
Storing up Treasures: Storage Potential of Macrolophus pygmaeus (Hemiptera: Heteroptera: Miridae) Nymphs for Application in Biological Control



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and General Methods
2.2. The First Experiment: The Effect of Storage Conditions on Nymph Longevity
- The temperature conditions during the pre-storage development (20 or 24 °C);
- The photoperiodic conditions during the pre-storage development (L:D = 10:14 or 16:8 h);
- The food availability during the storage (starved or fed with the grain moth eggs);
- The temperature conditions during the storage (3, 6, 9, or 12 °C).
2.3. The Second Experiment: The Effect of Storage Conditions on Nymphal Survival and Female Fecundity
- The storage conditions (3 °C without feeding or 9 °C with feeding);
- The period of storage (0, 10, 20, or 30 days at 3 °C and 0, 30, 60, 90, or 120 days at 9 °C).
- The nymphal survival (i.e., the proportion of nymphs that reached the adult stage);
- The fecundity (i.e., the number of eggs laid by a female during the first 10 days of her adult life).
2.4. Statstical Analysis
3. Results
3.1. The First Experiment: Effect of Storage Conditions on Nymph Longevity
3.2. The Second Experiment: Effect of Storage Conditions on Nymphal Survival and Female Fecundity
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ballal, C.R.; Yamada, K. Anthocorid predators. In Ecofriendly Pest Management for Food Security; Omkar, Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 183–216. [Google Scholar] [CrossRef]
- Colinet, H.; Boivin, G. Insect parasitoids cold storage: A comprehensive review of factors of variability and consequences. Biol. Contr. 2011, 58, 83–95. [Google Scholar] [CrossRef]
- van Lenteren, J.C.; Alomar, O.; Ravensberg, W.J.; Urbaneja, A. Biological control agents for control of pests in greenhouses. In Integrated Pest and Disease Management in Greenhouse Crops; Gullino, M.L., Albajes, R., Nicot, P.C., Eds.; Springer International Publishing: New York, NY, USA, 2020; pp. 409–439. [Google Scholar]
- van Lenteren, J.C. IOBC Internet Book of Biological Control, Version 6. 2012. Available online: https://www.iobc-global.org/publications_iobc_internet_book_of_biological_control.html (accessed on 10 May 2024).
- van Lenteren, J.C.; Tommasini, M.G. Mass production, storage, shipment and release of natural enemies. In Quality Control and Production of Biological Control Agents: Theory and Testing Procedures; van Lenteren, J.C., Ed.; CABI Publishing: Wallingford, UK, 2003; pp. 181–189. [Google Scholar]
- Danks, H.V. The elements of seasonal adaptations in insects. Can. Entomol. 2007, 139, 1–44. [Google Scholar] [CrossRef]
- Denlinger, D.L. Insect Diapause; Cambridge University Press: Cambridge, UK, 2022. [Google Scholar]
- Tauber, M.J.; Tauber, C.A.; Masaki, S. Seasonal Adaptations of Insects; Oxford University Press: New York, NY, USA, 1986. [Google Scholar]
- Denlinger, D.L. Dormancy in tropical insects. Ann. Rev. Entomol. 1986, 31, 239–264. [Google Scholar] [CrossRef]
- van den Meiracker, R.A.F. Induction and termination of diapause in Orius predatory bugs. Entomol. Exp. Appl. 1994, 73, 127–137. [Google Scholar] [CrossRef]
- Musolin, D.L.; Ito, K. Photoperiodic and temperature control of nymphal development and induction of reproductive diapause in two predatory Orius bugs: Interspecific and geographic differences. Physiol. Entomol. 2008, 33, 291–301. [Google Scholar] [CrossRef]
- Kobayashi, T.; Osakabe, M. Pre-winter copulation enhances overwintering success of Orius females (Heteroptera: Anthocoridae). Appl. Entomol. Zool. 2009, 44, 47–52. [Google Scholar] [CrossRef]
- Arnó, J.; Castañé, C.; Alomar, O.; Riudavets, J.; Agustí, N.; Gabarra, R.; Albajes, R. Forty years of biological control in Mediterranean tomato greenhouses: The story of success. Isr. J. Entomol. 2018, 48, 209–226. [Google Scholar] [CrossRef]
- Dutra, T.M.; da Graça Batista, M.; Teixeira, J.C.; Todorova, S.; Oliveira, L.; Tavares, J.; Borges, I.; Soares, A.O. Economic and financial model to the mass-rearing of Macrolophus pygmaeus (Rambur) (Heteroptera: Miridae), a biological control agent against the tomato moth Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) in protected culture. Pest Manag. Sci. 2023, 79, 3712–3720. [Google Scholar] [CrossRef]
- Gigon, V.; Camps, C.; Le Corff, J. Biological control of Tetranychus urticae by Phytoseiulus macropilis and Macrolophus pygmaeus in tomato greenhouses. Exp. Appl. Acarol. 2016, 68, 55–70. [Google Scholar] [CrossRef]
- Moerkens, R.; Berckmoes, E.; Van Damme, V.; Wittemans, L.; Tirry, L.; Casteels, H.; De Clercq, P.; De Vis, R. Inoculative release strategies of Macrolophus pygmaeus Rambur (Hemiptera: Miridae) in tomato crops: Population dynamics and dispersal. J. Plant Dis. Prot. 2017, 124, 295–303. [Google Scholar] [CrossRef]
- Sanchez, J.A.; Lopez-Gallego, E.; Pérez-Marcos, M.; Perera-Fernandez, L.G.; Ramirez-Soria, M.J. How safe is it to rely on Macrolophus pygmaeus (Hemiptera: Miridae) as a biocontrol agent in tomato crops? Front. Ecol. Evol. 2018, 6, 132. [Google Scholar] [CrossRef]
- Urbaneja, A.; Montón, H.; Mollá, O. Suitability of the tomato borer Tuta absoluta as prey for Macrolophus pygmaeus and Nesidiocoris tenuis. J. Appl. Entomol. 2009, 133, 292–296. [Google Scholar] [CrossRef]
- Fantinou, A.A.; Perdikis, D.C.; Labropoulos, P.D.; Maselou, D.A. Preference and consumption of Macrolophus pygmaeus preying on mixed instar assemblages of Myzus persicae. Biol. Contr. 2009, 51, 76–80. [Google Scholar] [CrossRef]
- Ingegno, B.L.; Pansa, M.G.; Tavella, L. Plant preference in the zoophytophagous generalist predator Macrolophus pygmaeus (Heteroptera: Miridae). Biol. Contr. 2011, 58, 174–181. [Google Scholar] [CrossRef]
- Martínez-García, H.; Sáenz-Romo, M.G.; Aragón-Sánchez, M.; Román-Fernández, L.R.; Sáenz-de-Cabezón, E.; Marco-Mancebón, V.S.; Pérez-Moreno, I. Temperature-dependent development of Macrolophus pygmaeus and its applicability to biological control. BioControl 2017, 62, 481–493. [Google Scholar] [CrossRef]
- Nissinen, A.I.; Pinto-Zevallos, D.M.; Jauhiainen, L.; Vänninen, I. The effect of photoperiod and light quality on Macrolophus pygmaeus Rambur (Hemiptera: Miridae) nymphal development, fecundity and longevity. Biol. Contr. 2017, 108, 30–39. [Google Scholar] [CrossRef]
- Pazyuk, I.M.; Reznik, S.Y. Influence of photoperiod on development and maturation of Macrolophus pygmaeus (Hemiptera, Miridae). Entomol. Rev. 2016, 96, 274–279. [Google Scholar] [CrossRef]
- Perdikis, D.; Lykouressis, D. Effects of various items, host plants, and temperatures on the development and survival of Macrolophus pygmaeus Rambur (Hemiptera: Miridae). Biol. Contr. 2000, 17, 55–60. [Google Scholar] [CrossRef]
- Sylla, S.; Brévault, T.; Diarra, K.; Bearez, P.; Desneux, N. Life-history traits of Macrolophus pygmaeus with different prey foods. PLoS ONE 2016, 11, e0166610. [Google Scholar] [CrossRef]
- Vandekerkhove, B.; De Clercq, P. Pollen as an alternative or supplementary food for the mirid predator Macrolophus pygmaeus. Biol. Contr. 2010, 53, 238–242. [Google Scholar] [CrossRef]
- Vandekerkhove, B.; De Puysseleyr, V.; Bonte, M.; De Clercq, P. Fitness and predation potential of Macrolophus pygmaeus reared under artificial conditions. Insect Sci. 2011, 18, 682–688. [Google Scholar] [CrossRef]
- Puchkov, V.G. Plant bugs (Heteroptera, Miridae) of Poltava region. Proc. Inst. Zool. 1961, 17, 71–85. (In Ukrainian) [Google Scholar]
- Puchkov, V.G. Macrolophus Fieber, 1858 species (Heteroptera, Miridae) of the USSR. Dokl. Akad. Nauk Ukr. SSR Ser. B. Geol. Chem. Biol. 1978, 9, 854–857. (In Russian) [Google Scholar]
- Cobben, R.H. Evolutionary Trends in Heteroptera. Part I. Eggs, Architecture of the Shell, Gross Embryology and Eclosion; Centre for Agricultural Publishing and Documentation: Wageningen, The Netherlands, 1968. [Google Scholar]
- Kullenberg, B. Studien über die Biologie der Capsiden. Zool. Bidr. Upps. 1944, 20, 1–522. [Google Scholar]
- Wheeler, A.G., Jr. Biology of the Plant Bugs (Hemiptera: Miridae): Pests, Predators, Opportunists; Cornell Univ. Press: Ithaca, NY, USA, 2001. [Google Scholar]
- Saulich, A.K.; Musolin, D.L. Seasonal development of plant bugs (Heteroptera, Miridae): Subfamily Mirinae, tribe Mirini. Entomol. Rev. 2020, 100, 135–156. [Google Scholar] [CrossRef]
- Saulich, A.K.; Musolin, D.L. Seasonal development of plant bugs (Heteroptera, Miridae): Subfamily Mirinae, tribe Stenodemini. Entomol. Rev. 2021, 101, 147–161. Available online: https://link.springer.com/article/10.1134/S0013873821020019 (accessed on 10 May 2024). [CrossRef]
- Musolin, D.L.; Saulich, A.K. Seasonal development of plant bugs (Heteroptera, Miridae): Subfamily Orthotylinae, tribes Halticini and Orthotylini. Entomol. Rev. 2021, 101, 743–754. Available online: https://link.springer.com/article/10.1134/S0013873821060026 (accessed on 10 May 2024). [CrossRef]
- Musolin, D.L.; Saulich, A.K. Seasonal development of plant bugs (Heteroptera, Miridae): Subfamily Phylinae, tribes Pilophorini, Hallodapini and Phylini. Entomol. Rev. 2022, 102, 20–40. [Google Scholar] [CrossRef]
- Blümel, S. Effect of selected mass-rearing parameters on O. majusculus (Reuter) and O. laevigatus (Fieber). IOBC WPRS Bull. 1996, 19, 15–18. [Google Scholar]
- Bueno, V.H.P.; Carvalho, L.M.; Lenteren, J.V. Performance of Orius insidiosus after storage, exposure to dispersal material, handling and shipment processes. Bull. Insectol. 2014, 67, 175–183. [Google Scholar]
- Coudron, T.A.; Ellersieck, M.R.; Shelby, K.S. Influence of diet on long-term cold storage of the predator Podisus maculiventris (Say) (Heteroptera: Pentatomidae). Biol. Contr. 2007, 42, 186–195. [Google Scholar] [CrossRef]
- De Clercq, P.; Degheele, D. Cold storage of the predatory bugs Podisus maculiventris (Say) and Podisus sagitta (Fabricius) (Heteroptera: Pentatomidae). Parasitica 1993, 49, 27–41. [Google Scholar]
- Guo, J.Y.; Wu, M.; Wan, F.H. Effects of cold storage on the adults and eggs of Orius sauteri. Chin. J. Biol. Contr. 2002, 18, 10–12. [Google Scholar]
- Ito, K.; Nakata, T. Diapause and survival in winter in two species of predatory bugs, Orius sauteri and O. minutus. Entomol. Exp. Appl. 1998, 89, 271–276. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, H.Y.; Han, M.J.; Choi, M.; Hwang, S.J.; Lee, M.S. Cold storage effect on the biological characteristics of Orius laevigatus (Fieber) (Hemiptera: Anthocoridae) and Phytoseiulus persimilis Athias-Henriot (Acari: Phytoseiidae). Korean J. Appl. Entomol. 2009, 48, 361–368. [Google Scholar] [CrossRef]
- Pazyuk, I.M.; Dolgovskaya, M.Y.; Reznik, S.Y.; Musolin, D.L. Is diapause not needed? Long-term storage potential of zoophytophagous plant bug Dicyphus errans in relation to day length, temperature, and trophic regimes. Biocontrol Sci. Technol. 2021, 31, 53–64. [Google Scholar] [CrossRef]
- Rudolf, E.; Malausa, J.C.; Millot, P.; Pralavorio, R. Influence des basses températures sur les potentialités biologiques d’Orius laevigatus et d’Orius majusculus (Het.: Anthocoridae). Entomophaga 1993, 38, 317–325. [Google Scholar] [CrossRef]
- Seo, M.; Kim, J.H.; Seo, B.Y.; Park, B.; Lee, S.G.; Kim, K.H.; Cho, J.R. Effect of cold storage on quality of Geocoris pallidipennis and Micromus angulatus. Korean J. Appl. Entomol. 2019, 58, 151–157. [Google Scholar] [CrossRef]
- Bahşi, Ş.Ü.; Tunc, İ. Optimization of Orius majusculus release: Photoperiodic sensitivity at different temperatures and storage of diapausing adults. Turk. J. Agric. For. 2014, 38, 935–941. [Google Scholar] [CrossRef]
- Yanik, E.; Ünlü, L. Storage studies of different stages of Anthocoris minki Dohrn (Hemiptera: Anthocoridae) under low temperatures. Turk. Entomol. Derg. 2015, 39, 277–286. Available online: https://dergipark.org.tr/tr/download/article-file/65304 (accessed on 10 May 2024). [CrossRef]
- Franco, K.; Jauset, A.; Castañé, C. Monogamy and polygamy in two species of mirid bugs: A functional-based approach. J. Insect Physiol. 2011, 57, 307–315. [Google Scholar] [CrossRef] [PubMed]
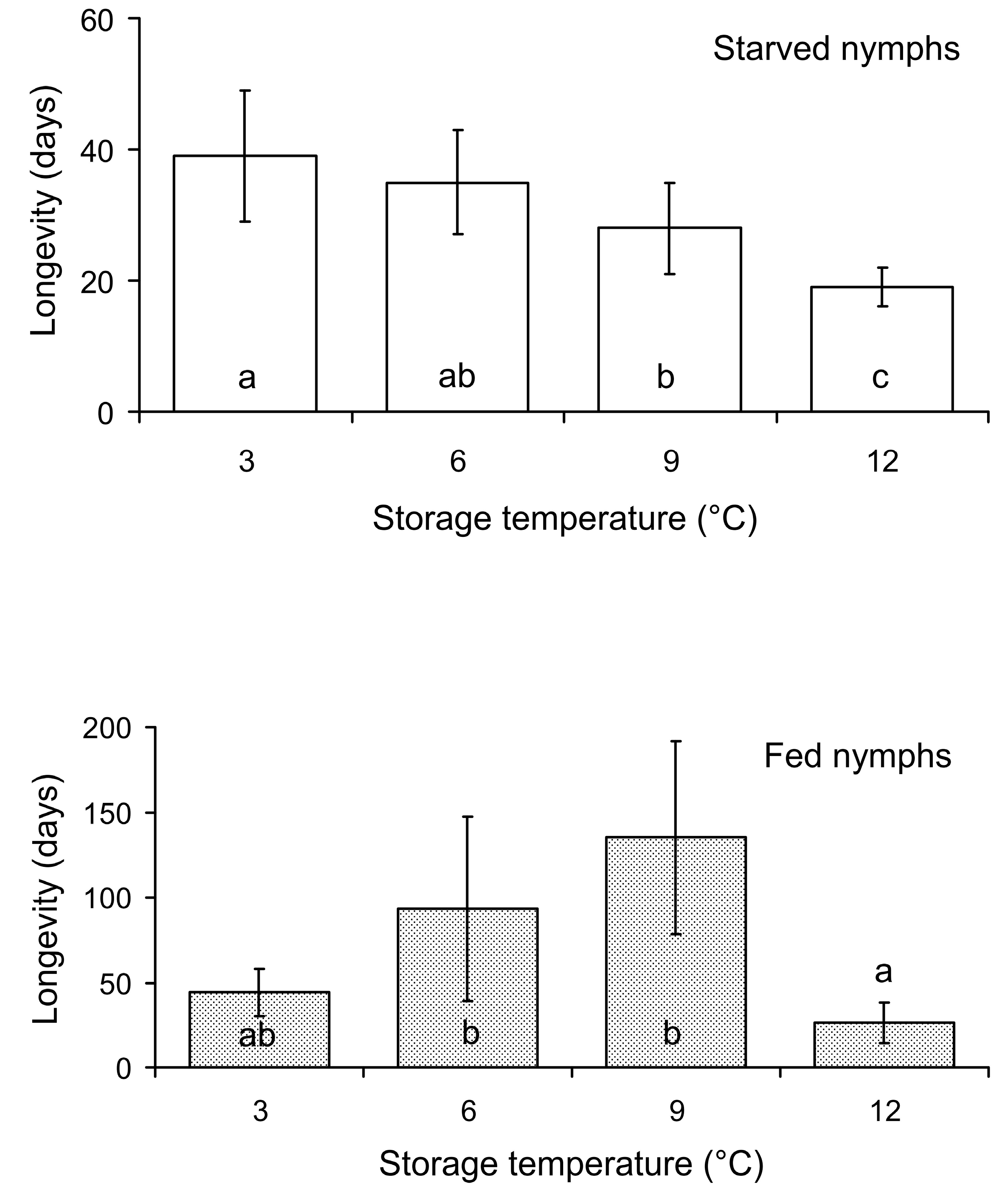
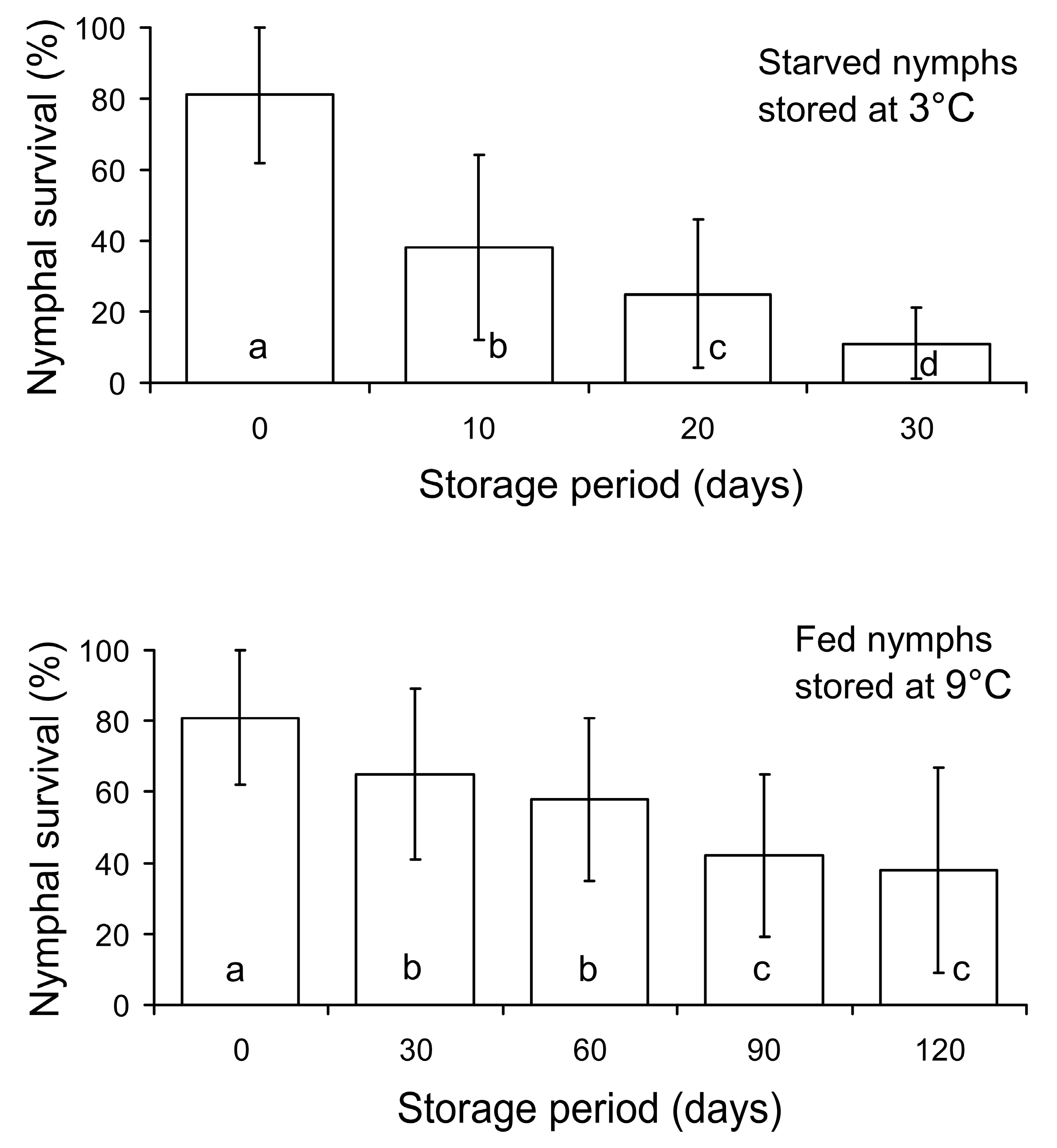


{kind=link}
{kind=link}
{kind=link}
| A Factor or a Combination of Two Factors | The Results of Four-Way ANOVA on Ranked Data (n = 68, dferror = 36): The Number of Degrees of Freedom (df), Fisher Coefficient (F), and p-Value |
|---|---|
| Temperature during development | df = 1, F = 2.8, p = 0.103 |
| Photoperiod during development | df = 1, F = 1.3, p = 0.254 |
| Temperature during storage | df = 3, F = 18.9, p < 0.001 |
| Feeding during storage | df = 1, F = 52.7, p < 0.001 |
| Temperature during development × Photoperiod during development | df = 1, F = 3.1, p = 0.088 |
| Temperature during development × Temperature during storage | df = 3, F = 0.7, p = 0.566 |
| Temperature during development × Feeding during storage | df = 1, F = 8.9, p = 0.005 |
| Photoperiod during development × Temperature during storage | df = 3, F = 3.3, p = 0.032 |
| Photoperiod during development × Feeding during storage | df = 3, F = 5.3, p = 0.027 |
| Temperature during storage × Feeding during storage | df = 3, F = 8.7, p < 0.001 |
| Factors | The Results of GLM Analysis on Ranked Data (n = 68): The Coefficient of Regression ± SE and p-Value |
|---|---|
| Temperature during development | C = –0.684 ± 0.988, p = 0.491 |
| Photoperiod during development | C = –0.923 ± 0.661, p = 0.167 |
| Temperature during storage | C = –2.197 ± 0.646, p = 0.001 |
| Feeding during storage | C = –4.079 ± 0.527, p < 0.001 |
| The number of nymphs per Petri dish | C = –7.172 ± 4.783, p = 0.139 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pazyuk, I.M.; Dolgovskaya, M.Y.; Reznik, S.Y.; Musolin, D.L. Storing up Treasures: Storage Potential of Macrolophus pygmaeus (Hemiptera: Heteroptera: Miridae) Nymphs for Application in Biological Control. Insects 2024, 15, 414. https://doi.org/10.3390/insects15060414
Pazyuk IM, Dolgovskaya MY, Reznik SY, Musolin DL. Storing up Treasures: Storage Potential of Macrolophus pygmaeus (Hemiptera: Heteroptera: Miridae) Nymphs for Application in Biological Control. Insects. 2024; 15(6):414. https://doi.org/10.3390/insects15060414
Chicago/Turabian StylePazyuk, Irina M., Margarita Y. Dolgovskaya, Sergey Y. Reznik, and Dmitrii L. Musolin. 2024. "Storing up Treasures: Storage Potential of Macrolophus pygmaeus (Hemiptera: Heteroptera: Miridae) Nymphs for Application in Biological Control" Insects 15, no. 6: 414. https://doi.org/10.3390/insects15060414