 °C·day−1 for TH, 1658.40 °C·day−1 for AMF + AN, 1650.38 °C·day−1 for AMF + TH, 1945.70 °C·day−1 for AN + TH, and 1940.35 °C·day−1 for AMF + AN + TH (Figure 7).
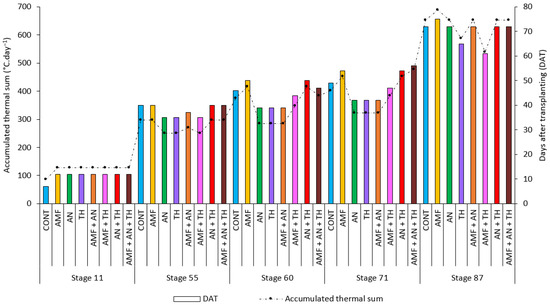
Figure 7.
The accumulated thermal sum required for each treatment to reach the main strawberry phenological stages. CONT: control (absence of biostimulants); AMF: arbuscular mycorrhizal fungi; AN: A. nodosum; TH: T. harzianum.
The earliest treatments to start flowering were AN, TH, and AMF + TH, which required 250.63 °C·day−1 to reach this stage (Figure 7). Plants treated with AMF + TH started fruiting when they accumulated 384.94 °C·day−1, and, after 14 days, this treatment reached 538.43 °C·day−1, at which point the main fruit was harvested and characterized AMF + TH as the earliest treatment (Table 3, Figure 6 and Figure 7). Plants treated only with AMF needed to accumulate 688.70 °C·day−1 to reach harvest and were therefore considered the latest (Table 3, Figure 6 and Figure 7). This difference between the highest (AMF) and lowest (AMF + TH) accumulation of °C·day−1 to start fruit harvest (stage 87) was 22% (Figure 7).
The multivariate analysis indicated dissimilarity among the eight treatments studied. Based on phenology, four groups were generated by the UPGMA method. Heterogeneity was illustrated by a dendrogram (Figure 8), whose fit, calculated by the CCC, was 93%.
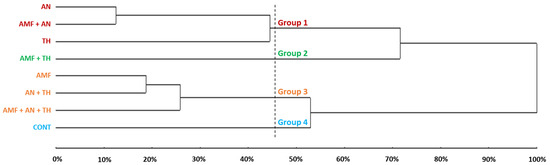
Figure 8.
Grouping by the UPGMA of the treatments in relation to the occurrence (in DAT) of the main strawberry phenological stages. CONT: control (absence of biostimulants); AMF: arbuscular mycorrhizal fungi; AN: A. nodosum; TH: T. harzianum.
Group 1 (red) brought together three treatments (AN, AMF + AN, and TH) (Figure 8). This group performed similarly in terms of stage 11 duration and stages 60 and 71 occurrences (Table 3). Group 2 (green) consisted only of AMF + TH (Figure 8). This treatment stood out in terms of phyllochron (Table 2), the shortest day interval from the start of flowering to the start of harvest, and the shortest time (in days) between transplanting and the first fruit harvest (Table 3). Group 3 (orange) brought together three treatments (AMF, AN + TH, and AMF + AN + TH) (Figure 8). This group was similar in terms of the duration of stages 11 and 55, the occurrence of stages 60 and 71, and the interval of days from transplanting to the start of flowering (Table 3). Finally, group 4 (blue) was formed only by the control (absence of biostimulants) (Figure 8), which had a longer duration of stage 11 (Table 3).
The phenological stage that contributed the most to the divergence among the four groups formed (Figure 8) was the receptacle protruding from the sepal crown (stage 71), which explained 39.51% of the variability among the biostimulants studied (Table 4).
Table 4.
Relative contribution (Sj) of phenological stages to heterogeneity among strawberry plants, ‘San Andreas’ cultivar, in the absence and presence of biostimulants.
3.5. Fruit Quality
There was a significant effect of biostimulants only on the TSS/TTA ratio. All biostimulant treatments and their combinations provided significantly better quality than the control. Plants grown with AMF + TH produced tastier fruits, with a TSS/TTA ratio 48% higher than the fruits produced by plants grown without biostimulants (Table 5).
Table 5.
Fruit quality of strawberry plants, ‘San Andreas’ cultivar, in the absence and presence of biostimulants.
4. Discussion
In this study, we show that the use of biostimulants altered the growth and development of strawberry plants in the ‘San Andreas’ cultivar.
Air temperature plays a significant role in shaping strawberry development, particularly in processes such as leaf emission. The thermal sum (°C·day−1) is a biologically realistic measure for estimating growth and development in relation to air temperature [38]. In our research, the thermal sum that was accumulated from transplanting to the first fruit harvest had a mean value of 1775.59 °C day·leaf−1 for all strawberry treatments in the ‘San Andreas’ cultivar. The thermal sum is more significant for plants than calendar days.
The strawberry phyllochron extends beyond mere cultivar evaluations, encompassing diverse agroecosystems and spanning numerous years of cultivation [39]. This research provides insights into the variability of phyllochron in response to environmental, genetic, and agronomic factors. Studies have explored how different producing regions, with their unique climatic and soil characteristics, can affect the phyllochron in strawberries. In addition, the temporal dimension adds complexity to the study of strawberry phyllochron by considering variations across cultivation years [27]. The biostimulants AMF + TH exhibited a 47% decrease in phyllochron per °C day·leaf−1 in comparison to the biostimulant AMF (Table 2).
Additionally, our research shows that the average value of leaf emission rate for strawberry, ‘San Andreas’ cultivar, is 100.27 ± 29.76 °C day·leaf−1. In another experiment, it was found that the ‘Albion’ cultivar exhibited a leaf emission rate of 149.34 ± 31.3 °C day·leaf−1 [40]. The leaf emission speed has already been studied for various strawberry cultivars [41]. In our study, strawberry plants that were treated with the biostimulant mixture AMF + TH required 63.30% less time to emit a new leaf compared to the plants that were treated with the biostimulant AMF.
The BBCH scale is a standardized system designed for the consistent coding of phenologically analogous growth stages across all plant species, including both mono- and dicotyledonous varieties. This stage serves as a valuable tool in phenological research and agricultural management by providing a uniform language to describe and categorize the developmental phases of plants. The BBCH scale thus stands as an indispensable tool in the realm of plant phenology, promoting consistency and clarity in the characterization of growth stages across a wide spectrum of plant species [42,43].
In the present study, the assessment of BBCH phenological growth stages revealed noteworthy distinctions in the temporal development of strawberry plants. Specifically, the stage denoted by the first unfolding of leaves (BBCH 11) in the control group exhibited a prolonged duration when contrasted with strawberry plants subjected to various biostimulant applications (Table 3). The decrease in days between the control and other treatments was 30.30% in AN, TH, and AMF + TH. This divergence in temporal patterns underscores the potential impact of biostimulants on the developmental trajectory of strawberry plants. The observed discrepancy in the duration of the first leaf unfolding stage suggests that the application of biostimulants may exert an influence on the rate or efficiency of early growth processes in strawberries. Arcidiacono et al. [44] investigated the colonization of tomato roots, ‘Pisanello’ cultivar, by AMF at different growth stages, and they observed the presence of arbuscules and vesicles in roots at stage BBCH 62.
A. nodosum was the source of the first commercialized liquid seaweed extract [45]. The literature indicates that this biostimulant improves fruit ripening, crop yield, and nutritional quality [46,47]. In addition, algal extracts can mitigate abiotic stresses (e.g., salinity) because they affect membrane stability, the synthesis of compatible solutes, and osmoprotective compounds [48]. Despite the large number of genes involved in the complex mechanisms of A. nodosum [49], these benefits are attributed to the bioactive compounds constituents of the extracts, such as phytohormones, polyamines, polysaccharides, amino acids, polyunsaturated fatty acids, and sterols [50,51,52].
Understanding the temporal dynamics of phenological events is critical for discerning the nuanced effects of biostimulants on the development of strawberry plants. Our work showed that the earliest treatment was using AMF + TH biostimulants, needing only 35 days to reach the BBCH 87 stage: main harvests (more fruits colored). In this regard, Bona et al. [53] reported that AMF promoted flowering by increasing phosphorus intake. On the other hand, according to Fraceto et al. [54], the application of T. harzianum can increase the availability of nitrogen for metabolic reactions, resulting in higher levels of amino acids and proteins and faster strawberry plant growth. In addition to the agronomic benefits, the application of beneficial fungi in agriculture, such as AMF and Trichoderma spp., makes it possible to reduce the use of biocides and their negative effects on agroecosystems.
The combined use of AMF and T. harzianum shortened the strawberry cycle, from transplanting daughter plants to harvesting the first fruits. For scientists, this finding opens avenues for new research aimed at enhancing the yield and quality of strawberries through the use of these seven biostimulants. For producers, our findings contribute to the establishment of sustainable management with beneficial fungi as a way to anticipate fruit harvests. In addition, our results on precocity, demonstrated by the phenology and phyllochron of plants cultivated with AMF and T. harzianum, enable the early delivery of fruits to the consumer market when supply is still low and the price paid for the fruits is higher.
Various biotic and abiotic factors influence the multifunctionality of microorganisms in agricultural systems [55]. Cross-talk between microorganisms such as AMF and Trichoderma spp. depends on this. Generally, mutualism between plants and microorganisms represents a reciprocal symbiotic system in which the associated symbionts perform more powerful functions compared to their use in isolation [56]. For example, Wang et al. [57] reported that the combination of AMF and T. harzianum reduced the incidence of soil-borne diseases and enhanced the quality of Salvia miltiorrhiza Bunge, while Martínez-Medina et al. [58] showed that the use of these associated microorganisms improved the nutritional composition of melon seedlings (Cucumis melo L.).
Although AMF and Trichoderma spp. have been used as plant biostimulants for many years, little is known about the molecular changes that occur in plants when these microorganisms are applied together [59]. The association among symbiotic microorganisms can have positive effects by regulating the physiological metabolism of plants [56]. During the first hours of interaction, the process of root colonization by AMF or Trichoderma spp. is mediated by phytohormones, including salicylic acid, jasmonic acid, and ethylene [60,61]. For colonization to take place, AMF [62] and Trichoderma spp. [63] must produce and secrete small molecules recognizable by plant cell receptors, represented by jasmonic acid in the case of AMF [64] and salicylic acid for Trichoderma spp. [61].
The synergy among endophytic fungi can trigger numerous mechanisms related to plant defense against biotic and abiotic stresses [65], which explains the combined treatment (AMF + T. harzianum) superiority in relation to the growth and development of ‘San Andreas’. AMF mechanisms include increased nutrient acquisition and photosynthetic activity, as well as phytohormone profile modulation [66,67,68]. Trichoderma spp. stimulates the production of proteins, auxins, phenylpropanoids, and phytoalexins and acts in the release of volatile and non-volatile molecules that improve the absorption of macronutrients and micronutrients [69,70,71]. Therefore, the better performance in terms of phyllochron, phenology, and fruit quality of plants cultivated with AMF + T. harzianum can be attributed to the synergistic effect of these microorganisms in promoting plant growth. Based on its efficiency, we can recommend the use of this combination to increase the development of strawberry plants in the ‘San Andreas’ cultivar.
The better flavor of the fruits of plants cultivated with AMF + T. harzianum may be the result of the greater supply of nutrients, especially potassium [72], and the action of growth-stimulating substances (phytohormones and phytochemicals) produced by the activity of both fungi in the root zone of plants [73]. For example, the fungal infection observed in the roots (Figure 4 and Figure 5) may have represented a stressful biotic factor for the plant host. As a defense response, plants intensify the production of secondary metabolites [4]. These biomolecules, depending on the environmental conditions to which they are exposed (moderate temperatures around 20 °C, for example), produce sugars through their degradation [74], and this explains the increase in the sugar content of fruits and, consequently, the best flavor of the berries (Table 5).
In our study, the use of biostimulants alone did not have the best effects on the strawberry plant ‘San Andreas’ cultivar. For example, plants treated with AMF alone were the latest to start fruit harvesting (Figure 6 and Table 2). This is because AMF demands carbohydrates from the plants for its maintenance, and thus, the partitioning of photoassimilates occurs among the plant organs and includes AMF [75]. Therefore, mycorrhized plants allocate less energy to leaf emission and show greater phyllochron (Figure 6 and Table 2), which results in an energy imbalance for leaf emission [75]. However, in association with T. harzianum, AMF promoted plant earliness (Figure 6 and Table 2). This suggests that T. harzianum acts as a mitigating agent for AMF demand for carbohydrates, probably due to the complex regulatory network of secondary metabolites, such as phytohormones, and the regulation of biochemical and physiological expression related to plants.
Results about the phyllochron and phenology of the ‘San Andreas’ cultivar help determine the earliness of the plants treated with biostimulants. It should be noted that the combination of AMF and T. harzianum shortened the strawberry cycle, from transplanting the daughter plants to harvesting the first fruit. This finding could have a significant impact from an agronomic point of view since the combined application of these fungi could generate a significant increase in the yield of this horticultural crop.
5. Conclusions
In the growing conditions of southern Brazil, plants grown with AMF + TH are the earliest to start fruit harvest, and those treated only with AMF are the latest. The combination of AMF and T. harzianum shortens the strawberry cycle, from transplanting the daughter plants to the first fruit harvest, and improves the fruit flavor. These findings contribute valuable insights into the dynamics of strawberry plant phenology and quality in response to biostimulant interventions. Our approach provides useful tools and insights into the use of biostimulants to enhance the balance between sugar content and citric acid content in fruits, reduce the number of days from planting to strawberry fruit harvest, and optimize leaf emissions based on temperature.
Author Contributions
All authors contributed equally to the design and writing of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data will be made available upon request.
Acknowledgments
To Koppert (São Paulo, São Paulo, Brazil) and Bioagro Comercial Agropecuária Ltd.a. (Araucária, Paraná, Brazil). Pedro Palencia was supported by the Spanish Ministry of Universities grant European Union NextGenerationEU (BDNS Code: 571952).
Conflicts of Interest
The AMF that were used in this work are regulated by the Sistema Nacional de Gestão do Patrimônio Genético e do Conhecimento Tradicional Associado (SisGen) of the Ministry of the Environment, Brazil, according to the registration number A198F50.
References
- Alves, M.C.; Matoso, E.S.; Peil, R.M.N. What is the profile of strawberry producers in the south Brazilian region and what do they think about substrate cultivation? Hortic. Bras. 2020, 38, 428–433. [Google Scholar] [CrossRef]
- Chiomento, J.L.T.; De Nardi, F.S.; Kujawa, S.C.; Deggerone, Y.S.; Fante, R.; Kaspary, I.J.; Dornelles, A.G.; Huzar-Novakowiski, J.; Trentin, T.S. Multivariate contrasts of seven strawberry cultivars in soilless cultivation and greenhouse in southern Brazil. Adv. Chem. Res. 2023, 2, 62–76. [Google Scholar] [CrossRef]
- Ho-Plágaro, T.; García-Garrido, J.M. Molecular regulation of arbuscular mycorrhizal symbiosis. Int. J. Mol. Sci. 2022, 23, 5960. [Google Scholar] [CrossRef] [PubMed]
- Chiomento, J.L.T.; De Nardi, F.S.; Filippi, D.; Trentin, T.S.; Dornelles, A.G.; Fornari, M.; Nienow, A.A.; Calvete, E.O. Morpho-horticultural performance of strawberry cultivated on substrate with arbuscular mycorrhizal fungi and biochar. Sci. Hortic. 2021, 282, 110053. [Google Scholar] [CrossRef]
- Trentin, T.S.; Dornelles, A.G.; Trentin, N.S.; Huzar-Novakowiski, J.; Calvete, E.O.; Chiomento, J.L.T. Addition of arbuscular mycorrhizal fungi and biochar in the cultivation substrate benefits macronutrient contents in strawberry plants. J. Soil. Sci. Plant Nutr. 2022, 22, 2980–2991. [Google Scholar] [CrossRef]
- Chiomento, J.L.T.; De Nardi, F.S.; Filippi, D.; Trentin, T.S.; Anzolin, A.P.; Bertol, C.D.; Nienow, A.A.; Calvete, E.O. Mycorrhization of strawberry plantlets potentiates the synthesis of phytochemicals during ex vitro acclimatization. Acta Sci. Agron. 2022, 44, 55682. [Google Scholar] [CrossRef]
- Tajdinian, S.; Rahmati-Joneidabad, M.; Ghodoum Parizipour, M.H. Macroalgal treatment to alleviate the strawberry yield loss caused by Macrophomina phaseolina (Tassi) Goid. in greenhouse cultivation system. Front. Sustain. Food Syst. 2022, 6, 1089553. [Google Scholar] [CrossRef]
- El-Sheekh, M.M.; Ahmed, A.Y.; Soliman, A.S.; Abdel-Ghafour, S.E.; Sobhy, H.M. Biological control of soil borne cucumber diseases using green marine macroalgae. Egypt. J. Biol. Pest. Control 2021, 31, 72. [Google Scholar] [CrossRef]
- Mostafa, Y.S.; Alamri, S.A.; Alrumman, S.A.; Hashem, M.; Taher, M.A.; Baka, Z.A. In vitro and in vivo biocontrol of tomato Fusarium wilt by extracts from brown, red, and green macroalgae. Agriculture 2022, 12, 345. [Google Scholar] [CrossRef]
- Patel, J.S.; Selvaraj, V.; More, P.; Bahmani, R.; Borza, T.; Prithiviraj, B. A plant biostimulant from Ascophyllum nodosum potentiates plant growth promotion and stress protection activity of Pseudomonas protegens CHA0. Plants 2023, 12, 1208. [Google Scholar] [CrossRef]
- Alam, M.Z.; Brown, G.; Norrie, J.; Hodges, D.M. Effect of Ascophyllum extract application on plant growth, fruit yield and soil microbial communities of strawberry. Can. J. Plant Sci. 2013, 93, 23–36. [Google Scholar] [CrossRef]
- El-Miniawy, S.M.; Ragab, M.E.; Youssef, S.M.; Metwally, A.A. Influence of foliar spraying of seaweed extract on growth, yield and quality of strawberry plants. J. Appl. Sci. Res. 2014, 10, 88–94. [Google Scholar]
- Bajpai, S.; Shukla, P.S.; Asiedu, S.; Pruski, K.; Prithiviraj, B. A biostimulant preparation of brown seaweed Ascophyllum nodosum suppresses powdery mildew of strawberry. Plant Pathol. J. 2019, 35, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.L.; Hermosa, R.; Lorito, M.; Monte, E. Trichoderma: A multipurpose, plant-beneficial microorganism for eco-sustainable agriculture. Nat. Rev. Microbiol. 2023, 21, 312–326. [Google Scholar] [CrossRef] [PubMed]
- Mercado, J.A.; Barceló, M.; Pliego, C.; Rey, M.; Caballero, J.L.; Muñoz-Blanco, J.; Ruano-Rosa, D.; López-Herrera, C.; Santos, B.; Romero-Muñoz, F.; et al. Expression of the β-1,3-glucanase gene bgn13.1 from Trichoderma harzianum in strawberry increases tolerance to crown rot diseases but interferes with plant growth. Transgenic Res. 2015, 24, 979–989. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, N.; Caira, S.; Troise, A.D.; Scaloni, A.; Vitaglione, P.; Vinale, F.; Marra, R.; Salzano, A.M.; Lorito, M.; Woo, S.L. Trichoderma applications on strawberry plants modulate the physiological processes positively affecting fruit production and quality. Front. Microbiol. 2020, 11, 1364. [Google Scholar] [CrossRef] [PubMed]
- Lopes-da-Silva, F.; Escribano-Bailon, M.T.; Alonso, J.J.P.; Rivas-Gonzalo, J.C.; Santos-Buelga, C. Anthocyanin pigments in strawberry. LWT Food Sci. Technol. 2007, 40, 374–382. [Google Scholar] [CrossRef]
- Xue, Q.; Weiss, A.; Baenziger, P.S. Predicting leaf appearance in field-grown winter wheat: Evaluating linear and non-linear models. Ecol. Modell. 2004, 175, 261–270. [Google Scholar] [CrossRef]
- Chiomento, J.L.T.; Stürmer, S.L.; Carrenho, R.; Costa, R.C.; Scheffer-Basso, S.M.; Antunes, L.E.C.; Nienow, A.A.; Calvete, E.O. Composition of arbuscular mycorrhizal fungi communities signals generalist species in soils cultivated with strawberry. Sci. Hortic. 2019, 253, 286–294. [Google Scholar] [CrossRef]
- Redecker, D.; Schubler, A.; Stockinger, H.; Stürmer, S.L.; Morton, J.B.; Walker, C. An evidence based consensus for the classification of arbuscular mycorrhizal fungi (Glomeromycota). Mycorrhiza 2013, 23, 515–531. [Google Scholar] [CrossRef]
- Stutz, J.C.; Morton, J.B. Successive pot culture reaveal high species richness of arbuscular endomycorrhizal fungi in arid ecosystems. Can. J. Bot. 1996, 74, 1883–1889. [Google Scholar] [CrossRef]
- Furlani, P.R.; Fernandes Júnior, F. Cultivo Hidropônico de Morango em Ambiente Protegido; Embrapa: Curitiba, Brasil, 2004; pp. 102–115. [Google Scholar]
- De Nardi, F.S.; Trentin, T.S.; Trentin, N.S.; Costa, R.C.; Calvete, E.O.; Palencia, P.; Chiomento, J.L.T. Mycorrhizal biotechnology reduce phosphorus in the nutrient solution of strawberry soilless cultivation systems. Agronomy 2024, 14, 355. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Trouvelot, A.; Kouch, J.; Gianinazzi-Pearson, V. Mesure du taux de mycorhization VA d’un système radiculaire: Recherche of method d’estimation ayant une signification fonctionelle. In Aspects Physiologiques et Génétiques des Mycorhizes; Gianinazzi-Pearson, V., Gianinazzi, S., Eds.; Inra Press: Paris, France, 1986; pp. 217–221. [Google Scholar]
- Fernandez, M.R. Manual Para Laboratório de Fitopatologia; Embrapa: Passo Fundo, Brasil, 1993; p. 128. [Google Scholar]
- Deggerone, Y.S.; Trentin, T.S.; Kujawa, S.C.; Albrecht, G.E.; Fante, R.; Kaspary, I.J.; Fornari, M.; Chiomento, J.L.T. Phenology and phyllochron of seven strawberry cultivars grown in substrate and greenhouse in the Brazilian subtropics. Comun. Sci. 2023, 14, 4054. [Google Scholar] [CrossRef]
- Antunes, O.T.; Calvete, E.O.; Rocha, H.C.; Nienow, A.A.; Mariani, F.; Wesp, C.L. Floração, frutificação e maturação de frutos de morangueiro cultivados em ambiente protegido. Hortic. Bras. 2006, 24, 426–430. [Google Scholar] [CrossRef]
- Gilmore Junior, E.C.; Rogers, J.S. Heat units as a method of measuring maturity in corn. Agron. J. 1958, 50, 611–615. [Google Scholar] [CrossRef]
- Arnold, C.Y. Maximum-minimum temperature as a basis for computing heat units. J. Am. Soc. Hortic. Sci. 1960, 76, 682–692. [Google Scholar]
- Klepper, B.; Rickman, R.W.; Peterson, C.M. Quantitative characterization of vegetative development in small cereal grains. Agron. J. 1982, 7, 780–792. [Google Scholar]
- Kirby, E.J.M. Environmental factors influencing the phyllochron. Crop Sci. 1995, 35, 11–19. [Google Scholar] [CrossRef]
- Meier, U.; Graf, H.; Hack, M.; Hess, M.; Kennel, W.; Klose, R.; Mappes, D.; Seipp, D.; Stauss, R.; Streif, J.; et al. Phänologische entwicklungsstadien des kernobstes (Malus domestica Borkh. und Pyrus communis L.), des steinobstes (Prunus-Arten), der johannisbeere (Ribes-Arten) und der erdbeere (Fragaria × ananassa Duch.). Nachrichtenbl. Deut. Pflanzenschutzd. 1994, 46, 141–153. [Google Scholar]
- Zenebon, O.; Pascuet, N.S.; Tiglea, P. Métodos Físico-Químicos para Análise de Alimentos, 4th ed.; Instituto Adolfo Lutz: São Paulo, Brasil, 2008; p. 1020. [Google Scholar]
- CoHort Software. CoStat: Graphics and Statistics Software for Scientists and Engineers, CoHort Software: California, CA, USA, 2003.
- Cruz, C.D. Genes Software—Extended and Integrated with the R, Matlab and Selegen. Acta Sci. Agron. 2016, 38, 547–552. [Google Scholar] [CrossRef]
- Singh, D. The relative importance of characters affecting genetic divergence. Indian J. Genet. Plant Breed. 1981, 41, 237–245. [Google Scholar]
- Lucas, D.D.P.; Streck, N.A.; Bortoluzzi, M.P.; Trentin, R.; Maldaner, I.C. Temperatura base para emissão de nós e plastocrono de plantas de melancia. Rev. Ciênc. Agron. 2012, 43, 288–292. [Google Scholar] [CrossRef][Green Version]
- Schoffel, A.; Koefender, J.; Camera, J.; Golle, D.; Broch, J.; Mugnol, J.; Manfio, C. Transplanting seasons, phyllochron and production of strawberry cultivars in soil. Contrib. Las Cienc. Soc. 2023, 16, 8358–8374. [Google Scholar]
- Mendonça, H.F.C.; Calvete, E.O.; Nienow, A.A.; Costa, R.C.; Zerbielli, L.; Bonafé, M. Estimativa do filocrono de morangueiro em sistemas consorciado e solteiro em ambientes protegidos. Rev. Bras. Frutic. 2012, 4, 15–23. [Google Scholar] [CrossRef]
- Costa, R.C.; Calvete, E.O.; De Nardi, F.S.; Pedersen, A.C.; Chiomento, J.L.T.; Trentin, N.S. Quality of strawberry seedlings can determine precocity. Aust. J. Crop Sci. 2018, 2, 81–86. [Google Scholar] [CrossRef]
- Meier, U. Growth Stages of Mono- and Dicotyledonous Plants; Federal Biological Research Centre for Agriculture and Forestry: Berlin, Germany, 2001. [Google Scholar]
- Meier, U. Strawberry: Growth Stages of Plants; Blackwell Wissenschafts-Verlag: Berlin, Germany, 1997; pp. 62–64. [Google Scholar]
- Arcidiacono, M.; Pellegrino, E.; Nuti, M.; Ercoli, L. Field inoculation by arbuscular mycorrhizal fungi with contrasting life-history strategies differently affects tomato nutrient uptake and residue decomposition dynamics. Plant Soil. 2023, 1–23. [Google Scholar] [CrossRef]
- Stirk, W.A.; Rengasamy, K.R.R.; Kulkarni, M.G.; van Staden, J. Plant biostimulants from seaweed. In The Chemical Biology of Plant Biostimulants; Geelen, D., Xu, L., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2020; pp. 31–55. [Google Scholar]
- Khan, W.; Menon, U.; Subramanian, S.; Jithesh, M.; Rayorath, P.; Hodges, D.; Crithcley, A.; Craigie, J.; Norrie, J.; Prithiviraj, B. Seaweed extracts as biostimulants of plant growth and development. J. Plant Growth. Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Espinosa-Antón, A.; Hernández-Herrera, R.; González, M. Extractos bioactivos de algas marinas como bioestimulantes del crecimiento y la protección de las plantas. Biot. Veg. 2020, 20, 257–282. [Google Scholar]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef]
- De Saeger, J.; Van Praet, S.; Vereecke, D.; Park, J.; Jacques, S.; Han, T.; Depuydt, S. Toward the molecular understanding of the action mechanism of Ascophyllum nodosum extracts on plants. J. Appl. Phycol. 2020, 32, 573–597. [Google Scholar] [CrossRef]
- Vera, J.; Castro, J.; González, A.; Moenne, A. Seaweed polysaccharides and derived oligosaccharides stimulate defense responses and protection against pathogens in plants. Mar. Drugs 2011, 9, 2514–2525. [Google Scholar] [CrossRef] [PubMed]
- Craigie, J.S. Seaweed extract stimuli in plant science and agriculture. J. Appl. Phycol. 2011, 23, 371–393. [Google Scholar] [CrossRef]
- Yalçin, S.; Okudan, E.S.; Karakaș, O.; Önem, A.N.; Bașkan, K.S. Identification and quantification of some phytohormones in seaweeds using UPLC-MS/MS. J. Liq. Chromatogr. Relat. Technol. 2019, 42, 475–484. [Google Scholar] [CrossRef]
- Bona, E.; Lingua, G.; Manassero, P.; Cantamessa, S.; Marsano, F.; Todeschini, V.; Copetta, A.; D’Agostino, G.; Massa, N.; Avidano, L.; et al. AM fungi and PGP pseudomonads increase flowering, fruit production, and vitamin content in strawberry grown at low nitrogen and phosphorus levels. Mycorrhiza 2015, 25, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Fraceto, L.F.; Maruyama, C.R.; Guilger, M.; Mishra, S.; Keswani, C.; Singh, H.B.; Lima, R. Trichoderma harzianum-based novel formulations: Potential applications for management of Next-Gen agricultural challenges. J. Chem. Technol. Biotechnol. 2018, 93, 2056–2063. [Google Scholar] [CrossRef]
- Cano, M.A. A review of interaction of beneficial microorganisms in plants: Mycorrhizae, Trichoderma spp. and Pseudomonas spp. Rev. UDC Actual. Divulg. Cient. 2011, 14, 15–31. [Google Scholar] [CrossRef]
- Cai, X.; Zhao, H.; Liang, C.; Li, M.; Liu, R. Effects and mechanisms of symbiotic microbial combination agents to control tomato Fusarium crown and root rot disease. Front. Microbiol. 2021, 12, 629793. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, M.; Yang, G.; Li, X.; Li, P.; Chen, M. Effect of Glomus versiforme and Trichodema harzianum on growth and quality of Salvia miltiorrhiza. Zhong Yao Cai 2014, 39, 1574–1578. [Google Scholar]
- Martínez-Medina, A.; Pascual, J.A.; Lloret, E.; Roldán, A. Interactions between arbuscular mycorrhizal fungi and Trichoderma harzianum and their effects on Fusarium wilt in melon plants grown in seedling nurseries. J. Sci. Food Agric. 2009, 89, 1843–1850. [Google Scholar] [CrossRef]
- Poveda, J.; Hermosa, R.; Monte, E.; Nicolás, C. Trichoderma harzianum favours the access of arbuscular mycorrhizal fungi to non-host Brassicaceae roots and increases plant productivity. Sci. Rep. 2019, 9, 11650. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; Roldán, A.; Pascual, J.A. Interaction between arbuscular mycorrhizal fungi and Trichoderma harzianum under conventional and low input fertilization field condition in melon crops: Growth response and Fusarium wilt biocontrol. Appl. Soil Ecol. 2011, 47, 98–105. [Google Scholar] [CrossRef]
- Morán-Diez, E.; Rubio, B.; Domínguez, S.; Hermosa, R.; Monte, E.; Nicolás, C. Transcriptomic response of Arabidopsis thaliana after 24 h incubation with the biocontrol fungus Trichoderma harzianum. J. Plant Physiol. 2012, 169, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Maillet, F.; Poinsot, V.; André, O.; Puech-Pagès, V.; Haouy, A.; Gueunier, M.; Cromer, L.; Giraudet, D.; Formey, D.; Niebel, A.; et al. Fungal lipochitooligosaccharide symbiotic signals in arbuscular mycorrhiza. Nature 2011, 469, 58–63. [Google Scholar] [CrossRef]
- Mendoza-Mendoza, A.; Zaid, R.; Lawry, R.; Hermosa, R.; Monte, E.; Horwitz, B.A.; Mukherjee, P.K. Molecular dialogues between Trichoderma and roots: Role of the fungal secretome. Fungal Biol. Rev. 2018, 32, 62–85. [Google Scholar] [CrossRef]
- Hause, B.; Maier, W.; Miersch, O.; Kramell, R.; Strack, D. Induction of jasmonate biosynthesis in arbuscular mycorrhizal barley roots. Plant Physiol. 2002, 130, 1213–1220. [Google Scholar] [CrossRef]
- El-Sharkawy, H.H.A.; Rashad, Y.M.; Ibrahim, S.A. Biocontrol of stem rust disease of wheat using arbuscular mycorrhizal fungi and Trichoderma spp. Physiol. Mol. Plant Pathol. 2018, 103, 84–91. [Google Scholar] [CrossRef]
- Rouphael, Y.; Franken, P.; Schneider, C.; Schwarz, D.; Giovannetti, M.; Agnolucci, M.; De Pascale, S.; Bonini, P.; Colla, G. Arbuscular mycorrhizal fungi act as biostimulants in horticultural crops. Sci. Hortic. 2015, 196, 91–108. [Google Scholar] [CrossRef]
- López-Bucio, J.; Pelagio-Flores, R.; Herrera-Estrella, A. Trichoderma as biostimulant: Exploiting the multilevel properties of a plant beneficial fungus. Sci. Hortic. 2015, 196, 109–123. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Synergistic Biostimulatory action: Designing the next generation of plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef]
- Formisano, L.; Miras-Moreno, B.; Ciriello, M.; El-Nakhel, C.; Corrado, G.; Lucini, L.; Colla, G.; Rouphael, Y. Trichoderma and phosphite elicited distinctive secondary metabolite signatures in zucchini squash plants. Agronomy 2021, 11, 1205. [Google Scholar] [CrossRef]
- Bonini, P.; Rouphael, Y.; Miras-Moreno, B.; Lee, B.; Cardarelli, M.; Erice, G.; Cirino, V.; Lucini, L.; Colla, G. A Microbial-based biostimulant enhances sweet pepper performance by metabolic reprogramming of phytohormone profile and secondary metabolism. Front. Plant Sci. 2020, 11, 567388. [Google Scholar] [CrossRef] [PubMed]
- Nanjundappa, A.; Bagyaraj, D.J.; Saxena, A.K.; Kumar, M.; Chakdar, H. Interaction between arbuscular mycorrhizal fungi and Bacillus spp. in soil enhancing growth of crop plants. Fungal Biol. Biotechnol. 2019, 6, 23. [Google Scholar] [CrossRef]
- Thejaswini, H.P.; Shivakumar, B.S.; Sarvajna, B.S.; Ganapathi, M.; Yallesh, H.S.K. Studies on split application of NPK fertilizers and liquid bio-formulation (Jeevamrutha) on yield and quality of pomegranate (Punica granatum L.) in central dry zone of Karnataka. Pharm. Innov. 2022, 11, 494–498. [Google Scholar]
- Salem, A.; Khandaker, M.M.; Mahmud, K.; Alsufyani, S.J.; Majrashi, A.A.; Rashid, Z.M.; Alenazi, M.M.; Osman, N.; Badaluddin, N.A. Enhancing photosynthesis and root development for better fruit quality, aroma, and lessening of radioactive materials in key lime (Citrus aurantifolia) using Trichoderma harzianum and Bacillus thuringiensis. Plant Physiol. Biochem. 2024, 206, 108295. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Jalaluddin, M.; Garner, J.O.; Yoshimoto, M.; Yamakawa, O. Artificial shading and temperature influence on anthocyanin compositions in sweetpotato leaves. HortScience 2005, 40, 176–180. [Google Scholar] [CrossRef]
- Chiomento, J.L.T.; Filippi, D.; Zanin, E.; Piuco, M.G.; Trentin, T.S.; Dornelles, A.G.; Fornari, M. Arbuscular mycorrhiza potentiates the quality of fruits but does not influence the precocity of goldenberry plants. Braz. J. Dev. 2020, 6, 79041–79056. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).