

Detection of Quantitative Trait Loci Associated with Alkaline Tolerance Using Recombinant Inbred Line Population Derived from Longdao5 × Zhongyouzao8 at Seedling Stage


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Linkage Map Construction
2.3. Alkaline-Tolerance Evaluation
2.4. QTL Mapping for Alkali Tolerance
3. Results
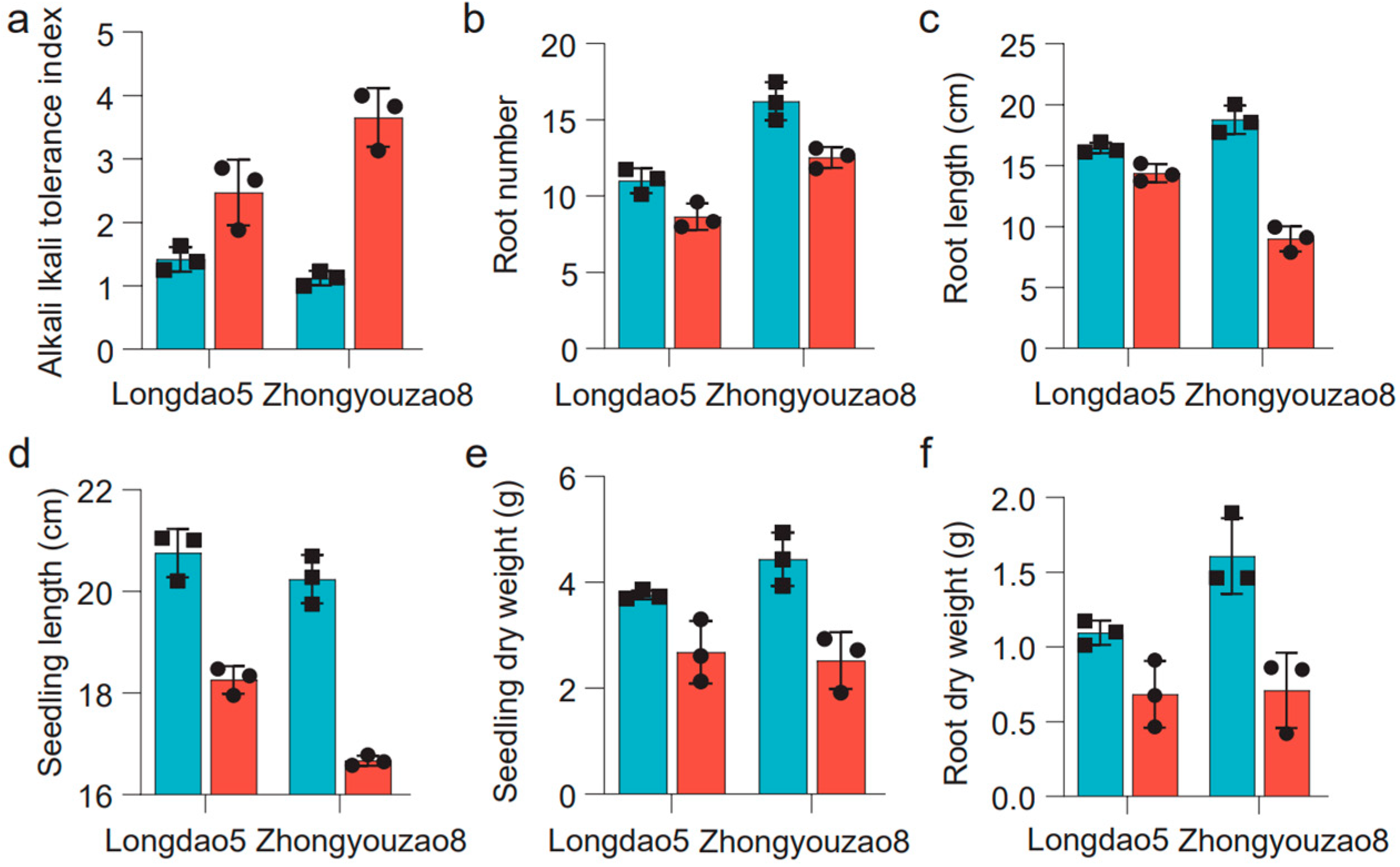
3.1. Seedling Performance between Longdao5 and Zhongyouzao8 under Alkaline Stress
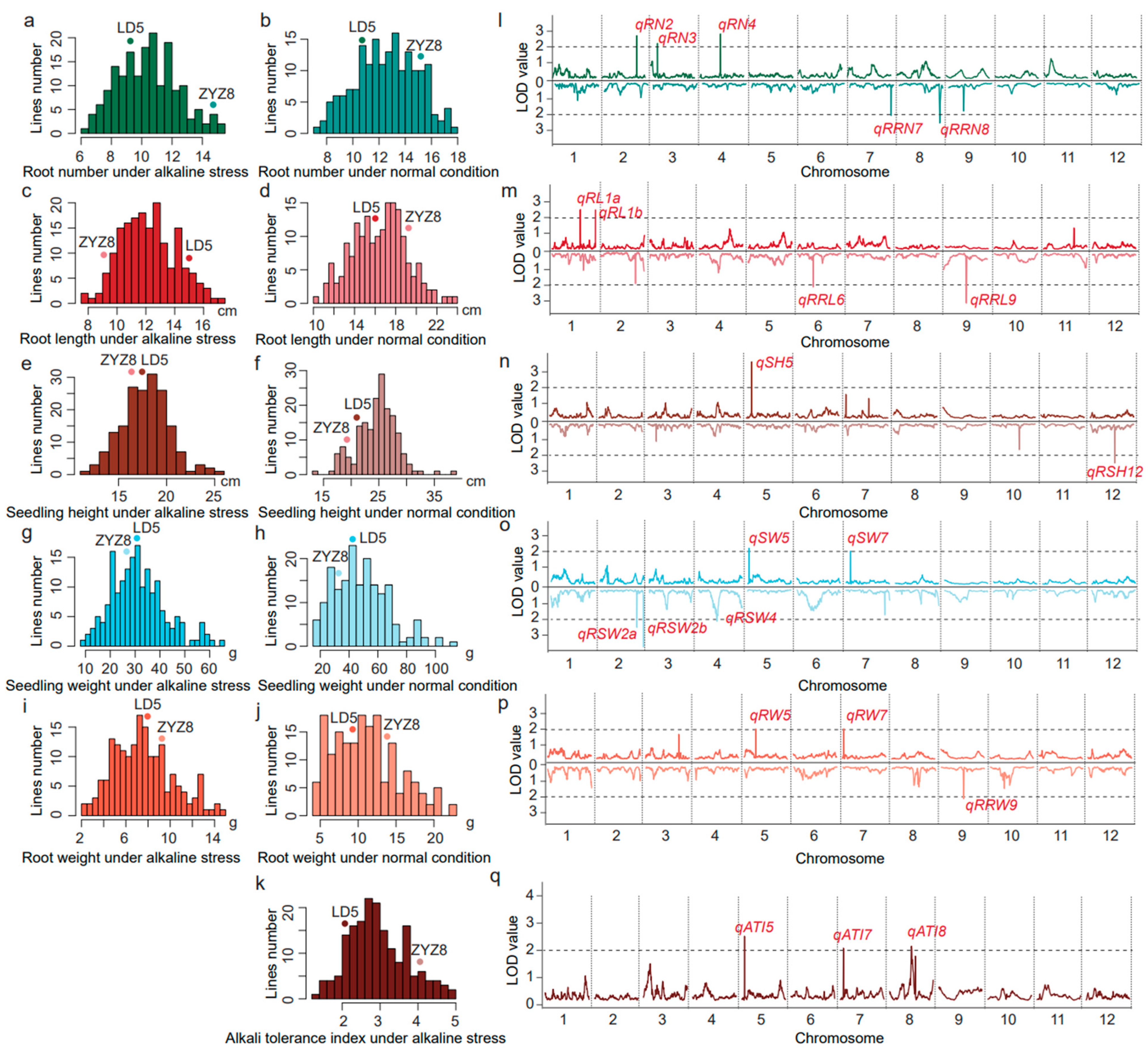
3.2. Phenotypic Variation in the RIL Population under Alkaline Stress
3.3. The Correlation Coefficients between the ATI and the Seedling Traits in the RIL Population
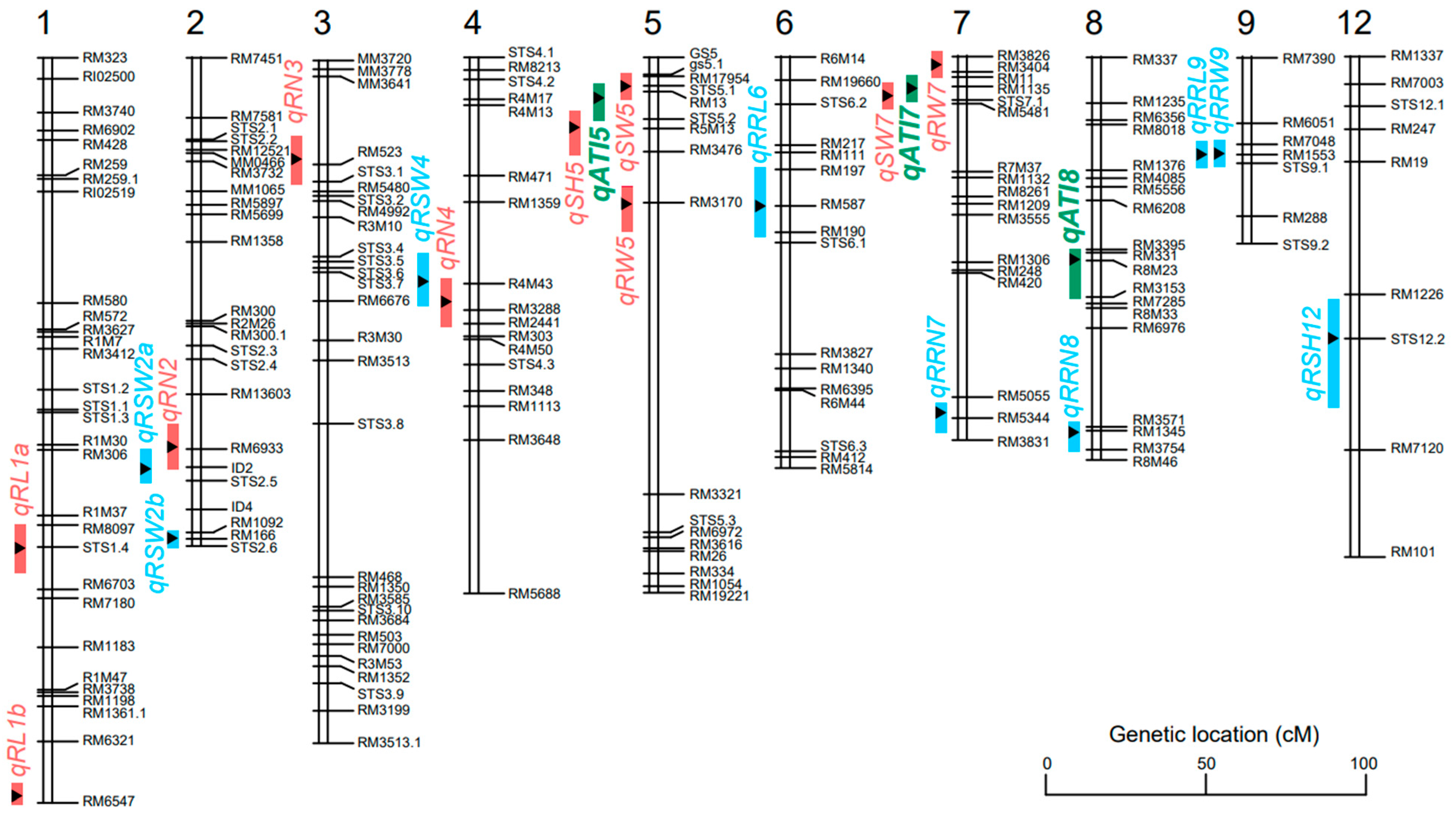
3.4. QTL Mapping for Seedling- and Root-Related Traits under Alkali Stresses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, J.; Wan, J.; Yin, X.; Gu, X.; Yin, H.; Yu, M.; Luan, Y.; Li, R.; Mi, T.; Li, J. Progress and prospect of developing salt and alkali tolerant rice using hybrid rice technology in China. Plant Breed. 2024, 143, 86–95. [Google Scholar] [CrossRef]
- Tsai, H.; Schmidt, W. The enigma of environmental pH sensing in plants. Nat. Plants 2021, 7, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Song, H.; Zhang, L. New Insight into Plant Saline-Alkali Tolerance Mechanisms and Application to Breeding. Int. J. Mol. Sci. 2022, 23, 16048. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yu, F.; Xie, P.; Sun, S.; Qiao, X.; Tang, S.; Chen, C.; Yang, S.; Mei, C.; Yang, D.; et al. A Gγ protein regulates alkaline sensitivity in crops. Science 2023, 379, eade8416. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Zhang, H.; Yang, S.; Liu, L.; Xie, P.; Li, J.; Zhu, Y.; Ouyang, Y.; Xie, Q.; Zhang, H.; et al. Genetic modification of Gγ subunit AT1 enhances salt-alkali tolerance in main graminaceous crops. Natl. Sci. Rev. 2023, 10, nwad075. [Google Scholar] [CrossRef] [PubMed]
- Jamil, A.; Riaz, S.; Ashraf, M.; Foolad, M. Gene Expression Profiling of Plants under Salt Stress. Crit. Rev. Plant Sci. 2011, 30, 435–458. [Google Scholar] [CrossRef]
- Li, B.; Wang, Z.; Sun, Z.; Chen, Y.; Yang, F. Resources and sustainable resource exploitation of salinized land in China. Agric. Res. Arid. Areas 2005, 23, 154–158. [Google Scholar]
- Liang, Z.; Wang, Z.; Ma, H.; Yang, F.; Chen, Y.; Huang, L.; Kong, X.; Yan, C.; Liu, M.; Wang, M.; et al. The Progress in Improvement of High pH Saline-Alkali Soil in the Songnen Plain by Stress Tolerant Plants. J. Jilin Agric. Univ. 2008, 30, 517–528. [Google Scholar]
- Zhao, L.; Feng, J.; Wang, Y.; Wang, H.; Geng, Y.; Wang, S.; Li, C. Theoretical and Technological Problems in the Development of Planting Paddy in Saline-alkali Land of Songnen Plain. J. Jilin Agric. Univ. 2012, 34, 237–241. [Google Scholar]
- Zhang, L.; Qian, Q.; Song, S. Combating alkaline stress with Alkaline tolerance 1, which encodes a conserved Gγ protein in multiple crops. Plant Commun. 2023, 4, 100603. [Google Scholar] [CrossRef]
- Li, Q.; Yang, A.; Zhang, W. Efficient acquisition of iron confers greater tolerance to saline-alkaline stress in rice (Oryza sativa L.). J. Exp. Bot. 2016, 67, 6431–6444. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Meng, W.; Wang, Y.; Zhou, Y.; Wang, S.; Qi, F.; Wang, N.; Ma, J. Comparative Analysis of Physiological, Hormonal and Transcriptomic Responses Reveal Mechanisms of Saline-Alkali Tolerance in Autotetraploid Rice (Oryza sativa L.). Int. J. Mol. Sci. 2022, 23, 16146. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Takano, T.; Liu, S. Screening and Evaluation of Saline–Alkaline Tolerant Germplasm of Rice (Oryza sativa L.) in Soda Saline–Alkali Soil. Agronomy 2018, 8, 205. [Google Scholar] [CrossRef]
- Qi, D.; Guo, G.; Lee, M.; Zhang, J.; Cao, G.; Zhang, S.; Suh, S.; Zhou, Q.; Han, L. Identification of quantitative trait loci for the dead leaf rate and the seedling dead rate under alkaline stress in rice. J. Genet. Genom. 2008, 35, 299–305. [Google Scholar] [CrossRef]
- Qi, D.; Guo, G.; Lee, M.; Yang, C.; Zhang, J.; Cao, G.; Zhang, S.; Suh, S.; Zhou, Q.; Han, L. Identification of Quantitative Trait Loci for Alkaline Tolerance at Early Seedling Stage of Japonica Rice Under Alkaline Stress. Acta Agron. Sin. 2009, 35, 301–308. [Google Scholar]
- Qi, D.; Li, D.; Yang, C.; Li, M.; Cao, G.; Zhang, J.; Zhou, Q.; Xu, X.; Zhang, S.; Han, L. Detection of QTL for Alkali Tolerance at the Germination Stage in japonica Rice. Chin. J. Rice Sci. 2009, 23, 589–594. [Google Scholar]
- Li, N.; Sun, J.; Wang, J.; Liu, H.; Zheng, H.; Yang, L.; Liang, Y.; Li, X.; Zou, D. QTL analysis for alkaline tolerance of rice and verification of a major QTL. Plant Breed. 2017, 136, 881–891. [Google Scholar] [CrossRef]
- Li, N.; Zheng, H.; Cui, J.; Wang, J.; Liu, H.; Sun, J.; Liu, T.; Zhao, H.; Lai, Y.; Zou, D. Genome-wide association study and candidate gene analysis of alkalinity tolerance in japonica rice germplasm at the seedling stage. Rice 2019, 12, 24. [Google Scholar] [CrossRef]
- Li, X.; Zheng, H.; Wu, W.; Liu, H.; Wang, J.; Jia, Y.; Li, J.; Yang, L.; Lei, L.; Zou, D.; et al. QTL Mapping and Candidate Gene Analysis for Alkali Tolerance in Japonica Rice at the bud Stage Based on Linkage Mapping and Genome-Wide Association Study. Rice 2020, 13, 48. [Google Scholar] [CrossRef]
- Singh, L.; Coronejo, S.; Pruthi, R.; Chapagain, S.; Subudhi, P. Integration of QTL Mapping and Whole Genome Sequencing Identifies Candidate Genes for Alkalinity Tolerance in Rice (Oryza sativa). Int. J. Mol. Sci. 2022, 23, 11791. [Google Scholar] [CrossRef]
- Mei, S.; Zhang, G.; Jiang, J.; Lu, J.; Zhang, F. Combining Genome-Wide Association Study and Gene-Based Haplotype Analysis to Identify Candidate Genes for Alkali Tolerance at the Germination Stage in Rice. Front. Plant Sci. 2022, 13, 887239. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Wang, R.; Wang, J.; Hua, K.; Wang, Y.; Liu, X.; Yao, S. ALT1, a Snf2 family chromatin remodeling ATPase, negatively regulates alkaline tolerance through enhanced defense against oxidative stress in rice. PLoS ONE 2014, 9, e112515. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.; Kim, Y.; Shin, R.; Park, C. Nucleus-Encoded Thylakoid Protein, OsY3IP1, Confers Enhanced Tolerance to Saline and Alkaline Stresses in Rice. Rice Sci. 2022, 29, 225–236. [Google Scholar] [CrossRef]
- Shen, T.; Xu, F.; Chen, D.; Yan, R.; Wang, Q.; Li, K.; Zhang, G.; Ni, L.; Jiang, M. A B-box transcription factor OsBBX17 regulates saline-alkaline tolerance through the MAPK cascade pathway in rice. New Phytol. 2024 241, 2158–2175. [CrossRef]
- Zhu, F.; Wang, K.; Li, D.; Liu, Z.; Li, M.; Wang, Z.; Li, X.; Lan, X.; Guan, Q. OsSAP6 Positively Regulates Soda Saline-Alkaline Stress Tolerance in Rice. Rice 2022, 15, 69. [Google Scholar] [CrossRef]
- Guan, Q.; Ma, H.; Wang, Z.; Wang, Z.; Bu, Q.; Liu, S. A rice LSD1-like-type ZFP gene OsLOL5 enhances saline-alkaline tolerance in transgenic Arabidopsis thaliana, yeast and rice. BMC Genom. 2016, 17, 142. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, X.; Xue, S.; Quan, T.; Cui, D.; Han, L.; Cong, W.; Li, M.; Yun, D.; Liu, B.; et al. SET DOMAIN GROUP 721 protein functions in saline-alkaline stress tolerance in the model rice variety Kitaake. Plant Biotechnol. J. 2021, 19, 2576–2588. [Google Scholar] [CrossRef]
- Ni, L.; Wang, S.; Shen, T.; Wang, Q.; Chen, C.; Xia, J.; Jiang, M. Calcium/calmodulin-dependent protein kinase OsDMI3 positively regulates saline-alkaline tolerance in rice roots. Plant Signal Behav. 2020, 15, 1813999. [Google Scholar] [CrossRef]
- Ganie, S.; Molla, K.; Henry, R.; Bhat, K.; Mondal, T. Advances in understanding salt tolerance in rice. Theor. Appl. Genet. 2019, 132, 851–870. [Google Scholar] [CrossRef]
- Liu, C.; Mao, B.; Yuan, D.; Chu, C.; Duan, M. Salt tolerance in rice: Physiological responses and molecular mechanisms. Crop J. 2022, 10, 13–25. [Google Scholar] [CrossRef]
- Meng, L.; Li, H.; Zhang, L.; Wang, J. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J. 2015, 3, 269–283. [Google Scholar] [CrossRef]
- Arends, D.; Prins, P.; Jansen, R.; Broman, K. R/qtl: High-throughput multiple QTL mapping. Bioinformatics 2010, 26, 2990–2992. [Google Scholar] [CrossRef] [PubMed]
- Hou, B.; Yang, C.; Zhang, X.; Yang, X.; Wang, L.; Wang, J.; Li, H.; Jiang, S. Mapping of Grain Shape QTLs Using RIL Population from Longdao 5/Zhongyouzao 8. Chin. J. Rice Sci. 2024, 38, 13–24. [Google Scholar]
- Zhang, C.; Jin, F.; Li, S.; Liu, W.; Ma, X.; Yang, S.; Yang, D.; Li, X. Fine mapping of major QTLs for alkaline tolerance at the seedling stage in maize (Zea mays L.) through genetic linkage analysis combined with high-throughput DNA sequencing. Euphytica 2018, 214, 120. [Google Scholar] [CrossRef]
- Xiao, J.; Li, J.; Grandillo, S.; Ahn, S.; Yuan, L.; Tanksley, S.; McCouch, S. Identification of trait-improving quantitative trait loci alleles from a wild rice relative, Oryza rufipogon. Genetics 1998, 150, 899–909. [Google Scholar] [CrossRef]
- Han, L.; Qiao, Y.; Zhang, Y.; An, Y.; Ye, J.; Koh, H. QTL analysis on cold tolerance during early growth period in rice. Chin. J. Rice Sci. 2005, 19, 122–126. [Google Scholar]
- Zhang, F.; Hao, X.; Gao, Y.; Hua, Z.; Ma, X.; Chen, W.; Xu, Z.; Zhu, L.; Li, Z. Improving seedling cold tolerance of japonica rice by using the "hidden diversity" in indica rice germplasm in a backcross breeding program. Acta Agron. Sin. 2007, 33, 1618–1624. [Google Scholar]
- Andaya, V.; Mackill, D. Mapping of QTLs associated with cold tolerance during the vegetative stage in rice. J. Exp. Bot. 2003, 54, 2579–2585. [Google Scholar] [CrossRef]
- Mickelbart, M.; Hasegawa, P.; Bailey-Serres, J. Genetic mechanisms of abiotic stress tolerance that translate to crop yield stability. Nat. Rev. Genet. 2015, 16, 237–251. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Parents | RIL Population | |||||
|---|---|---|---|---|---|---|---|
| Longdao5 | Zhongyouzao8 | Mean ± SD | Range | CV (%) | Skewness | Kurtosis | |
| RNAT | 2.68 ± 0.59 | 2.52 ± 0.53 | 10.43 ± 1.96 | 6.00–15.19 | 18.81 | 0.22 | −0.41 |
| RN | 3.76 ± 0.09 | 4.34 ± 0.51 | 16.58 ± 10.67 | 0.08–46.83 | 64.34 | 0.40 | −0.57 |
| RLAT | 14.38 ± 0.75 | 8.99 ± 1.03 | 12.20 ± 1.90 | 7.83–17.08 | 15.55 | 0.21 | −0.51 |
| RL | 16.45 ± 0.44 | 18.77 ± 1.17 | 24.62 ± 13.11 | 0.27–52.91 | 53.24 | −0.07 | −0.97 |
| SHAT | 18.26 ± 0.27 | 16.67 ± 0.10 | 17.82 ± 2.45 | 11.19–25.41 | 13.74 | 0.18 | 0.31 |
| SH | 20.76 ± 0.47 | 20.24 ± 0.48 | 27.17 ± 7.07 | 4.96–47.55 | 26.03 | −0.12 | 0.36 |
| SWAT | 2.68 ± 0.59 | 2.52 ± 0.53 | 31.39 ± 10.91 | 9.67–64.88 | 34.76 | 0.62 | 0.26 |
| SW | 3.76 ± 0.09 | 4.44 ± 0.51 | 32.24 ± 16.23 | 2.41–77.31 | 50.36 | 0.32 | −0.34 |
| RWAT | 0.69 ± 0.22 | 0.71 ± 0.25 | 7.62 ± 2.68 | 2.44–14.77 | 35.19 | 0.47 | −0.25 |
| RW | 1.10 ± 0.08 | 1.61 ± 0.25 | 28.03 ± 18.48 | 0.62–76.21 | 65.92 | 0.27 | −0.91 |
| ATI | 2.47 ± 0.52 | 3.65 ± 0.46 | 2.96 ± 0.76 | 1.36–5.00 | 25.82 | 0.40 | −0.27 |
| QTL | Trait | Chr. a | Peak Marker | QTL Interval | LOD b | Var (%) c | Add. d | Positive Allele |
|---|---|---|---|---|---|---|---|---|
| qATI5 | Alkali tolerance index | 5 | RM13 | STS5.1-STS5.2 | 2.60 | 7.98 | −0.15 | LD5 |
| qATI7 | Alkali tolerance index | 7 | RM1135 | RM11-STS7.1 | 2.11 | 6.53 | 0.12 | ZYZ8 |
| qATI8 | Alkali tolerance index | 8 | R8M23 | RM331-RM3153 | 2.19 | 6.76 | −0.06 | LD5 |
| qRN2 | Root number | 2 | RM6933 | RM13603-ID2 | 2.71 | 8.30 | 0.43 | ZYZ8 |
| qRN3 | Root number | 3 | RM523 | MM3641-STS3.1 | 2.17 | 6.70 | −0.31 | LD5 |
| qRN4 | Root number | 4 | RM3288 | R4M43-RM2441 | 2.82 | 8.62 | 0.44 | ZYZ8 |
| qSH5 | Seedling height | 5 | R5M13 | STS5.2-RM3476 | 3.55 | 10.73 | −0.68 | LD5 |
| qRL1a | Root length | 1 | STS1.4 | RM8097-RM6703 | 2.51 | 7.71 | −0.48 | LD5 |
| qRL1b | Root length | 1 | RM6547 | RM6321-RM6547 | 2.52 | 7.74 | −0.42 | LD5 |
| qSW5 | Seedling dry weight | 5 | RM17954 | gs5.1-RM13 | 2.29 | 7.06 | −2.74 | LD5 |
| qSW7 | Seedling dry weight | 7 | STS7.1 | RM1135-RM5481 | 2.01 | 6.23 | 2.07 | ZYZ8 |
| qRW5 | Root dry weight | 5 | RM3170 | RM3476-RM3321 | 2.01 | 6.23 | −0.54 | LD5 |
| qRW7 | Root dry weight | 7 | RM3404 | RM3826-RM11 | 2.04 | 6.32 | 0.61 | ZYZ8 |
| QTL | Trait | Chr. a | Peak Marker | QTL Interval | LOD b | Var (%) c | Add. d | Positive Allele |
|---|---|---|---|---|---|---|---|---|
| qRRN7 | Relative root number | 7 | RM5344 | RM5055-RM3831 | 2.04 | 6.32 | −1.95 | LD5 |
| qRRN8 | Relative root number | 8 | RM1345 | RM3571-RM3754 | 2.54 | 7.80 | 0.04 | ZYZ8 |
| qRSH12 | Relative seedling height | 12 | STS12.2 | RM1226-RM7120 | 2.49 | 7.65 | −1.52 | LD5 |
| qRRL6 | Relative root length | 6 | RM587 | RM197-RM190 | 2.13 | 6.58 | 3.57 | ZYZ8 |
| qRRL9 | Relative root length | 9 | RM1553 | RM7048-STS9.1 | 3.17 | 9.64 | −3.32 | LD5 |
| qRSW2a | Relative seedling dry weight | 2 | ID2 | RM6933-STS2.5 | 2.39 | 7.36 | 2.12 | ZYZ8 |
| qRSW2b | Relative seedling dry weight | 2 | RM166 | RM1092-STS2.6 | 3.63 | 10.96 | −3.47 | LD5 |
| qRSW4 | Relative seedling dry weight | 4 | R4M43 | RM1359-RM3288 | 2.01 | 6.23 | 3.16 | ZYZ8 |
| qRRW9 | Relative root dry weight | 9 | RM1553 | RM7048-STS9.1 | 2.03 | 6.29 | −3.57 | LD5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Liu, K.; Yang, C.; Hou, B.; Yang, X.; Wang, L.; Cui, S.; Lai, Y.; Li, Z.; Jiang, S. Detection of Quantitative Trait Loci Associated with Alkaline Tolerance Using Recombinant Inbred Line Population Derived from Longdao5 × Zhongyouzao8 at Seedling Stage. Life 2024, 14, 1151. https://doi.org/10.3390/life14091151
Zhang X, Liu K, Yang C, Hou B, Yang X, Wang L, Cui S, Lai Y, Li Z, Jiang S. Detection of Quantitative Trait Loci Associated with Alkaline Tolerance Using Recombinant Inbred Line Population Derived from Longdao5 × Zhongyouzao8 at Seedling Stage. Life. 2024; 14(9):1151. https://doi.org/10.3390/life14091151
Chicago/Turabian StyleZhang, Xijuan, Kai Liu, Chuanming Yang, Benfu Hou, Xianli Yang, Lizhi Wang, Shize Cui, Yongcai Lai, Zhugang Li, and Shukun Jiang. 2024. "Detection of Quantitative Trait Loci Associated with Alkaline Tolerance Using Recombinant Inbred Line Population Derived from Longdao5 × Zhongyouzao8 at Seedling Stage" Life 14, no. 9: 1151. https://doi.org/10.3390/life14091151
APA StyleZhang, X., Liu, K., Yang, C., Hou, B., Yang, X., Wang, L., Cui, S., Lai, Y., Li, Z., & Jiang, S. (2024). Detection of Quantitative Trait Loci Associated with Alkaline Tolerance Using Recombinant Inbred Line Population Derived from Longdao5 × Zhongyouzao8 at Seedling Stage. Life, 14(9), 1151. https://doi.org/10.3390/life14091151