Abstract
Solanum torvum, a wild relative of eggplant in the Solanaceae family, is widely used as a rootstock to prevent various soil-borne diseases. Despite this valued trait, S. torvum seeds exhibit strong seed dormancy, resulting in low-frequency germination and uneven emergence rates during agricultural production. In this study, exogenous gibberellin (GA) was applied to induce the germination of seeds, thereby enabling the application of transcriptome analysis to explore the effects of exogenous GA on germination. A total of 9723 genes were identified to be involved in GA-induced germination of S. torvum seeds. These GA-associated genes were enriched in amino sugar and nucleotide sugar metabolism, glycolysis, the citrate cycle, pyruvate metabolism, hormone biosynthesis, and signaling transduction. Further analysis of these genes reveals that exogenous GA impaired the endogenous biosynthesis of GA. Analysis of the 285 key genes influenced by GA, auxin, abscisic acid (ABA), and other hormones suggests mutual crosstalk in the signaling of various hormones, enabling the joint regulation of the germination of S. torvum seeds. Additionally, a total of 440 genes encoding transcription factors were identified to be responsible for transcriptional reprogramming during GA-induced seed germination, and putative candidate genes responsible for S. torvum seed dormancy and germination, including ABI5, DOG1, DRM1, LEC1, and PIF, were significantly down-regulated in germinated S. torvum seeds. These findings provide the first insights into transcriptome regulation during GA-induced germination of S. torvum seeds.
1. Introduction
Eggplant (Solanum melongena L.) is a popular vegetable crop because of its preferred flavor and high nutrition. According to the data provided by the World Food and Agriculture Organization (FAO) in 2023, eggplant is one of the most cultivated vegetable crops, attaining global production of 59 million tonnes [1]. Eggplants are susceptible to several soil-borne diseases, including bacterial wilt, fusarium wilt, and verticillium wilt [2]. Along with the continuous expansion of the eggplant cultivation area, soil-borne diseases have become more frequent due to repeated cropping as well as increased temperature and high humidity [3]. Once the bacteria or fungal pathogens infect the host roots and invade the vascular system, the host plants exhibit wilt-symptomatic vessels [4], which eventually causes plant death and, in turn, severe production loss worldwide.
To prevent vascular wilt diseases, eggplants are usually grafted onto resistant rootstocks [5,6,7]. The commonly used rootstock is Solanum torvum Swartz., a wild relative of eggplant in the Solanaceae family. S. torvum is a native shrub that originates from tropical regions of Africa and Asia and is widely found in scrub, roadside, wasteland, or damp places. The species shows high resistance to bacterial and fungal wilt [8] and various environmental stresses, such as salt and cadmium stresses [9,10]. It is, therefore, widely employed in agricultural production as a rootstock for grafting other solanaceous plants, such as tomato, eggplant, and pepper, to improve plant growth, yield, and resistance to diseases and pests [11,12,13].
Due to the strong dormancy, the germination of S. torvum seeds is poor and usually requires considerable time [14], which causes uneven emergence during S. torvum rootstock production. Several methods, such as the application of low temperature, soaking treatment in K3PO4, KNO3, and NaNO3, and the application of exogenous gibberellin (GA), have been applied to break seed dormancy and promote S. torvum seed germination [14,15,16]. Among these methods, the application of GA is the most effective treatment, which breaks seed dormancy and significantly improves the seed germination rate [14,15,16].
GA is a key phytohormone that regulates seed dormancy and germination. It stimulates seed germination by breaking seed dormancy and promotes embryo–seedling transition by turning over seed inhibitors in the seed and activating the synthesis of enzymes that are necessary for germination [17,18,19]. Consistent with the view that elevated GA levels are responsible for dormancy loss in wheat and barley [20,21,22], Arabidopsis and tomato seeds carrying mutations in GA biosynthetic enzymes exhibited a defect in germination [23,24,25]. For example, Peng and Harberd [26] found that tomato seeds lacking GA (gib-1) could not germinate without exogenous GA. These results suggest that the level of GA plays a crucial role in breaking seed dormancy and enabling seed germination. Thus, exogenous application of GA has been widely applied to break seed dormancy and facilitate seed germination.
Although the impact of GA on seed germination has been widely studied in many plant species [27,28,29,30], no study has been undertaken to investigate the underlying mechanism by which exogenous GA stimulates S. torvum germination. In this study, transcriptome analysis was performed to investigate the regulation of gene expression during GA-induced germination of S. torvum seeds. Such effort identified potential genes that are induced by GA and thereby regulate plant energy metabolism and hormone signaling transduction to promote the seed germination process.
2. Materials and Methods
2.1. Plant Materials and Seed Germination
Seeds from S. torvum were obtained from Will Seed Company (Jinan, Shandong, China) and used for a germination test. The seeds were first incubated in 1.5 mM bioactive gibberellin GA3 or distilled water for 24 h at room temperature in the dark. After rinsing 5 times with distilled water, the seeds were placed on two layers of filter paper in Petri dishes and incubated in a growth chamber at 28 °C in darkness. During the germination test, the filter paper was kept moist at all times. Seed germination was monitored for 14 days. When the radicle emerged from the seed coat and extended more than 0.5 cm, the seed was considered to be germinated [31,32]. Germination rates and standard deviations were calculated based on five independent experiments. The germination percentage and the germination index were calculated with the following equations [33]:
2.2. Seed Samples for Transcriptomic Analysis
To ensure all the germinated (G) seeds used for transcriptome analysis were uniform, the seeds were collected immediately once their radicles extended to 0.5 cm. After 12 days, the seeds from the GA-treated group without the emergence of the radicles were collected as ungerminated (NG) seeds. No seeds treated with distilled water for 12 days were observed to be germinated, and were thereby used as a control group (W). Three biological replicates were performed for each treatment.
2.3. Transcriptomic Analysis
Total RNA was extracted from each sample using TRIzol reagent (Invitrogen, Carlsbad, CA, USA), and RNA integrity was analyzed using an Agilent 2100 bioanalyzer. The mRNA obtained by Oligo(dT) magnetic beads was used for library construction using the NEBNext® Ultra™ II Directional RNA Library Prep Kit for Illumina (NEB, Ipswich, MA, USA) according to the manufacturer’s protocol. Initial quantification of the constructed library was performed using Qubit 2.0. After passing the library inspection, the libraries were sequenced on Illumina Novaseq X. After removing low-quality reads, the remaining high-quality reads were mapped against the recently sequenced genome of S. torvum [34].
2.4. Identification of DEGs
Raw data of Fragments Per Kilobase of transcript per Million mapped reads (FPKM) of all recognized genes are presented in Supplemental Table S3. Based on the expression level of FPKM, genes with a log2 fold change > 1 and a p value < 0.01 were defined as differentially expressed genes (DEGs) (Supplemental Table S6). ClusterProfiler software (Version: 3.18.0) was used to analyze the enrichment of Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) terms. GO and KEGG terms with p values < 0.05 were considered to be significantly enriched. Significant differences (p value) were calculated by Student’s t-test.
3. Results
3.1. Effect of GA Treatment on Seed Germination of S. torvum
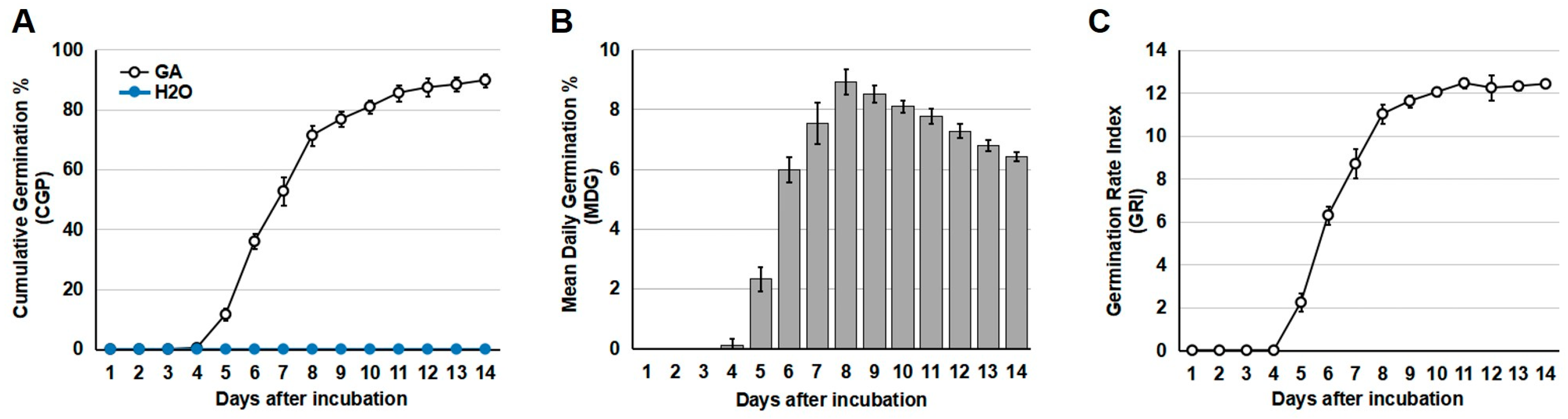
In our preliminary experiment (Supplemental Figure S1), no germination of S. torvum seeds was observed when treated at low temperature (4 °C) or when pre-soaked in KNO3 (20 g/L), MgSO4 (20 g/L), or CaCl2 (20 g/L). Exogenous application of GA treatment, however, stimulated the germination of S. torvum seeds (Supplemental Figure S1). Among the various concentrations of GA, 1.5 mM was the minimum concentration to effectively promote S. torvum seed germination (Supplemental Figure S1). Thus, treatment with 1.5 mM GA was used for the subsequent analysis. After 24 h pre-treatment of 1.5 mM GA, around 10% of GA-treated S. torvum seeds germinated at day 5. Eight days after treatment, the germination rate reached 70%. Although slow germination was still observed after day 8, the peak of germination (around 85%) and the highest Germination Rate Index were observed at day 11 (Figure 1A). By contrast, no germination of S. torvum seeds was observed under pre-treatment with H2O (Figure 1A). Based on these results, pre-soaking treatment with 1.5 mM GA was used to induce the germination of S. torvum seeds, and H2O treatment was used as a negative control.
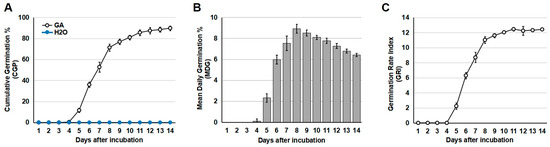
Figure 1.
Germination of S. torvum seeds treated by exogenous application of 1.5 mM GA. (A) Cumulative germination percentage (CGP) of S. torvum seeds. (B) Mean daily germination rate (MDG) of S. torvum seeds. (C) Germination Rate Index (GRI) of S. torvum seeds. X axis, days after incubation with 1.5 mM GA. The data were calculated based on five independent experiments.
3.2. Identification of Differentially Expressed Genes (DEGs) between Germinated and Ungerminated S. torvum Seeds
To investigate the germination mechanism of S. torvum seeds, RNA-seq analyses on a total of nine samples from germinated and ungerminated seeds were performed (Supplemental Figure S2). Three RNA samples, G1, G2, and G3, were extracted from the germinated seeds after GA treatment, while three RNA samples of GA-treated ungerminated seeds were labeled as NG1, NG2, and NG3. The remaining three samples were prepared from H2O treatment (W) as control.
A total of 30,543 genes, including 4842 novel genes, were detected in the nine samples (Supplemental Tables S2–S4). Among them, 26,526 and 25,944 genes were detected in W and NG samples, respectively, while 28,119 genes were detected in G samples. This result suggests that the transcriptome was more active in germinated seeds when compared to water-treated and GA-treated ungerminated seeds. Principal component analysis (PCA) and correlation analysis (Supplemental Figure S3, Table S5) revealed that after GA pre-treatment, the transcriptome of NG seeds was highly related to that in group W, while the transcriptome in group G showed a large variation from groups W and NG.
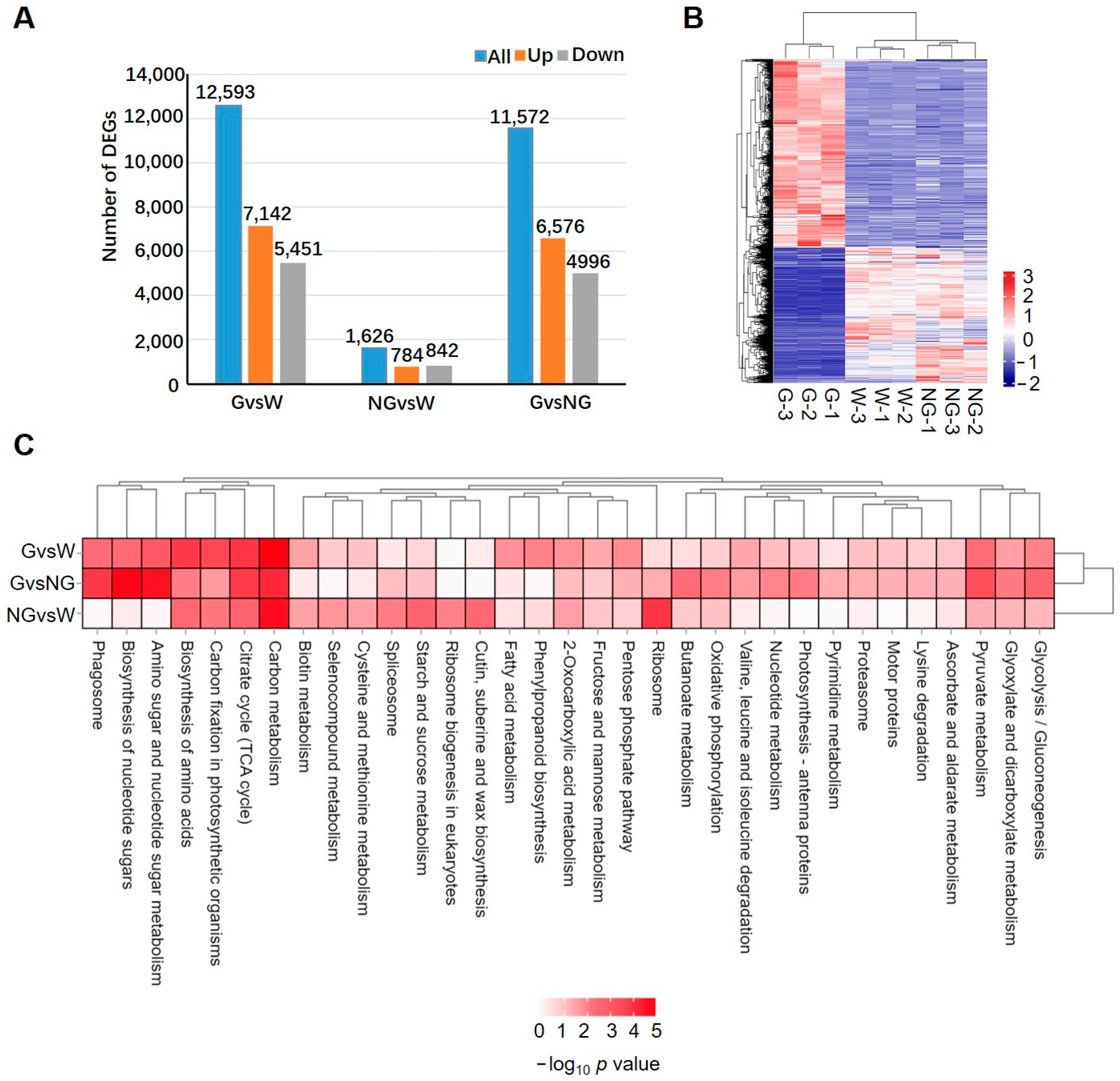
Compared to the H2O-treated control, 12,593 and 1626 genes were differentially expressed genes in the germinated (G vs. W) and ungerminated seeds (NG vs. W), respectively (Figure 2A, Supplemental Table S6). The number of up-regulated genes in the G group was 7142, which was more than the down-regulated genes (5451). The numbers of up- and down-regulated genes in the NG group were 784 and 842, respectively. In the comparison between groups G and NG (G vs. NG), 11,572 DEGs, including 6576 up- and 4996 down-regulated genes, were detected. A heatmap cluster analysis (Figure 2B) further revealed that most of the DEGs showed similar expression patterns in W and NG when compared to the G group. All these results suggested that the transcriptome was significantly reprogrammed in germinated seeds, while only a small number of genes in S. torvum seeds were induced in GA-treated ungerminated seeds.
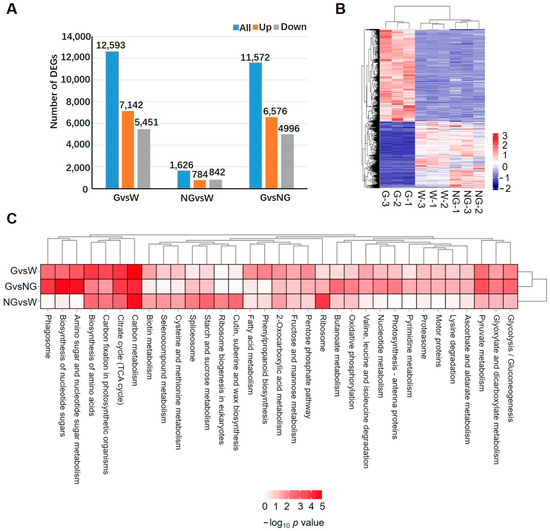
Figure 2.
Analysis of differential expressed genes (DEGs) in germinated seeds in comparison to ungerminated seeds. (A) Number of DEGs identified among germinated (G) and ungerminated seeds (NG) treated by GA compared to control seeds treated by ddH2O. Blue, orange and gray bars represent the number of total, up-regulated, and down-regulated DEGs. (B) Heatmap analysis to show expression pattern of DEGs in each sample. (C) KEGG pathways enriched in DEGs identified in each comparison group.
KEGG enrichment analysis of DEGs (Figure 2C) revealed that pathways of carbon metabolism, the citrate cycle (TCA cycle), carbon fixation in photosynthetic organisms, and the biosynthesis of amino acids were significantly enriched in DEGs detected in both NG- and G-group seeds, suggesting that GA pre-treatment induced a change in energy metabolism and amino acid biosynthesis. No significant difference in pathway enrichment was observed between the G vs. W and G vs. NG comparisons. The enriched pathways included amino sugar and nucleotide sugar metabolism, the biosynthesis of nucleotide sugars, phagosome, glycolysis/gluconeogenesis, glycoxylate metabolism, and dicarboxylate metabolism and pyruvate metabolism. However, DEGs in the NG vs. W group were highly enriched in ribosome-related synthesis, which was different from that in G vs. W and G vs. NG comparisons. Further analysis of up- and down-regulated DEGs (Supplemental Figure S4) revealed that while up-regulated pathways were similar to that detected in all DEGs, only a few of pathways, such as basal transcription factors and spliceosome, were significantly down-regulated in the G vs. W and G vs. NG groups.
GO enrichment analyses (Supplemental Figures S5–S7) were further performed on each group. DEGs in the G vs. W and G vs. NG groups were enriched in similar GO terms for carbohydrate metabolism, the endomembrane system, and pyrophosphatase activity (Supplemental Figures S5 and S6). With regard to up-regulated GO terms, carbohydrate metabolism-related processes, cytoplasmic part, and structural molecular activity were prominent. On the other hand, GO terms related to membrane-bound organelle and RNA metabolic pathways were significantly down-regulated. In the NG vs. W group (Supplemental Figure S7), however, GO terms of non-membrane-bound organelle and small molecular metabolic processes were highly up-regulated, while DNA repair-related GO terms were detected in down-regulated DEGs.
3.3. Identification of Germination-Related Genes (GRGs)
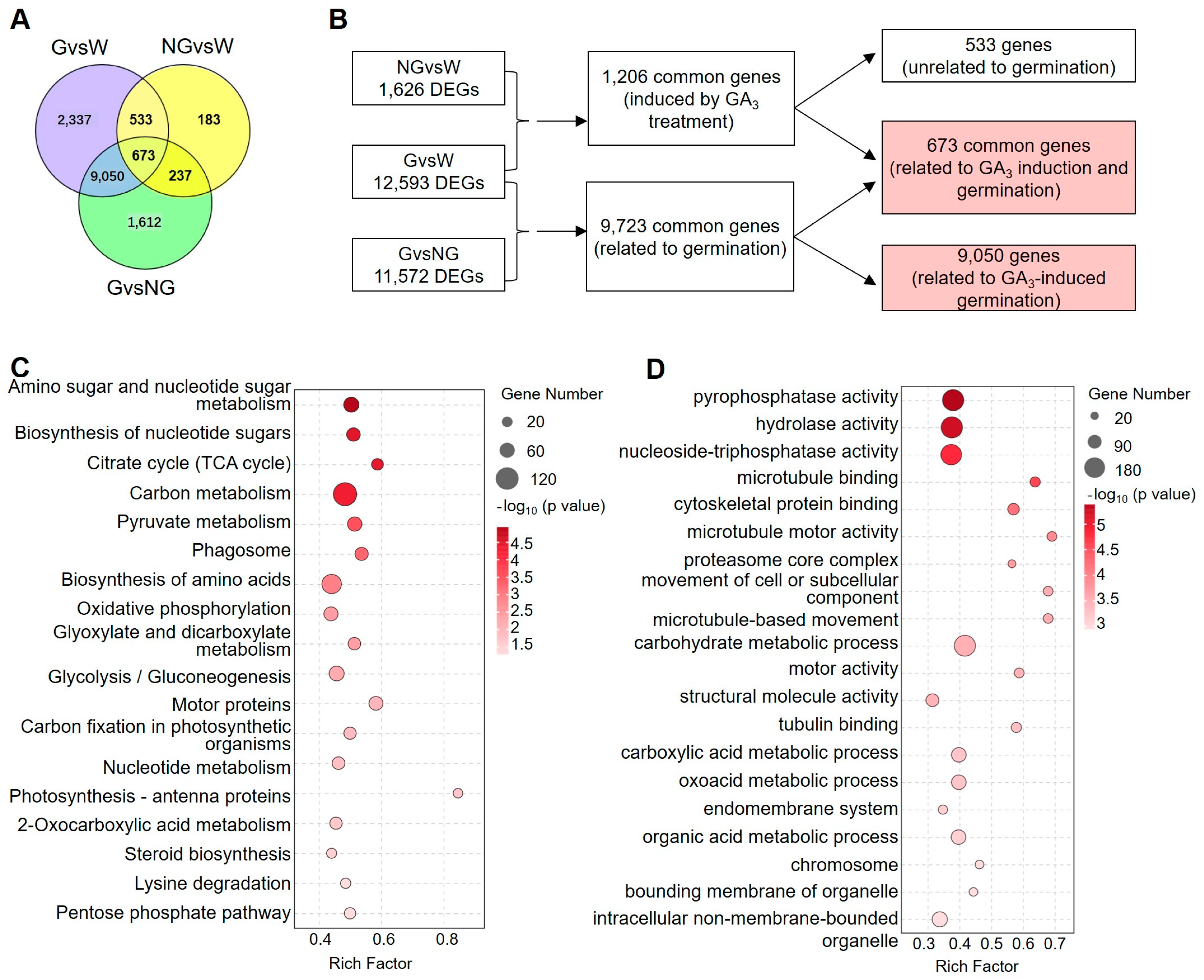
Venn diagrams revealed that only 1206 common genes were detected between the NG vs. W and G vs. W groups (Figure 3A,B). Those genes may be directly induced by GA pre-treatment. On the other hand, a total of 9723 DEGs (Supplemental Table S7), including 673 commonly regulated genes in seeds pre-treated by GA and 9050 specifically regulated genes in germinated seeds, were identified to be germination-related genes (GRGs).
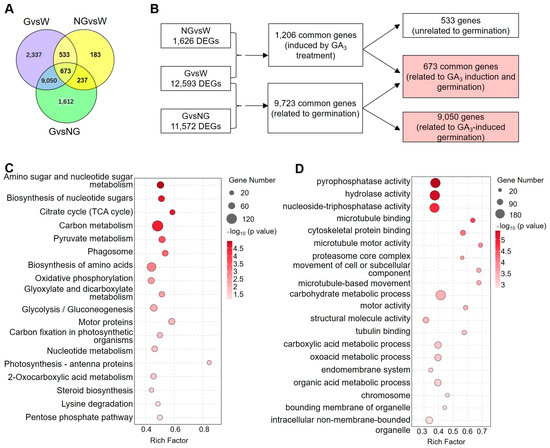
Figure 3.
Analysis of germination-related genes (GRGs). (A) Venn diagram analysis showing the overlap of DEGs among different comparison groups. (B) Flow chart to identify 9723 germination-related genes (GRGs). (C,D) KEGG pathway (C) and GO term (D) enrichment analyses of GRGs.
KEGG and GO enrichment analyses (Figure 3C,D) were performed on those GRGs. The pathways of amino sugar and nucleotide sugar metabolism, the biosynthesis of nucleotide sugars, the citrate cycle (TCA cycle), carbon metabolism, and pyruvate metabolism were profoundly enriched (Figure 3C), suggesting that genes related to energy metabolism were significantly regulated during germination. On the other hand, GO terms related to pyrophosphatase activity, hydrolase activity, and nucleoside-triphosphatase activity were significantly prominent, which supported the KEGG enrichment of energy metabolism. In addition, GRGs related to photosynthesis and the cytoskeleton were also over-represented in KEGG and GO enrichment analyses.
3.4. GRGs Related to Energy Metabolism in S. torvum Seed Germination
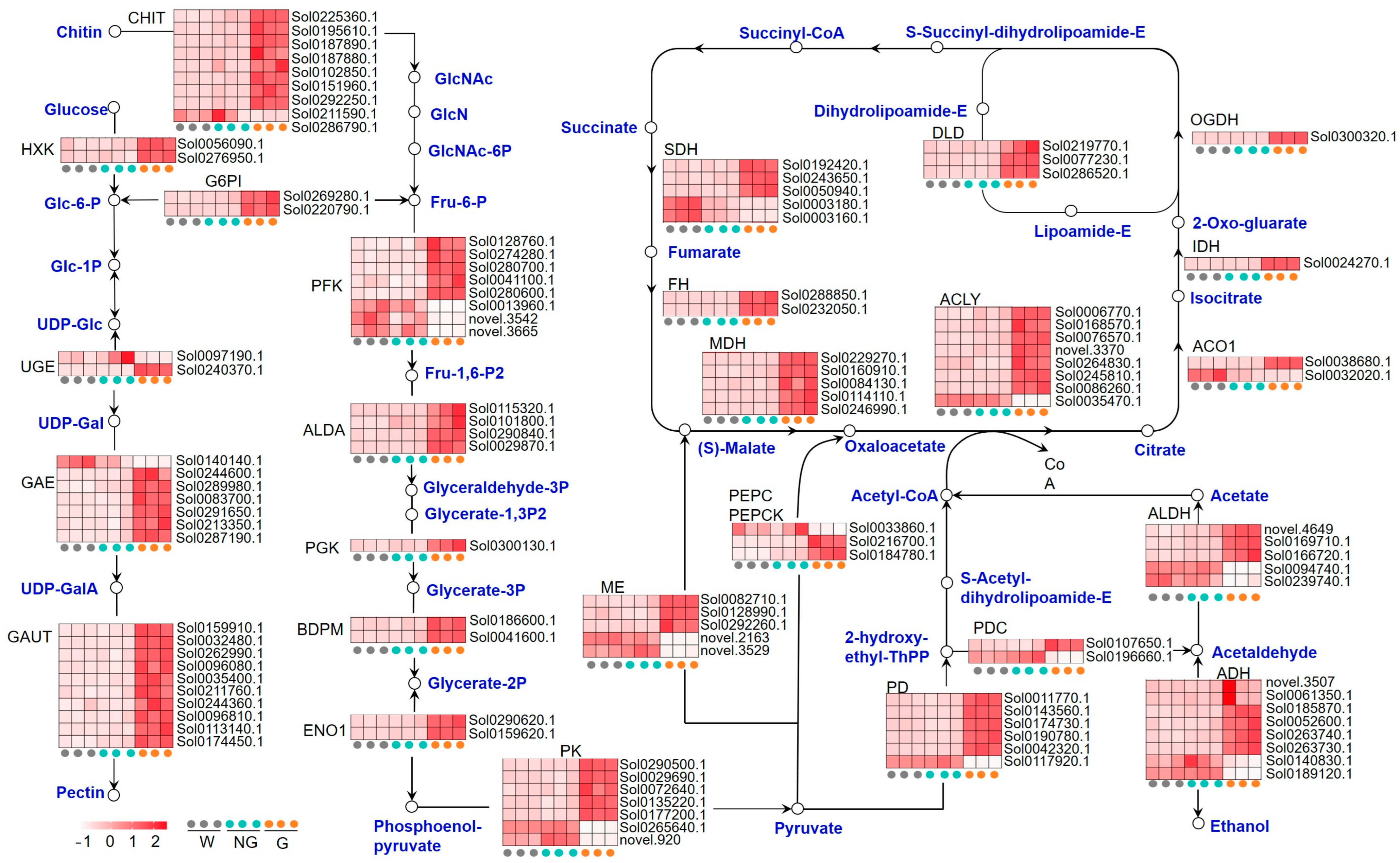
Given that energy-related metabolic pathways were highly enriched during the germination of S. torvum seeds, the GRGs related to amino sugar and nucleotide sugar metabolism, glycolysis, the citrate cycle (TCA cycle), and pyruvate metabolism were further analyzed (Figure 4, Supplemental Table S8). Expression profiling analysis on 122 GRGs encoding the key enzymes involved in these pathways indicated that most of them were up-regulated in germinated seeds (Figure 4). The expression of eight genes encoding endochitinase (CHIT), two genes encoding hexokinase (HXK), and two genes encoding UTP-glucose-1-phosphate uridylyltransferase (G6PI), with the latter two enzymes promoting the biosynthesis of Fru-6-P, was significantly enhanced in germinated seeds. Fru-6-P is the key substrate of ATP-dependent 6-phosphofructokinase (PFK) for the synthesis of the metabolite regulator, Fru-1,6-P2, which is eventually converted into pyruvate in the glycolysis process. Most of the genes encoding the key glycolytic enzymes, including PFK and pyruvate kinase (PK), were up-regulated. Up-regulated expression was also detected in the genes encoding the key enzymes involved in pyruvate metabolism, including NAD-dependent malic enzyme (ME), phosphoenolpyruvate carboxylase (PEPC), pyruvate decarboxylase (PDC), pyruvate dehydrogenase (PDH), alcohol dehydrogenase (ADH), and aldehyde dehydrogenase (ALDH). The expression of 22 TCA cycle genes encoding citrate cleavage-related enzymes, such as ATP-citrate synthase (ACYL), malate dehydrogenase (MDH), succinate dehydrogenase (SDH), and dihydrolipoyl dehydrogenase (DLD), was generally up-regulated in germinated seeds. These results suggest that energy-related metabolism is enhanced during GA-induced seed germination.
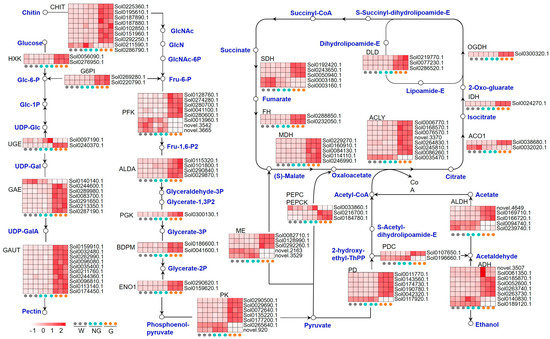
Figure 4.
Expression of the annotated GRGs encoding the key enzymes related to energy metabolism. The pathways include amino sugar and nucleotide sugar metabolism, glycolysis, citrate cycle (TCA cycle), and pyruvate metabolism. The key compounds and substrates are shown in blue color. GlcNAc, N-Acetylglucosamine; GlcN, glucosamine; GlcNAc-6P, N-acetyl-D-glucosamine 6-phosphate; Fru-6-P, Fructose 6-phosphate; Glc-6-P, Glucose 6-phosphate; Glc-1P, Glucose-1-phosphate; UDP-Glc, UDP-glucose; UDP-GalA, UDP-α-d-galacturonic acid; Fru-1,6-P2, Fructose 1,6-bisphosphate; Glyceraldehyde-3P, Glyceraldehyde 3-phosphate; Glycerate-1,3P2, glycerate-1,3-bisphosphate; Glycerate-3P, glycerate 3-bisphosphate; Glycerate-2P, glycerate 2-bisphosphate; CHIT, endochitinase; HXK, hexokinase; UGE, UDP-glucose 4-epimerase; GAE, UDP-glucuronate 4-epimerase; GAUT, galacturonosyltransferase; G6PI, UTP-glucose-1-phosphate uridylyltransferase; PFK, ATP-dependent 6-phosphofructokinase; ALDA, fructose-bisphosphate aldolase; PGK, phosphoglycerate kinase; BDPM, 2,3-bisphosphoglycerate-dependent phosphoglycerate mutase; ENO1, enolase; PK, pyruvate kinase; ME, NAD-dependent malic enzyme; PEPC, phosphoenolpyruvate carboxylase; PEPCK, phosphoenolpyruvate carboxykinase; PDC, pyruvate decarboxylase; PDH, pyruvate dehydrogenase; ADH, alcohol dehydrogenase; ALDH, aldehyde dehydrogenase; ACO1, aconitate hydratase; IDH, isocitrate dehydrogenase; ACYL, ATP-citrate synthase; OGDH, 2-oxoglutarate dehydrogenase; DLD, dihydrolipoyl dehydrogenase; SDH, succinate dehydrogenase; FH, Fumarate hydratase; MDH, malate dehydrogenase.
3.5. GRGs Related to Hormone Biosynthesis and Signaling Transduction during S. torvum Seed Germination
Although genes related to the hormone signaling transduction pathway were not detected to be highly enriched in DEGs or GRGs, several key factors involved in the hormone signaling transduction pathway were observed to be significantly influenced during GA-induced seed germination (Supplemental Figure S8). We then further analyzed the expression profile of GRGs that are related to the hormone biosynthesis and signaling transduction pathways (Supplemental Table S9). Among the 260 GRGs related to hormone biosynthesis and signaling, the major contribution came from GA, abscisic acid (ABA), auxin, and ethylene.
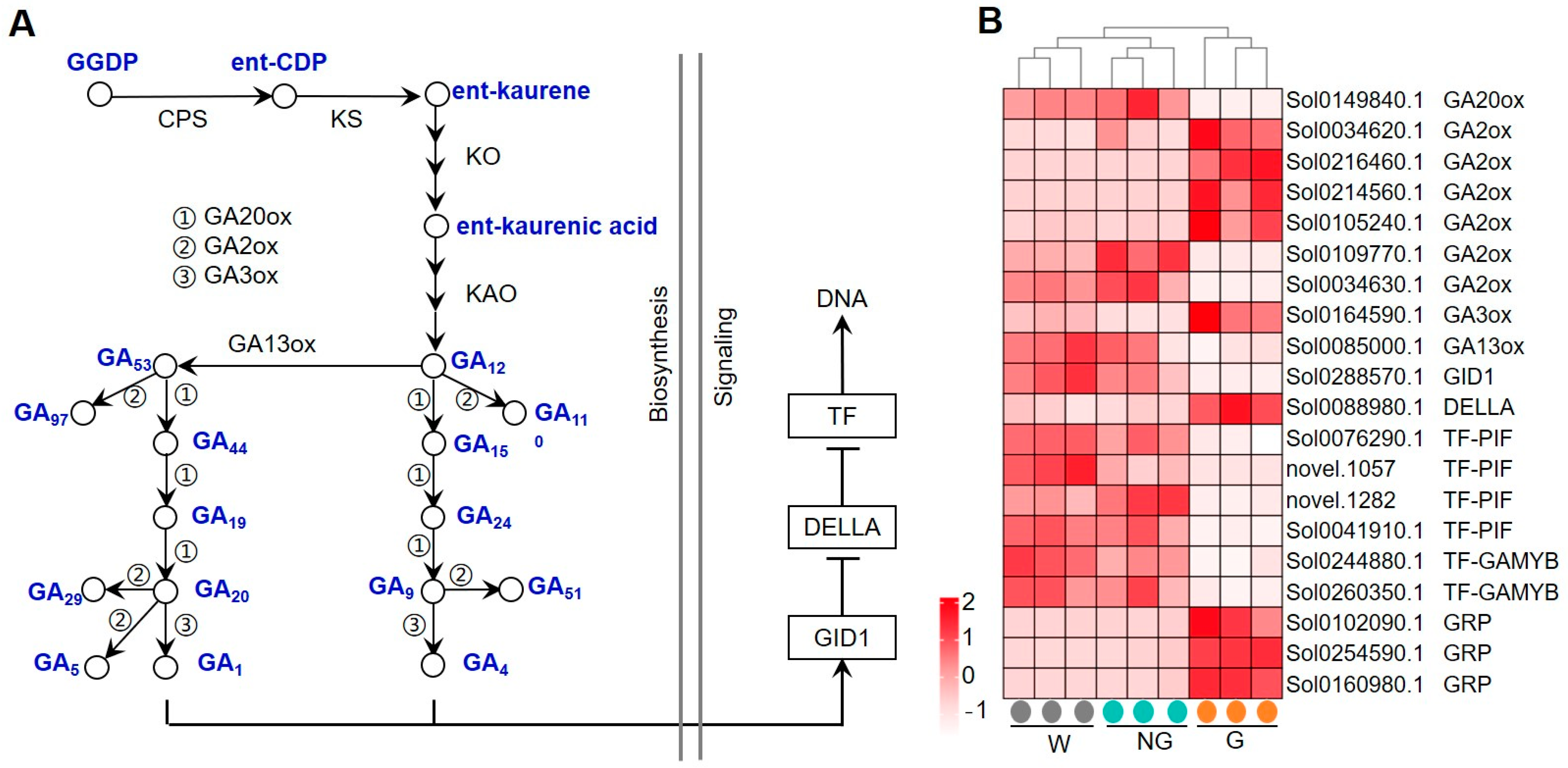
Whether exogenous application of GA affected the expression of genes involved in the GA biosynthesis and signaling pathway was firstly analyzed (Figure 5). GA biosynthesis in higher plants (Figure 5A) involves production of ent-kaurene in proplastids, the conversion of ent-kaurene to bioactive GA1, and the synthesis of various forms of GAs in the cytoplasm [31,32]. The GA signal is captured by the GA INSENSITIVE DWARF1 (GID1) receptor. The binding of GA with GID1 promotes the interaction of GID1 with DELLA proteins that mediates the degradation of DELLAs via the ubiquitin–proteasome pathway [35,36]. Thus, the transcriptional repression of GA response genes by DELLAs is released, which, in turn, induces GA signaling transduction. The expression of several genes encoding GA biosynthesis-related key enzymes such as gibberellin 20-oxidase (GA20ox), gibberellin 2-beta-dioxygenase (GA2ox), gibberellin 3-oxidase (GA3ox) and gibberellin 13-oxidase (GA13ox) was transcriptionally regulated (Figure 5B). Among these genes, Sol0149840 encoding GA20ox and Sol0085000 encoding GA13ox were significantly down-regulated, while Sol0164590 encoding GA3ox was greatly up-regulated. Most of the detected genes encoding GA2ox were transcriptionally enhanced. Several genes encoding key factors involved in GA signaling, including GID1, DELLA, and GA-responsive transcription factors (TFs), were transcriptionally regulated as well. While a GID gene was detected to be down-regulated, a gene (Sol0088980) that encodes the DELLA protein was found to be significantly up-regulated. Most GA-responsive TFs were repressed. These results suggest that exogenous GA treatment disturbed the endogenous GA biosynthesis and signaling transduction.
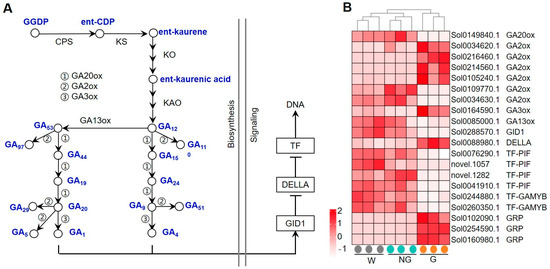
Figure 5.
Expression of GRGs encoding the key components related to GA biosynthetic and signaling pathways. (A) Diagram of the GA biosynthetic and signaling pathway. The key compounds and substrates in the GA biosynthetic pathway are shown in blue color. (B) Heatmap depicting the expression patterns of GRGs involved in GA biosynthesis and signaling. GGPP, trans-geranyl-geranyl diphosphate; ent-CDP, ent-copalyl diphosphate; CPS, copalyl diphosphate synthase; KS, ent-kaurene synthase; KO, ent-kaurene oxidase; KAO, ent-kaurenoic acid oxidase; GA13ox, gibberellin 13-oxidase; GA2ox, gibberellin 2-oxidase; GA20ox, gibberellin 20-oxidase; GA3ox, gibberellin 3-oxidase; GID1, gibberellin INSENSITIVE DWARF1; TF, transcription factor.
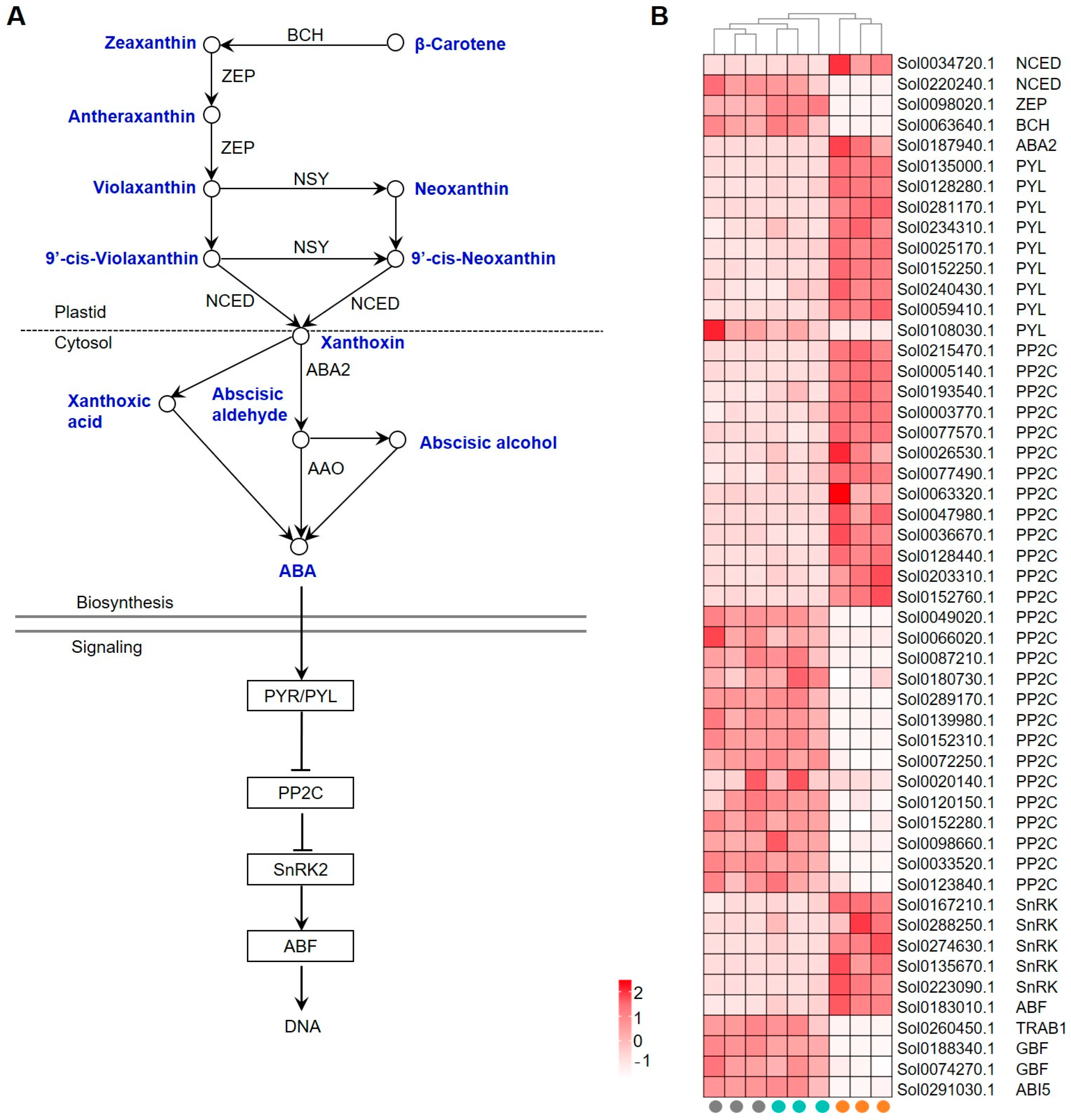
A total of 51 genes were identified to be involved in the ABA biosynthesis and signaling pathway. Xanthoxin, the intermediate metabolite during ABA biosynthesis, is synthesized from β-carotene (Figure 6A). The relative key enzymes, however, including beta-carotene hydroxylase (BCH), zeaxanthin epoxidase (ZEP), and 9-cis-epoxycarotenoid dioxygenase (NCED), were largely down-regulated (Figure 6B, Supplemental Table S9).
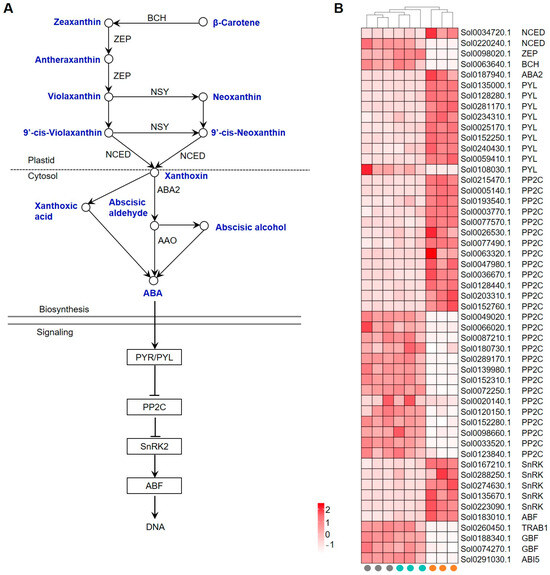
Figure 6.
Expression of GRGs encoding key components related to abscisic acid (ABA) biosynthetic and signaling pathways. (A) Diagram of ABA biosynthetic and signaling pathway. The key compounds and substrates in ABA biosynthetic pathway are shown in blue color. BCH, beta-carotene hydroxylase; ZEP, zeaxanthin epoxidase; NSY, neoxanthin synthase; NCED, 9-cis-epoxycarotenoid dioxygenase; AS-SDR, ABA-specific short-chain dehydrogenase/reductase; AAO, ABA-specific aldehyde oxidase. (B) Heatmap depicting the expression patterns of GRGs involved in ABA biosynthesis and signaling. TRAB1, Transcription factor responsible for ABA regulation 1; ABI5, ABSCISIC ACID INSENSITIVE5.
Core components in ABA signaling involve the receptor PYR/PYL, the negative regulator PP2C, the positive regulator SnRK, and the downstream responsive TFs. ABA activates its receptors, PYR/PYL family proteins, to inhibit the activity of PP2Cs on SnRKs, which activates SnRKs to stimulate the ABA response [37]. While most of the identified GRGs encoding PYL and SnRKs were transcriptionally up-regulated in germinated seeds, both up- and down-regulation of PP2C coding genes were observed (Figure 6B). Some ABA-responsive TFs, such as ABSCISIC ACID INSENSITIVE5 (ABI5), G-box binding factor (GBF), and Transcription factor responsible for ABA regulation 1 (TRAB1), were found to be down-regulated. These results suggest that ABA biosynthesis and signaling may be repressed during GA-induced seed germination.
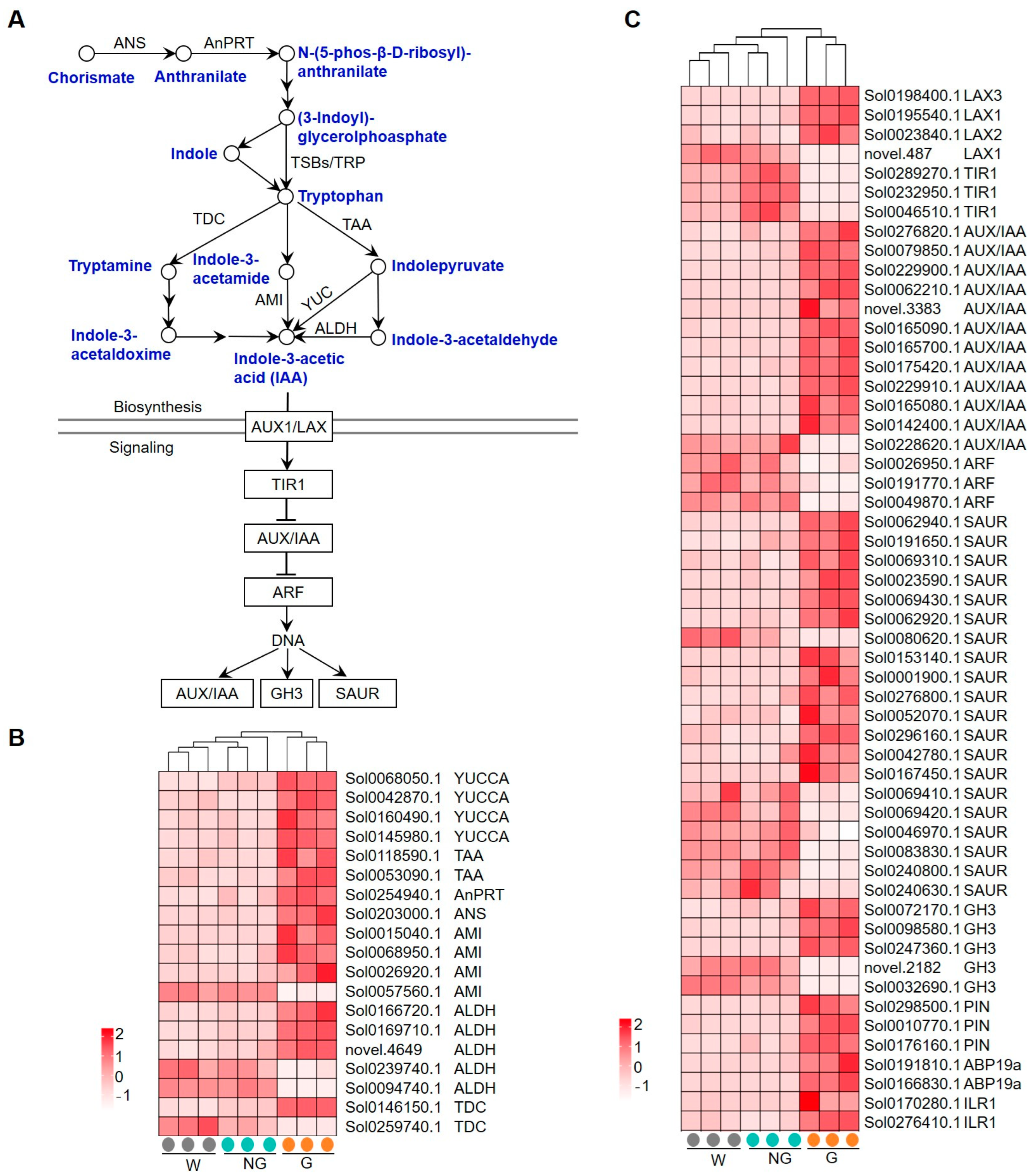
The expression patterns of GRGs involved in auxin biosynthesis and signaling are shown in Figure 7. Auxin is mainly produced via a tryptophan-dependent pathway. Tryptophan, as the auxin precursor, is synthesized from chorismate via the shikimate pathway (Figure 7A). A total of 19 genes encoding auxin biosynthetic enzymes, including flavin-containing monooxygenase (YUCCA), L-tryptophan–pyruvate aminotransferase (TAA), anthranilate phosphoribosyltransferase (AnPRT), anthranilate synthase (ANS), amidase (AMI), aldehyde dehydrogenase (ALDH), and tyrosine decarboxylase (TDC), were found to be transcriptionally regulated. While the expression of most of these genes was significantly enhanced (Figure 7B), one AMI-coding gene and two ALDH-coding genes were among the down-regulated GRGs. Auxin transport is carrier-mediated. AUXIN1/LIKE-AUX1 (AUX1/LAX) family members function as auxin influx carriers, while PIN-FORMED (PIN) family members are the major auxin efflux carriers [38]. The expression of six genes encoding LAX and PIN family proteins was observed to be enhanced in germinated seeds (Figure 7C). Three core signaling components, including the receptors TIR1/AFBs, the transcriptional repressors Aux/IAAs, and the transcription factors ARFs, mediate the transcriptional response to auxin in the nucleus [39,40,41,42,43]. A large number of genes encoding the core components involving auxin signaling, including TIR1, AUX/IAA, ARF, small auxin up-regulated RNA (SAUR), and GRETCHEN HAGEN 3 (GH3), were found to be transcriptionally regulated in germinated seeds (Figure 7C). While the majority of early auxin responsive genes like AUX/IAA, SAUR, and GH3 were found to be transcriptionally enhanced, TIR1-coding genes were significantly repressed. These results suggest that auxin biosynthesis and signaling may play a role in response to GA-induced seed germination.
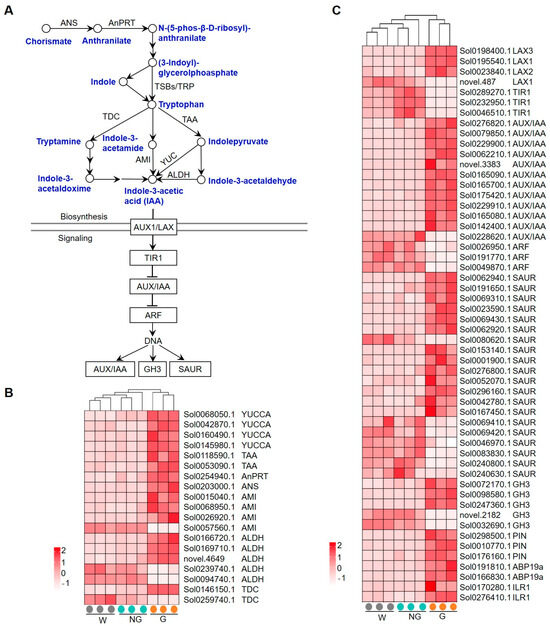
Figure 7.
Expression of GRGs encoding the key components related to auxin biosynthetic and signaling pathways. (A) Diagram of auxin biosynthetic and signaling pathway. The key compounds and substrates in auxin biosynthetic pathway are shown in blue color. ANS, anthranilate synthase; AnPRT, anthranilate phosphoribosyltransferase; TSB, Trp synthase α; TSB, Trp synthase β; TAA, L-tryptophan–pyruvate aminotransferase; TDC, tyrosine decarboxylase; AMI, amidase; YUCCA, flavin-containing monooxygenase; ALDH, aldehyde dehydrogenase; Aux1/LAX, AUXIN1/LIKE-AUX1; TIR1, TRANSPORT INHIBITOR RESPONSE 1; AUX/IAA, AUXIN/INDOLE ACETIC ACID; ARF, Auxin response factor; GH3, GRETCHEN HAGEN 3; SAUR, small auxin up-regulated RNA. (B,C) Heatmaps showing the expression patterns of GRGs involved in auxin biosynthesis and signaling, respectively. PIN, PIN-FORMED; ABP, auxin-binding protein; ILR1, IAA-amino acid hydrolase.
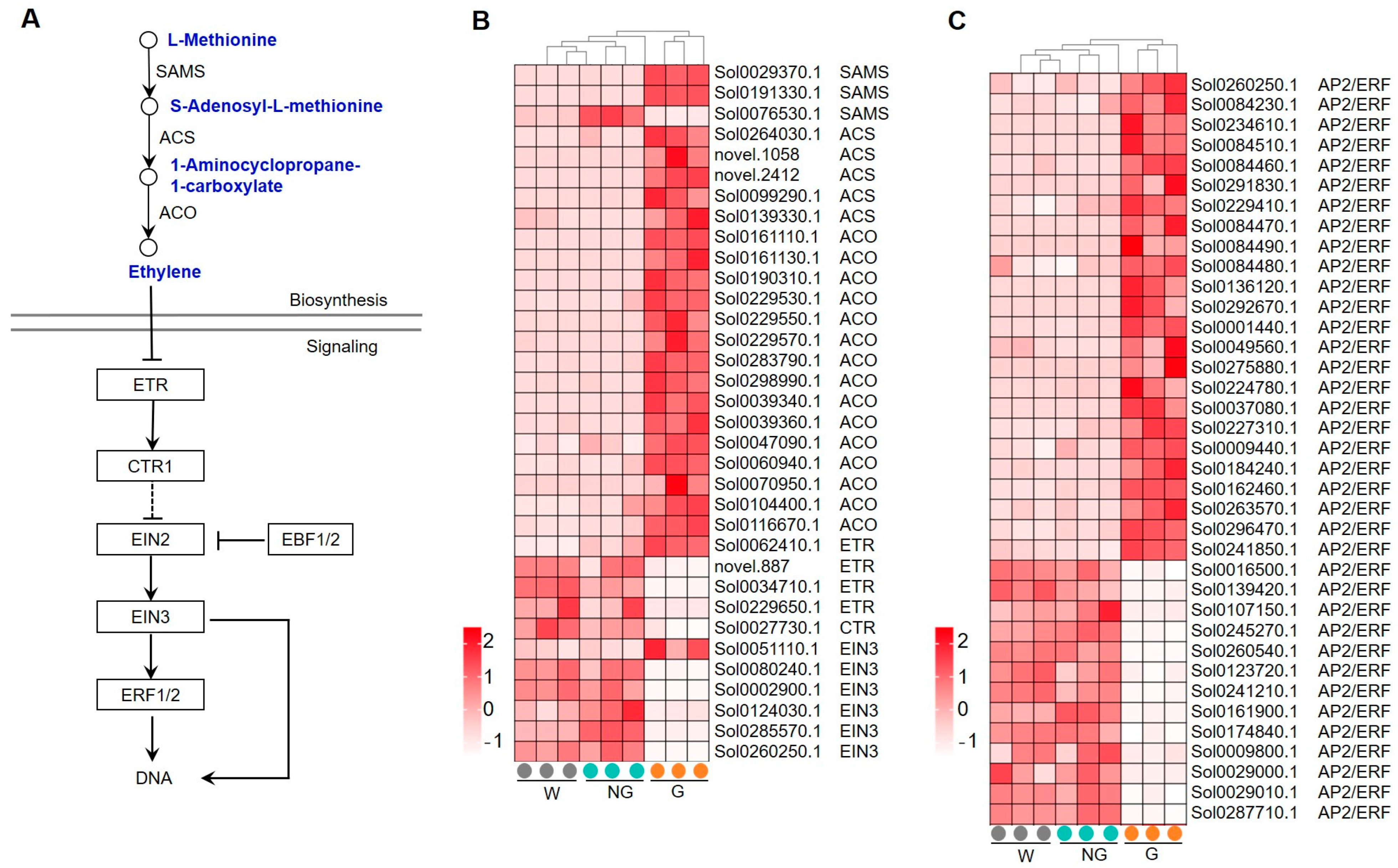
Another enriched hormone signaling pathway in GRGs is ethylene. Ethylene is synthesized from L-methionine by three key steps catalyzed by S-adenosylmethionine synthase (SAMS), 1-aminocyclopropane-1-carboxylate synthase (ACS), and 1-aminocyclopropane-1-carboxylate oxidase (ACO) (Figure 8A). As shown in Figure 8B, most of the genes encoding SAMS, ACS, and ACO were significantly up-regulated, suggesting an enhancement of ethylene biosynthesis in GA-induced germinated seeds. Core components in ethylene signaling, such as ethylene receptors (ETRs), constitutive triple response 1 (CTR1), ethylene-insensitive 2 (EIN2), ethylene-insensitive 3 (EIN3), and ethylene response factors (ERFs), were also analyzed (Figure 8B). Ethylene functions as an inverse agonist to its receptor ETR [44], which releases the inhibitory activity of CTR1 on the downstream ethylene responsive transcription. While the expression of the identified ETR and CTR genes were found to be significantly decreased in germinated seeds, most of the ethylene-responsive transcription factors EIN3 (Figure 8B) and ERF (Figure 8C) were activated. All these results suggest that both ethylene biosynthesis and signaling pathways were enhanced during GA-induced seed germination.
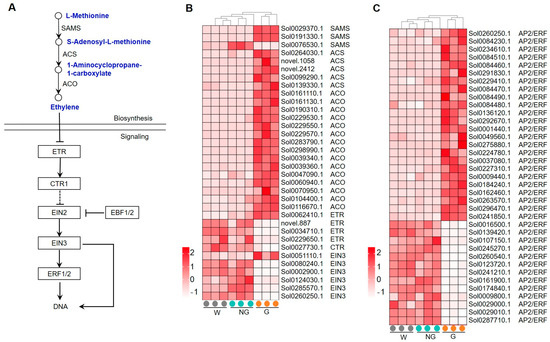
Figure 8.
Expression of GRGs encoding the key components related to ethylene biosynthetic and signaling pathways. (A) Diagram of ethylene biosynthetic and signaling pathway. The key compounds and substrates in ethylene biosynthetic pathway are shown in blue color. SAMS, S-adenosylmethionine synthase; ACS, 1-aminocyclopropane-1-carboxylate synthase; ACO, 1-aminocyclopropane-1-carboxylate oxidase; ETR, ethylene receptor; CTR1, constitutive triple response 1; EIN2, ethylene-insensitive 2; EIN3, ethylene-insensitive 3; ERF, ethylene response factor. (B) Heatmap showing the expression patterns of GRGs involved in ethylene biosynthesis and signaling. (C) Heatmap depicting the expression patterns of the identified AP2/ERFs.
The genes encoding jasmonate (JA) and salicylic acid (SA) signaling-related components, such as jasmonic acid-amido synthetase (JAR1) and JASMONATE ZIM-DOMAIN (JAZ) in JA signaling and nonexpressor of pathogenesis-related 1 (NPR1), transcription factor TGA1, and salicylic acid-binding protein 2 (SABP) in SA signaling, were mostly up-regulated (Supplemental Table S9). Most A-type ARABIDOPSIS RESPONSE REGULATOR (ARR) family members that mediate the cytokinin signaling response were up-regulated, while differential expression of brassinosteroid (BR)-insensitive 1 (BRI1) and BR-regulated transcription factor BZR1 in BR signaling was also observed (Supplemental Table S9). Collectively, these results suggest that hormone signaling pathways are significantly influenced during GA-induced seed germination.
3.6. Possible TFs Responsible for Seed Germination
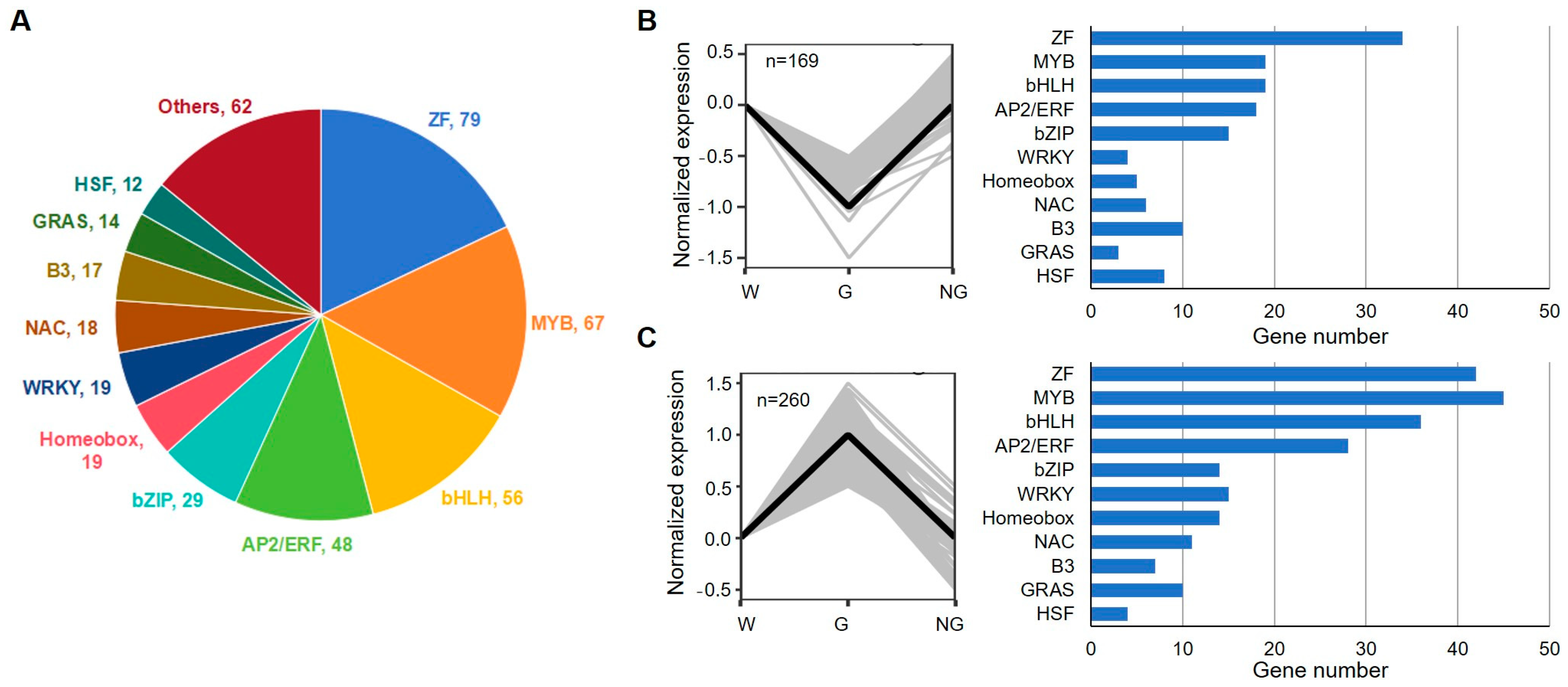
A total of 440 TFs were identified in GRGs (Figure 9, Supplemental Table S10). Among those TFs, 79 TFs belong to zinc finger motif-containing factors, 67 MYB family members, and 56 bHLH family members (Figure 9A). Based on their expression in germinated seeds in comparison with ungerminated seeds, the TFs were clustered into two groups. A total of 169 TF coding genes showed decreased expression while the expression of 260 TFs was increased in germinated seeds (Figure 9B,C). These genes may be responsible for transcriptional reprogramming during GA-induced seed germination.
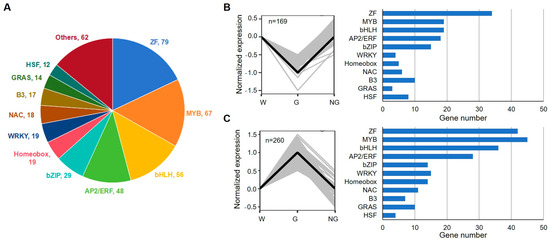
Figure 9.
Numbers of transcription factors (TFs) from the main TF families identified during germination of S. torvum seeds. (A) Pie chart showing the distribution of TFs from different families. (B,C) Expression pattern (left panel) and gene numbers (right panel) of down- (B) and up-regulated (C) TFs in germinated seeds. Gray lines in left panels represent the normalized expression level of each gene, while black thick lines represent the median centroids.
3.7. Genes That Are Responsible for Seed Dormancy and Germination
We further analyzed the expression of known genes encoding the key factors responsible for seed dormancy and germination (Table 1), including Delay of Germination-1 (DOG1), Dormancy-associated DRM1/APR-like protein, FUSCA 3 (FUS3), LEAFY COTYLEDON 1 (LEC1), chromatin-remodeling factor PICKLE (PKL)-related protein, PHYTOCHROME INTERACTING FACTOR1 (PIF1), and REVEILLE 1 (RVE1). Transcriptome data revealed that their expression was decreased during GA-induced seed germination compared to that observed in ungerminated seeds. For example, DOG1-like protein coding gene (Sol0034020.1) was expressed in H2O-treated ungerminated seeds with an average FPKM of 90, which was decreased to 1 in germinated seeds (Supplemental Table S6) with a log2 fold change of −6.5 (Table 1).

Table 1.
Expression of known genes related to seed dormancy and germination.
4. Discussion
Seed germination is a complex process involving dormancy breaking, embryo recovery from drying, seed coat rupture, radicle elongation, and germ transformation into seedlings [45,46]. The process is regulated by the coordination of different external environment and endogenous signals. A variety of techniques are often used to treat seeds to induce germination. For example, Arabidopsis seeds must be treated at low temperatures to induce germination [47]. Dufková et al. [48] proposed that hydrogen peroxide and temperature treatment improved seed germination. In addition, some physical methods, such as hand-dehulling seeds, can also effectively promote seed germination [49].
GA is an essential plant hormone which is mainly synthesized in buds, young roots, and immature seeds and is involved in many biological processes such as seed germination, flowering, and fruit development [26]. Exogenous treatment of the plant phytohormone GA is effective in promoting seed germination in a variety of crops, such as Arabidopsis [19], rice [50], and Fraxinus hupehensis [51]. In the study of peony, apple, poplar, pear, grape, etc., exogenous spraying of GA was used to replace low temperatures to break seed dormancy [52].
A previous study [14] revealed that although less efficient than GA, application of KNO3 had positive effects in releasing S. torvum seeds from dormancy and stimulating germination. However, in our study, KNO3 was not effective while GA was required to induce the germination of S. torvum seeds. It was suggested that the positive effects of chemical substances, such as KNO3, are not always observed due to their abilities to decrease the osmotic pressure of the substrate, thereby precluding imbibition [15,53]. The germination induced by KNO3 may require further investigation. It is worth mentioning that while GA helped to break the dormancy of S. torvum seeds, seedling emergence was not uneven. Eight days were required to reach the highest germination rate, with about 15% of the seeds remaining ungerminated after 12 days (Figure 1). Similar results were reported in previous studies [14,54]. While a test on multiple factors revealed an emergence rate of only 77% after 14 days [14], the highest germination of 81% was detected when GA treatment was combined with altering temperature [54]. Thus, to further improve the germination of S. torvum seeds for usage as a rootstock and for breeding purposes, the underlying mechanism for efficient germination rates needs to be investigated [7,16].
RNA sequencing allows for a quantitative view of gene expression at the transcript level. The transcriptome reprogramming reflected by RNA sequencing enables the investigation of the alteration of key regulation pathways during a specific biological process. The present study utilizes RNA-seq to investigate the effects of GA on the germination of S. torvum seeds.
Analysis of the detected DEGs and GRGs revealed that energy-producing metabolic pathways, including amino sugar and nucleotide sugar metabolism, the biosynthesis of nucleotide sugars, the citrate cycle (TCA cycle), carbon metabolism, and pyruvate metabolism, were largely elevated during the germination of S. torvum seeds. This result aligns with previous studies showing that glycolysis, the tricarboxylic acid cycle (TCA cycle), and amino acid metabolism play key roles in seed germination [55,56]. The enhancement of carbon metabolism reflects the utilization of stored sugars and starches in seeds during germination [57], which provide a large amount of energy for seed germination. Glycolysis and alcoholic fermentation are induced under limited oxygen content during seed imbibition [58]. They provide additional energy to satisfy the energy demands of germinating seeds. Consistent with this view, several genes encoding the rate-limiting enzymes in glycolysis such as HXK, PFK, and PK, as well as ethanol fermentation related-enzymes such as PDC and ADH, were up-regulated in GA-induced germinated seeds (Figure 4). Likewise, the TCA cycle was promoted in germinated seeds, which produces a large amount of energy (ATPs) and reduces the power (NADH and FADH2) required for seed germination (Figure 4). The rate-limiting enzyme in the TCA cycle, IDH, was also found to be activated (Figure 4). Meanwhile, the expression of genes encoding the vital enzyme, PDH, that links anaerobic glycolysis to the aerobic TCA cycle was largely activated (Figure 4), which boosts pyruvate metabolism to produce a large amount of acetyl-CoA as the major substrate of the TCA cycle. The mobilization of these metabolic pathways is to provide sufficient energy for seed germination. Such metabolic processes were not activated in ungerminated seeds.
The changes in energy metabolism may be regulated by exogenous GA treatment. Duan et al. [59] showed that GA promoted early seed germination by influencing physiological and metabolic activities of seeds. In a study on potato tubers, application of GA was found to induce the activity of α-amylases and β-amylases [60]. GA can also promote the synthesis of nucleic acid and protein by activating hydrolytic enzymes in seeds, thus promoting the germination of plant seeds. It was found that the active Cys peptide sites of reduced glutathione reductase 2, NADPH-thioredoxin reductase a/b, and thioredoxin-o1 acted as redox switches to induced early response during early germination [61]. Their mutations resulted in an enhancement in the oxygen consumption rate and dysregulation of TCA cycle dynamics and significantly reduced the seed germination rate [61]. Therefore, we hypothesize that the changes in energy metabolism-related genes during the germination of S. torvum seeds is caused by exogenous addition of GA.
However, exogenous addition of GA appears to affect endogenous GA biosynthesis and signaling transduction. The genes encoding enzymes involved in early steps of GA biosynthesis, i.e., CPS, KS, KO, and KAO, were not differentially expressed between germinated and ungerminated seeds. On the other hand, however, the enzymes in the third stage of GA biosynthesis, such as GA2ox, GA3ox, GA20ox, and GA13ox, were highly enriched in GRGs (Figure 5). Among them, GA20ox is the major rate-limiting enzyme in GA biosynthesis, which catalyzes the formation of active GAs. Its expression was greatly depressed in the GA-induced germinated seeds (Figure 5B). Meanwhile, germination enhanced the expression of four genes encoding GA2ox (Figure 5B), which diverges early intermediates from the biosynthesis pathway of active GA forms [62]. It was reported that overexpression of GA2ox significantly affected the germination rate of tomato seeds [63]. The depression of the endogenous biosynthesis of active GA might be a feedback inhibition mediated by the exogenous addition of GA. Consequently, GA signaling transduction is depressed, reflected by the down-regulation of a gene encoding gibberellin receptor GID1 and the up-regulation of the transcriptional repressor in GA signaling, DELLA (Figure 5B) [64,65].
Hormones are widely accepted to be vital in the regulation of seed dormancy and germination. GA and ABA are the two classic phytohormones that antagonistically regulate seed dormancy and germination [66]. While GA stimulates seed germination [67,68,69,70], ABA is involved in the establishment and maintenance of dormancy to inhibit seed germination by restricting the availability of energy and metabolites [71,72]. Seeds of the Arabidopsis ga1-3 mutant, which is deficient in GA biosynthesis, are unable to germinate [73]. On the other hand, artificial induction of ABA synthetase NCED6 and NCED9 was found to significantly delay seed germination [74]. Thus, a balance between ABA and GA is critical for the initiation of seed germination. In this study, exogenous GA-induced seed germination caused the down-regulation of more than 30 genes encoding the major enzymes involved in ABA biosynthesis in plastids, including NCED, ZEP, and BCH, suggesting the inhibition of ABA biosynthesis during seed germination. Meanwhile, the expression of a large number of genes coding the core components of ABA signaling, including PYL, PP2C, and SnRK2, were also disturbed in germinated S. torvum seeds (Figure 6).
Auxin is another hormone required for seed development and dormancy. Exogenous auxin regulates the secondary dormancy and germination of tobacco seeds [75]. Auxin is considered to promote seed dormancy and inhibit seed germination by increasing ABA content while impairing the biosynthesis of active GA forms [76,77]. The crosstalk between auxin and ABA is also reflected by the application of fluridone, an inhibitor of ABA biosynthesis, which reverses the auxin-mediated delay in germination [77]. On the other hand, once seed dormancy is broken, auxin is required to promote hypocotyl elongation [78,79]. We noticed that most of the identified genes encoding IAA biosynthetic enzymes were up-regulated (Figure 7), and more than 50 genes encoding the auxin transporters LAXs and PINs and core components in auxin signaling such as AUX/IAA and SAUR were significantly enhanced. These results suggest that auxin biosynthesis and signaling are activated to promote seedling development after the dormancy of S. torvum seeds is broken by exogenous GA.
We also found that the expression of a large number of genes involved in the biosynthesis of other hormones and signaling transduction, including ethylene, cytokinin, BR, JA, and SA, was altered during GA-induced germination of S. torvum seeds. For example, the key enzymes responsible for ethylene biosynthesis from methionine, including SAMS, ACS, and ACO, were largely activated (Figure 8), suggesting that ethylene biosynthesis is elevated in germinating seeds. Ethylene has been reported to play an important role in dormancy release and seed germination in many species [80]. Its regulatory role is considered to engage in crosstalk with other hormones, such as GA and ABA [80,81,82]. A similar situation may apply to other hormones as well [83,84,85,86]. It was reported that interactions among JAZ, ARF, and ABSCISIC ACID INSENSITIVE5 (ABI5) contribute to the crosstalk of JA, auxin, and ABA during the germination of Arabidopsis seeds [87]. ARFs and ABI5 are the core transcription factors of auxin and the ABA signaling pathway, respectively. ABI5 accumulation was reported to inhibit seed germination and early seedling establishment [88,89,90]. ARF 10 and ARF16 were reported to physically interact with ABI5, which activates the transcriptional function of ABI5 [87]. JAZ protein, the main inhibitor of the JA signaling pathway, competitively inhibits the interaction of ARF10/ARF16-ABI5 and thereby impairs ABI5 transcription [87,91]. Therefore, JA-mediated regulation of ABA signaling during seed germination may be synergistically regulated by auxin. We noticed that the expression of a gene encoding ABI5 was significantly down-regulated in germinating seeds (Figure 7). Although the specific regulatory mechanism needs further study, auxin and JA signaling may cooperate to regulate the expression of ABI5 during the germination of S. torvum seeds.
Seed dormancy-related genes were also investigated in this study (Table 1). DELAY OF GERMINATION1 (DOG1) gene is a master genetic regulator in determining the depth of seed dormancy [92]. Their amount is closely correlated with the dormancy level of freshly harvested seeds [93], and depression of DOG1 genes usually promotes seed germination [94]. In this study, we identified several DOG-coding genes that may be involved in the dormancy release and germination of S. torvum seeds (Table 1). Their expression was significantly down-regulated in germinated seeds but not in ungerminated seeds. This result suggests that the dormancy of GA-treated ungerminated seeds was not broken. We propose that the expression of DOG1-like genes may be employed as a dormancy indicator of S. torvum seeds. The regulatory role of DOG1 is considered to echo with ABA signaling, which affects the expression of ABI5 to regulate seed germination [95]. Given that the expression pattern of ABI5 was similar to that of DOG1 genes (Figure 6, Table 1), ABI5 may be a downstream target of DOG1 to regulate the germination of S. torvum seeds.
Other key regulators in seed dormancy and germination, such as Dormancy-associated DRM1-like protein, FUS3, LEC1, SOMNUS, and PIF1, showed a similar expression pattern with that of DOG1 (Table 1). DRM1 was found to be highly expressed in dormant seeds, while its ectopic expression delayed the germination of Arabidopsis seeds [96]. Previous studies reported that the expression of DOG1 relies directly or indirectly on FUS3 and LEC1 [97,98,99], which may involve SOMNUS and ABI3 in coordination with ABA signaling [95,100]. PIF1 interacts with REVEILLE 1 (RVE1) and they act as negative regulators in light-mediated seed germination via depression of the expression of GA3ox and thus the biosynthesis of active GA [101]. Thus, the down-regulation of these genes (Table 1) may contribute to the up-regulation of GA3ox (Figure 5) in germinated S. torvum seeds.
5. Conclusions
Exogenous application of 1.5 mM GA could effectively promote the germination of S. torvum seeds. GA treatment induces comprehensive changes in the expression of genes involved in energy metabolism, hormone biosynthesis, and signaling transduction. This study identified a large number of potential genes that are responsible for S. torvum seed germination, which provides theoretical basis for further research on the promotion of seed germination by GA.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/agronomy14081736/s1: Figure S1: Germination percentage of S. torvum seeds under different treatments; Figure S2: Seed samples collected for RNA-seq analysis; Figure S3: Transcriptome analysis of germinated and ungerminated S. torvum seeds; Figure S4: KEGG pathways enriched in DEGs identified in each comparison group; Figure S5: Top 20 GO terms enriched in the DEGs in G group compared to W group; Figure S6: Top 20 GO terms enriched in the DEGs in G group compared to NG group; Figure S7: Top 20 GO terms enriched in the DEGs in NG group compared to W group; Figure S8: Overview of plant hormone signaling transduction pathway via KEGG analysis; Table S1: Statistics of S. torvum seed germination induced by 1.5 mM GA; Table S2: Statistics of RNA-seq read filter and mapping; Table S3: Raw data of gene expression; Table S4: List of novel genes identified by RNA-seq; Table S5: Statistics of PCA on RNA-seq data; Table S6: List of differentially expressed genes (DEGs); Table S7: List of potential 9723 genes induced by GA and responsible for germination; Table S8: List of GRGs involved in energy metabolism; Table S9: List of GRGs involved in hormone biosynthesis and signaling transduction; Table S10: List of transcription factors responsible for GA-induced germination of S. torvum.
Author Contributions
Conceptualization, L.T. and Y.Y.; methodology and validation, L.T. and Y.Y.; formal analysis, S.W., Y.Y. and L.T.; investigation, Q.S., K.Y., W.Z. and Y.Y.; data curation, L.Z.; writing—original draft preparation, S.W., Y.Y. and L.T.; writing—review and editing, L.Z. and T.W.O.; supervision, L.T.; project administration, L.T.; funding acquisition, L.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the “Pioneer” and “Leading Goose” R&D Program of Zhejiang (Grant No 2022C02051); the Zhejiang A&F University Starting Funds of Scientific Research and Development, Grant Nos 203402000101, 203402000501 and 203402001301; and the Student Scientific research training program of Zhejiang A&F University (Grant No S202210341138).
Data Availability Statement
The original data presented in the study are openly available in the the Sequence Read Archive (SRA) database of the National Center for Biotechnology Information (NCBI) (https://www.ncbi.nlm.nih.gov/sra, accessed on 22 July 2024) with reference number PRJNA1138451.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- FAO. Agricultural Production Statistics 2000–2022; FAOSTAT Analytical Briefs; FAO: Rome, Italy, 2023; Volume 79. [Google Scholar]
- Daunay, M.C. Eggplant. In Vegetables II: Fabaceae, Liliaceae, Solanaceae, and Umbelliferae; Prohens, J., Nuez, F., Eds.; Springer: New York, NY, USA, 2008; pp. 163–220. [Google Scholar]
- Kaniyassery, A.; Thorat, S.A.; Kiran, K.R.; Murali, T.S.; Muthusamy, A. Fungal diseases of eggplant (Solanum melongena L.) and components of the disease triangle: A review. J. Crop Improv. 2023, 37, 543–594. [Google Scholar] [CrossRef]
- Loganathan, M.; Rai, A.B.; Ramesh, R.; Sharma, B.K.; Rai, R.K.; Rai, M. Vascular wilt diseases-a menace in vegetable crops. Veg. Sci. 2009, 36, 1–3. [Google Scholar]
- Bletsos, F.; Thanassoulopoulos, C.; Roupakias, D. Effect of grafting on growth, yield, and Verticillium wilt of eggplant. HortScience 2003, 38, 183–186. [Google Scholar] [CrossRef]
- Miceli, A.; Sabatino, L.; Moncada, A.; Vetrano, F.; D’Anna, F. Nursery and field evaluation of eggplant grafted onto unrooted cuttings of Solanum torvum Sw. Sci. Hortic. 2014, 178, 203–210. [Google Scholar] [CrossRef]
- Gisbert, C.; Prohens, J.; Raigón, M.D.; Stommel, J.R.; Nuez, F. Eggplant relatives as sources of variation for developing new rootstocks: Effects of grafting on eggplant yield and fruit apparent quality and composition. Sci. Hortic. 2011, 128, 14–22. [Google Scholar] [CrossRef]
- Gousset, C.; Collonnier, C.; Mulya, K.; Mariska, I.; Rotino, G.L.; Besse, P.; Servaes, A.; Sihachakr, D. Solanum torvum, as a useful source of resistance against bacterial and fungal diseases for improvement of eggplant. Plant Sci. 2005, 168, 319–327. [Google Scholar] [CrossRef]
- Brenes, M.; Pérez, J.; González-Orenga, S.; Solana, A.; Boscaiu, M.; Prohens, J.; Plazas, M.; Fita, A.; Vicente, O. Comparative Studies on the Physiological and Biochemical Responses to Salt Stress of Eggplant (Solanum melongena) and Its Rootstock, S. torvum. Agriculture 2020, 10, 328. [Google Scholar] [CrossRef]
- Xu, J.; Zhu, Y.; Ge, Q.; Li, Y.; Sun, J.; Zhang, Y.; Liu, X. Comparative physiological responses of Solanum nigrum and Solanum torvum to cadmium stress. New Phytol. 2012, 196, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Bletsos, F.A.; Olympios, C.M. Rootstocks and grafting of tomatoes, peppers and eggplants for soil-borne disease resistance, improved yield and quality. Eur. J. Plant Sci. Biotech. 2008, 2, 62–73. [Google Scholar]
- Öçal, S.; Özalp, T.; Devran, Z. Reaction of wild eggplant Solanum torvum to different species of root-knot nematodes from Turkey. J. Plant Dis. Prot. 2018, 125, 577–580. [Google Scholar] [CrossRef]
- Petran, A.; Hoover, E. Solanum torvum as a Compatible Rootstock in Interspecific Tomato Grafting. J. Hortic. 2014, 1, 103. [Google Scholar] [CrossRef]
- Ranil, R.H.; Niran, H.M.; Plazas, M.; Fonseka, R.M.; Fonseka, H.H.; Vilanova, S.; Andújar, I.; Gramazio, P.; Fita, A.; Prohens, J. Improving seed germination of the eggplant rootstock Solanum torvum by testing multiple factors using an orthogonal array design. Sci. Hortic. 2015, 193, 174–181. [Google Scholar] [CrossRef]
- Cutti, L.; Kuluckzynski, S.M. Treatment of Solanum torvum seeds improves germination in a batch-dependent manner. Pesqui. Agropecuária Trop. 2016, 46, 464–469. [Google Scholar] [CrossRef]
- Hayati, N.E.; Sukprakarn, S.; Juntakool, S. Seed germination enhancement in Solanum stramonifolium and Solanum torvum. Agric. Nat. Resour. 2005, 39, 368–376. [Google Scholar]
- Debeaujon, I.; Koornneef, M. Gibberellin requirement for Arabidopsis seed germination is determined both by testa characteristics and embryonic abscisic acid. Plant Physiol. 2000, 122, 415–424. [Google Scholar] [CrossRef]
- Hauvermale, A.L.; Steber, C.M. GA signaling is essential for the embryo-to-seedling transition during Arabidopsis seed germination, a ghost story. Plant Signal. Behav. 2020, 15, 1705028. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, Y.; Ogawa, M.; Kuwahara, A.; Hanada, A.; Kamiya, Y.; Yamaguchi, S. Activation of gibberellin biosynthesis and response pathways by low temperature during imbibition of Arabidopsis thaliana seeds. Plant Cell 2004, 16, 367–378. [Google Scholar] [CrossRef]
- Gubler, F.; Hughes, T.; Waterhouse, P.; Jacobsen, J. Regulation of dormancy in barley by blue light and after-ripening: Effects on abscisic acid and gibberellin metabolism. Plant Physiol. 2008, 147, 886–896. [Google Scholar] [CrossRef]
- Kashiwakura, Y.; Kobayashi, D.; Jikumaru, Y.; Takebayashi, Y.; Nambara, E.; Seo, M.; Kamiya, Y.; Kushiro, T.; Kawakami, N. Highly Sprouting-Tolerant Wheat Grain Exhibits Extreme Dormancy and Cold Imbibition-Resistant Accumulation of Abscisic Acid. Plant Cell Physiol. 2016, 57, 715–732. [Google Scholar] [CrossRef]
- Liu, X.; Hou, X. Antagonistic Regulation of ABA and GA in Metabolism and Signaling Pathways. Front. Plant Sci. 2018, 9, 251. [Google Scholar] [CrossRef]
- Cao, D.; Cheng, H.; Wu, W.; Soo, H.M.; Peng, J. Gibberellin mobilizes distinct DELLA-dependent transcriptomes to regulate seed germination and floral development in Arabidopsis. Plant Physiol. 2006, 142, 509–525. [Google Scholar] [CrossRef] [PubMed]
- Groot, S.P.; Karssen, C.M. Gibberellins regulate seed germination in tomato by endosperm weakening: A study with gibberellin-deficient mutants. Planta 1987, 171, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Mitchum, M.G.; Yamaguchi, S.; Hanada, A.; Kuwahara, A.; Yoshioka, Y.; Kato, T.; Tabata, S.; Kamiya, Y.; Sun, T.P. Distinct and overlapping roles of two gibberellin 3-oxidases in Arabidopsis development. Plant J. 2006, 45, 804–818. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Harberd, N.P. The role of GA-mediated signalling in the control of seed germination. Curr. Opin. Plant Biol. 2002, 5, 376–381. [Google Scholar] [CrossRef]
- Shah, S.H.; Islam, S.; Mohammad, F.; Siddiqui, M.H. Gibberellic Acid: A Versatile Regulator of Plant Growth, Development and Stress Responses. J. Plant Growth Regul. 2023, 42, 7352–7373. [Google Scholar] [CrossRef]
- Gong, D.; He, F.; Liu, J.; Zhang, C.; Wang, Y.; Tian, S.; Sun, C.; Zhang, X. Understanding of Hormonal Regulation in Rice Seed Germination. Life 2022, 12, 1021. [Google Scholar] [CrossRef] [PubMed]
- Ge, N.; Jia, J.S.; Yang, L.; Huang, R.M.; Wang, Q.Y.; Chen, C.; Meng, Z.G.; Li, L.G.; Chen, J.W. Exogenous gibberellic acid shortening after-ripening process and promoting seed germination in a medicinal plant Panax notoginseng. BMC Plant Biol. 2023, 23, 67. [Google Scholar] [CrossRef] [PubMed]
- Çayan, S.; Sarıkamış, G.; Özmen, C.Y.; Kibar, U.; Özden, E.; Ergül, A. The influence of exogenous gibberellic acid (GA3) and 24-epibrassinolide (24-EpiBL) on seed germination and the expression of genes involved in GA and BR synthesis/signalling in pepper (Capsicum annuum L.). Acta Sci. Pol. Hortorum Cultus 2021, 20, 15–23. [Google Scholar] [CrossRef]
- Müller, F.; Masemola, L.; Britz, E.; Ngcobo, N.; Modiba, S.; Cyster, L.; Samuels, I.; Cupido, C.; Raitt, L. Seed Germination and Early Seedling Growth Responses to Drought Stress in Annual Medicago L. and Trifolium L. Forages. Agronomy 2022, 12, 2960. [Google Scholar] [CrossRef]
- Nie, M.; Ning, N.; Liang, D.; Zhang, H.; Li, S.; Li, S.; Fan, X.; Zhang, Y. Seed priming with selenite enhances germination and seedling growth of Sorghum [Sorghum bicolor (L.) Moench] under salt stress. Acta Agric. Scand. B Soil Plant Sci. 2023, 73, 42–53. [Google Scholar] [CrossRef]
- Yang, N.; Guo, X.; Wu, Y.; Hu, X.; Ma, Y.; Zhang, Y.; Wang, H.; Tang, Z. The inhibited seed germination by ABA and MeJA is associated with the disturbance of reserve utilizations in Astragalus membranaceus. J. Plant Interact. 2018, 13, 388–397. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, H.; Tan, J.; Huang, S.; Chen, X.; Dong, H.; Zhang, R.; Wang, Y.; Wang, B.; Xiao, X.; et al. The chromosome-scale reference genome and transcriptome analysis of Solanum torvum provides insights into resistance to root-knot nematodes. Front. Plant Sci. 2023, 14, 1210513. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Jiao, C.; Singer, S.D.; Gao, M.; Xu, X.; Zhou, Y.; Li, Z.; Fei, Z.; Wang, Y.; Wang, X. Gibberellin-induced changes in the transcriptome of grapevine (Vitis labrusca × V. vinifera) cv. Kyoho flowers. BMC Genom. 2015, 16, 128. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.P. Gibberellin-GID1-DELLA: A pivotal regulatory module for plant growth and development. Plant Physiol. 2010, 154, 567–570. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.M.; Melcher, K.; Teh, B.T.; Xu, H.E. Abscisic acid perception and signaling: Structural mechanisms and applications. Acta Pharmacol. Sin. 2014, 35, 567–584. [Google Scholar] [CrossRef] [PubMed]
- Swarup, R.; Bhosale, R. Developmental Roles of AUX1/LAX Auxin Influx Carriers in Plants. Front. Plant Sci. 2019, 10, 1306. [Google Scholar] [CrossRef] [PubMed]
- Leyser, O. Auxin Signaling. Plant Physiol. 2018, 176, 465–479. [Google Scholar] [CrossRef] [PubMed]
- Matthes, M.S.; Best, N.B.; Robil, J.M.; Malcomber, S.; Gallavotti, A.; McSteen, P. Auxin EvoDevo: Conservation and Diversification of Genes Regulating Auxin Biosynthesis, Transport, and Signaling. Mol. Plant 2019, 12, 298–320. [Google Scholar] [CrossRef] [PubMed]
- Niemeyer, M.; Moreno Castillo, E.; Ihling, C.H.; Iacobucci, C.; Wilde, V.; Hellmuth, A.; Hoehenwarter, W.; Samodelov, S.L.; Zurbriggen, M.D.; Kastritis, P.L.; et al. Flexibility of intrinsically disordered degrons in AUX/IAA proteins reinforces auxin co-receptor assemblies. Nat. Commun. 2020, 11, 2277. [Google Scholar] [CrossRef]
- Salehin, M.; Bagchi, R.; Estelle, M. SCFTIR1/AFB-based auxin perception: Mechanism and role in plant growth and development. Plant Cell 2015, 27, 9–19. [Google Scholar] [CrossRef]
- Wang, R.; Estelle, M. Diversity and specificity: Auxin perception and signaling through the TIR1/AFB pathway. Curr. Opin. Plant Biol. 2014, 21, 51–58. [Google Scholar] [CrossRef]
- Binder, B.M. Ethylene signaling in plants. J. Biol. Chem. 2020, 295, 7710–7725. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.; Nambara, E. Stored and neosynthesized mRNA in Arabidopsis seeds: Effects of cycloheximide and controlled deterioration treatment on the resumption of transcription during imbibition. Plant Mol. Biol. 2010, 73, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Weitbrecht, K.; Muller, K.; Leubner-Metzger, G. First off the mark: Early seed germination. J. Exp. Bot. 2011, 62, 3289–3309. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.; Mukhtar, N.; Bassel, G. Low temperature stimulates spatial molecular reprogramming of the Arabidopsis seed germination programme. Seed Sci. Res. 2020, 30, 2–12. [Google Scholar] [CrossRef]
- Dufkova, H.; Berka, M.; Luklova, M.; Rashotte, A.M.; Brzobohaty, B.; Cerny, M. Eggplant Germination is Promoted by Hydrogen Peroxide and Temperature in an Independent but Overlapping Manner. Molecules 2019, 24, 4270. [Google Scholar] [CrossRef] [PubMed]
- Matus-Cádiz, M.A.; Hucl, P. Rapid and Effective Germination Methods for Overcoming Seed Dormancy in Annual Canarygrass. Crop Sci. 2005, 45, 1696–1703. [Google Scholar] [CrossRef]
- Wang, H.M.; Hou, Y.X.; Wang, S.; Tong, X.H.; Tang, L.Q.; Abolore, A.A.; Zhang, J.; Wang, Y.F. WRKY72 Negatively Regulates Seed Germination Through Interfering Gibberellin Pathway in Rice. Rice Sci. 2021, 28, 1–5. [Google Scholar] [CrossRef]
- Song, Q.; Cheng, S.; Chen, Z.; Nie, G.; Xu, F.; Zhang, J.; Zhou, M.; Zhang, W.; Liao, Y.; Ye, J. Comparative transcriptome analysis revealing the potential mechanism of seed germination stimulated by exogenous gibberellin in Fraxinus hupehensis. BMC Plant Biol. 2019, 19, 199. [Google Scholar] [CrossRef]
- Zhuang, W.; Gao, Z.; Wang, L.; Zhong, W.; Ni, Z.; Zhang, Z. Comparative proteomic and transcriptomic approaches to address the active role of GA4 in Japanese apricot flower bud dormancy release. J. Exp. Bot. 2013, 64, 4953–4966. [Google Scholar] [CrossRef]
- Xia, J.H.; Kermode, A.R. Dormancy of yellow cedar (Chamaecyparis nootkatensis [D. Don] Spach) seed is effectively terminated by treatment with 1-propanol or nitrate in combination with a warm water soaking gibberellin and moist chilling. Seed Sci. Technol. 2000, 28, 227–240. [Google Scholar]
- Ozden, E.; Dermir, I. GA3 enhanced seed germination of Solanum torvum. In Rad Poljoprivredno-Prehrambenog Fak; Univerzitet u Sarajevu: Sarajevo, Bosnia and Herzegovina, 2016; Volume 61, pp. 316–320. [Google Scholar]
- Firenzuoli, A.M.; Vanni, P.; Ramponi, G.; Baccari, V. Changes in Enzyme Levels During Germination of Seeds of Triticum durum. Plant Physiol 1968, 43, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Bykova, N.V.; Igamberdiev, A.U. Cell signaling mechanisms and metabolic regulation of germination and dormancy in barley seeds. Crop J. 2017, 5, 459–477. [Google Scholar] [CrossRef]
- Zhang, Y.; Dai, T.; Liu, Y.; Wang, J.; Wang, Q.; Zhu, W. Effect of Exogenous Glycine Betaine on the Germination of Tomato Seeds under Cold Stress. Int. J. Mol. Sci. 2022, 23, 10474. [Google Scholar] [CrossRef] [PubMed]
- Zaynab, M.; Pan, D.; Noman, A.; Fatima, M.; Abbas, S.; Umair, M.; Sharif, Y.; Chen, S.; Chen, W. Transcriptome approach to address low seed germination in Cyclobalanopsis gilva to save forest ecology. Biochem. Syst. Ecol. 2018, 81, 62–69. [Google Scholar] [CrossRef]
- Duan, C.; Wang, B.; Liu, W.; Chen, J.; Lian, J.; Huan, Z. Effect of chemical and physical factors to improve the germination rate of Echinacea angustifolia seeds. Colloids Surf. B Biointerfaces 2004, 37, 101–105. [Google Scholar] [CrossRef]
- Rentzsch, S.; Podzimska, D.; Voegele, A.; Imbeck, M.; Muller, K.; Linkies, A.; Leubner-Metzger, G. Dose-and tissue-specific interaction of monoterpenes with the gibberellin-mediated release of potato tuber bud dormancy, sprout growth and induction of alpha-amylases and beta-amylases. Planta 2012, 235, 137–151. [Google Scholar] [CrossRef]
- Nietzel, T.; Mostertz, J.; Ruberti, C.; Nee, G.; Fuchs, P.; Wagner, S.; Moseler, A.; Muller-Schussele, S.J.; Benamar, A.; Poschet, G.; et al. Redox-mediated kick-start of mitochondrial energy metabolism drives resource-efficient seed germination. Proc. Natl. Acad. Sci. USA 2020, 117, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Gottschalk, C.; van Nocker, S. Conservation and divergence of expression of GA2-oxidase homeologs in apple (Malus × domestica Borkh.). Front. Plant Sci. 2023, 14, 1117069. [Google Scholar] [CrossRef]
- Chen, S.; Wang, X.; Zhang, L.; Lin, S.; Liu, D.; Wang, Q.; Cai, S.; El-Tanbouly, R.; Gan, L.; Wu, H.; et al. Identification and characterization of tomato gibberellin 2-oxidases (GA2oxs) and effects of fruit-specific SlGA2ox1 overexpression on fruit and seed growth and development. Hortic. Res. 2016, 3, 16059. [Google Scholar] [CrossRef]
- Griffiths, J.; Murase, K.; Rieu, I.; Zentella, R.; Zhang, Z.L.; Powers, S.J.; Gong, F.; Phillips, A.L.; Hedden, P.; Sun, T.P.; et al. Genetic characterization and functional analysis of the GID1 gibberellin receptors in Arabidopsis. Plant Cell 2006, 18, 3399–3414. [Google Scholar] [CrossRef]
- Willige, B.C.; Ghosh, S.; Nill, C.; Zourelidou, M.; Dohmann, E.M.; Maier, A.; Schwechheimer, C. The DELLA domain of GA INSENSITIVE mediates the interaction with the GA INSENSITIVE DWARF1A gibberellin receptor of Arabidopsis. Plant Cell 2007, 19, 1209–1220. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Pan, J.; Wang, H.; Reiter, R.J.; Li, X.; Mou, Z.; Zhang, J.; Yao, Z.; Zhao, D.; Yu, D. Melatonin inhibits seed germination by crosstalk with abscisic acid, gibberellin, and auxin in Arabidopsis. J. Pineal Res. 2021, 70, e12736. [Google Scholar] [CrossRef]
- Chen, Y.; Su, D.; Li, J.; Ying, S.; Deng, H.; He, X.; Zhu, Y.; Li, Y.; Chen, Y.; Pirrello, J.; et al. Overexpression of bHLH95, a basic helix-loop-helix transcription factor family member, impacts trichome formation via regulating gibberellin biosynthesis in tomato. J. Exp. Bot. 2020, 71, 3450–3462. [Google Scholar] [CrossRef]
- Kucera, B.; Cohn, M.A.; Leubner-Metzger, G. Plant hormone interactions during seed dormancy release and germination. Seed Sci. Res. 2005, 15, 281–307. [Google Scholar] [CrossRef]
- Côme, D.; Thevenot, C. Environmental control of embryo dormancy and germination. In The Physiology of Biochemistry of Seed Development, Dormancy and Germination; Khan, A.A., Ed.; Elsevier Biomedical Press: Amsterdam, The Netherlands, 1982; pp. 271–298. [Google Scholar]
- Miransari, M.; Smith, D.L. Plant hormones and seed germination. Environ. Exp. Bot. 2014, 99, 110–121. [Google Scholar] [CrossRef]
- Garciarrubio, A.; Legaria, J.P.; Covarrubias, A.A. Abscisic acid inhibits germination of mature Arabidopsis seeds by limiting the availability of energy and nutrients. Planta 1997, 203, 182–187. [Google Scholar] [CrossRef]
- Gianinetti, A. A Travel through Landscapes of Seed Dormancy. Plants 2023, 12, 3963. [Google Scholar] [CrossRef] [PubMed]
- Dill, A.; Jung, H.S.; Sun, T.P. The DELLA motif is essential for gibberellin-induced degradation of RGA. Proc. Natl. Acad. Sci. USA 2001, 98, 14162–14167. [Google Scholar] [CrossRef]
- Martinez-Andujar, C.; Ordiz, M.I.; Huang, Z.; Nonogaki, M.; Beachy, R.N.; Nonogaki, H. Induction of 9-cis-epoxycarotenoid dioxygenase in Arabidopsis thaliana seeds enhances seed dormancy. Proc. Natl. Acad. Sci. USA 2011, 108, 17225–17229. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, J.; Liu, Y.; Zhao, J.; Fu, J.; Ren, X.; Wang, G.; Wang, J. Exogenous auxin regulates multi-metabolic network and embryo development, controlling seed secondary dormancy and germination in Nicotiana tabacum L. BMC Plant Biol. 2016, 16, 41. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, H.; Zhao, Y.; Feng, Z.; Li, Q.; Yang, H.Q.; Luan, S.; Li, J.; He, Z.H. Auxin controls seed dormancy through stimulation of abscisic acid signaling by inducing ARF-mediated ABI3 activation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2013, 110, 15485–15490. [Google Scholar] [CrossRef] [PubMed]
- Shuai, H.; Meng, Y.; Luo, X.; Chen, F.; Zhou, W.; Dai, Y.; Qi, Y.; Du, J.; Yang, F.; Liu, J.; et al. Exogenous auxin represses soybean seed germination through decreasing the gibberellin/abscisic acid (GA/ABA) ratio. Sci. Rep. 2017, 7, 12620. [Google Scholar] [CrossRef] [PubMed]
- Jensen, P.J.; Hangarter, R.P.; Estelle, M. Auxin transport is required for hypocotyl elongation in light-grown but not dark-grown Arabidopsis. Plant Physiol. 1998, 116, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Carville, J.S.; Spalding, E.P. ABCB19-mediated polar auxin transport modulates Arabidopsis hypocotyl elongation and the endoreplication variant of the cell cycle. Plant J. 2016, 85, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Corbineau, F.; Xia, Q.; Bailly, C.; El-Maarouf-Bouteau, H. Ethylene, a key factor in the regulation of seed dormancy. Front. Plant Sci. 2014, 5, 539. [Google Scholar] [CrossRef] [PubMed]
- Iglesias-Moya, J.; Cebrian, G.; Garrido, D.; Martinez, C.; Jamilena, M. The ethylene receptor mutation etr2b reveals crosstalk between ethylene and ABA in the control of Cucurbita pepo germination. Physiol. Plant 2023, 175, e13864. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, L.; Ye, T.; Zhao, S.; Liu, Z.; Feng, Y.Q.; Wu, Y. Cytokinin antagonizes ABA suppression to seed germination of Arabidopsis by downregulating ABI5 expression. Plant J. 2011, 68, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Greenboim-Wainberg, Y.; Maymon, I.; Borochov, R.; Alvarez, J.; Olszewski, N.; Ori, N.; Eshed, Y.; Weiss, D. Cross talk between gibberellin and cytokinin: The Arabidopsis GA response inhibitor SPINDLY plays a positive role in cytokinin signaling. Plant Cell 2005, 17, 92–102. [Google Scholar] [CrossRef]
- Shu, K.; Liu, X.D.; Xie, Q.; He, Z.H. Two Faces of One Seed: Hormonal Regulation of Dormancy and Germination. Mol. Plant 2016, 9, 34–45. [Google Scholar] [CrossRef]
- Varshney, V.; Majee, M. JA Shakes Hands with ABA to Delay Seed Germination. Trends Plant Sci. 2021, 26, 764–766. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yu, F.; Xie, Q. Balancing growth and adaptation to stress: Crosstalk between brassinosteroid and abscisic acid signaling. Plant Cell Environ. 2020, 43, 2325–2335. [Google Scholar] [CrossRef] [PubMed]
- Mei, S.; Zhang, M.; Ye, J.; Du, J.; Jiang, Y.; Hu, Y. Auxin contributes to jasmonate-mediated regulation of abscisic acid signaling during seed germination in Arabidopsis. Plant Cell 2023, 35, 1110–1133. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, R.R.; Lynch, T.J. The Arabidopsis abscisic acid response gene ABI5 encodes a basic leucine zipper transcription factor. Plant Cell 2000, 12, 599–609. [Google Scholar] [CrossRef]
- Lopez-Molina, L.; Chua, N.H. A null mutation in a bZIP factor confers ABA-insensitivity in Arabidopsis thaliana. Plant Cell Physiol. 2000, 41, 541–547. [Google Scholar] [CrossRef]
- Lopez-Molina, L.; Mongrand, S.; Chua, N.H. A postgermination developmental arrest checkpoint is mediated by abscisic acid and requires the ABI5 transcription factor in Arabidopsis. Proc. Natl. Acad. Sci. USA 2001, 98, 4782–4787. [Google Scholar] [CrossRef]
- Pan, J.; Hu, Y.; Wang, H.; Guo, Q.; Chen, Y.; Howe, G.A.; Yu, D. Molecular Mechanism Underlying the Synergetic Effect of Jasmonate on Abscisic Acid Signaling during Seed Germination in Arabidopsis. Plant Cell 2020, 32, 3846–3865. [Google Scholar] [CrossRef]
- Li, Q.; Chen, X.; Zhang, S.; Shan, S.; Xiang, Y. DELAY OF GERMINATION 1, the Master Regulator of Seed Dormancy, Integrates the Regulatory Network of Phytohormones at the Transcriptional Level to Control Seed Dormancy. Curr. Issues Mol. Biol. 2022, 44, 6205–6217. [Google Scholar] [CrossRef]
- Nakabayashi, K.; Bartsch, M.; Xiang, Y.; Miatton, E.; Pellengahr, S.; Yano, R.; Seo, M.; Soppe, W.J. The time required for dormancy release in Arabidopsis is determined by DELAY OF GERMINATION1 protein levels in freshly harvested seeds. Plant Cell 2012, 24, 2826–2838. [Google Scholar] [CrossRef]
- Huo, H.; Wei, S.; Bradford, K.J. DELAY OF GERMINATION1 (DOG1) regulates both seed dormancy and flowering time through microRNA pathways. Proc. Natl. Acad. Sci. USA 2016, 113, E2199–E2206. [Google Scholar] [CrossRef]
- Dekkers, B.J.; He, H.; Hanson, J.; Willems, L.A.; Jamar, D.C.; Cueff, G.; Rajjou, L.; Hilhorst, H.W.; Bentsink, L. The Arabidopsis DELAY OF GERMINATION 1 gene affects ABSCISIC ACID INSENSITIVE 5 (ABI5) expression and genetically interacts with ABI3 during Arabidopsis seed development. Plant J. 2016, 85, 451–465. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.; Zhou, Y.; Yao, L.; Yu, L.; Qiao, Z.; Tang, M.; Wei, F. Amomum tsaoko DRM1 regulate seed germination and improve heat tolerance in Arabidopsis. J. Plant Physiol. 2023, 286, 154007. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, J.M.; Kwong, R.W.; Park, S.; Le, B.H.; Baden, R.; Cagliari, A.; Hashimoto, M.; Munoz, M.D.; Fischer, R.L.; Goldberg, R.B.; et al. LEC1 sequentially regulates the transcription of genes involved in diverse developmental processes during seed development. Proc. Natl. Acad. Sci. USA 2017, 114, E6710–E6719. [Google Scholar] [CrossRef] [PubMed]
- Rikiishi, K.; Maekawa, M. Seed maturation regulators are related to the control of seed dormancy in wheat (Triticum aestivum L.). PLoS ONE 2014, 9, e107618. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, A.; Kagaya, Y.; Usui, H.; Hobo, T.; Takeda, S.; Hattori, T. Diverse roles and mechanisms of gene regulation by the Arabidopsis seed maturation master regulator FUS3 revealed by microarray analysis. Plant Cell Physiol. 2010, 51, 2031–2046. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Lee, N.; Kim, W.; Lim, S.; Choi, G. ABI3 and PIL5 collaboratively activate the expression of SOMNUS by directly binding to its promoter in imbibed Arabidopsis seeds. Plant Cell 2011, 23, 1404–1415. [Google Scholar] [CrossRef]
- Yang, L.; Jiang, Z.; Jing, Y.; Lin, R. PIF1 and RVE1 form a transcriptional feedback loop to control light-mediated seed germination in Arabidopsis. J. Integr. Plant Biol. 2020, 62, 1372–1384. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).