


Differential Modulation of Saliva-Derived Microcosm Biofilms by Antimicrobial Peptide LL-31 and D-LL-31

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Saliva Collection
2.2. Peptide Synthesis
2.3. Bacterial Strains and Growth Conditions
2.4. Bacterial Growth Inhibition by LL-31 and D-LL-31
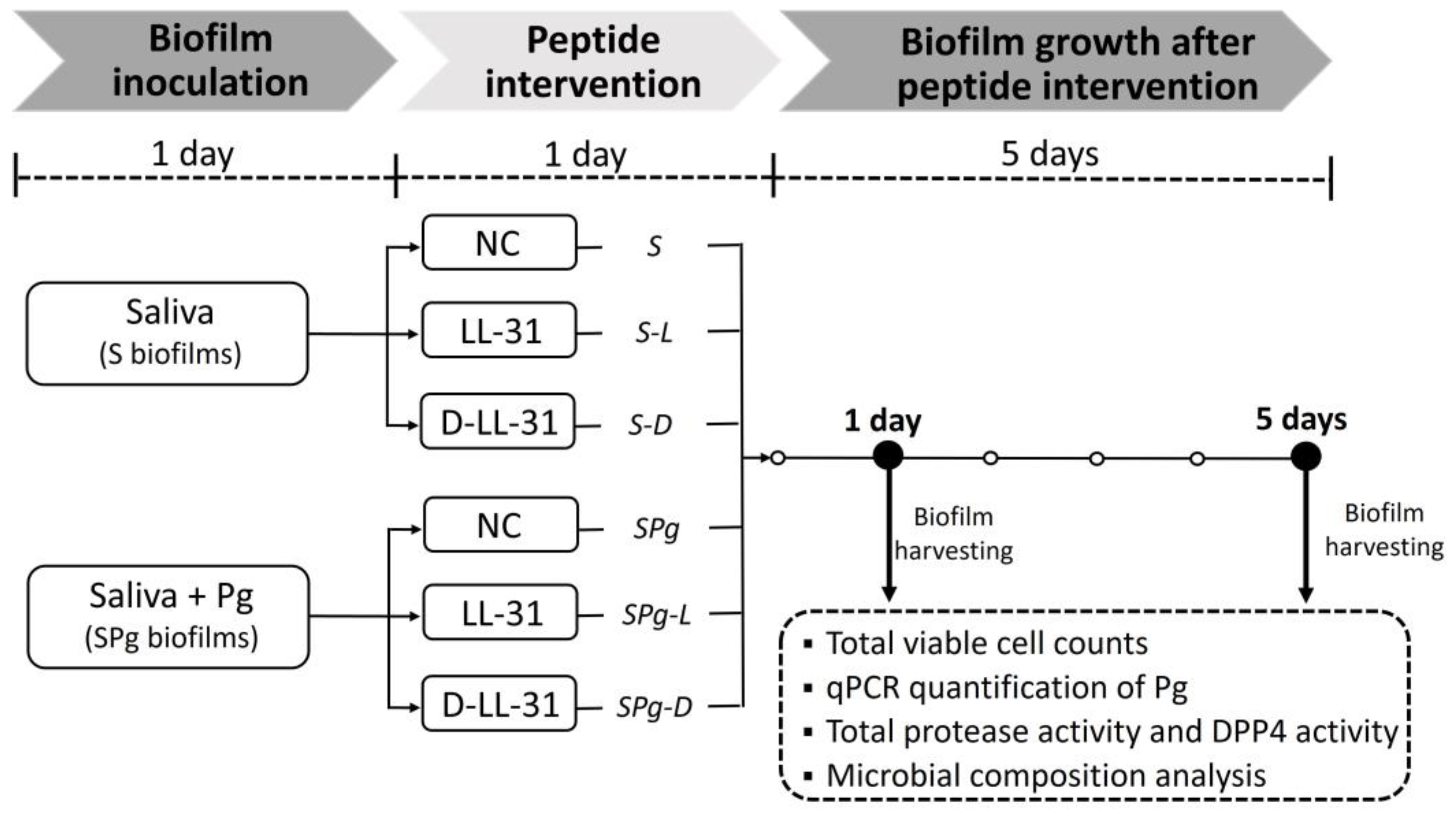
2.5. Microcosm Biofilm Formation and Peptide Intervention
2.6. Total Viable Cell Counts
2.7. Total Protease Activity and DPP4 Activity
2.8. gDNA Isolation and Quantification of P. gingivalis
2.9. 16S rRNA Gene Amplicon Sequencing and Data Processing
2.10. Data Analysis and Statistics
3. Results
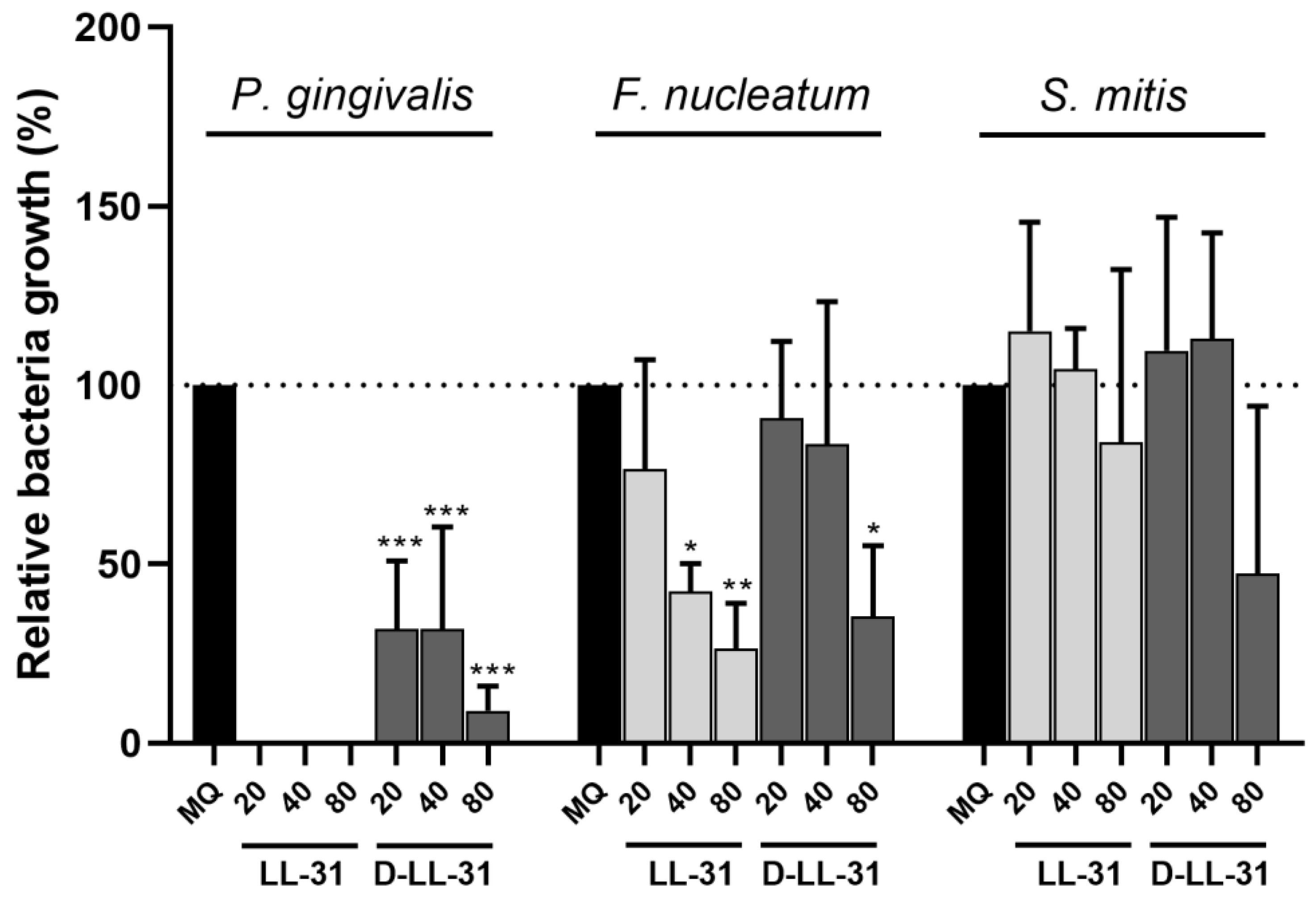
3.1. Bacterial Growth Inhibition by LL-31 and D-LL-31
3.2. Total Viable Cell Counts and the Amounts of P. gingivalis in Biofilms
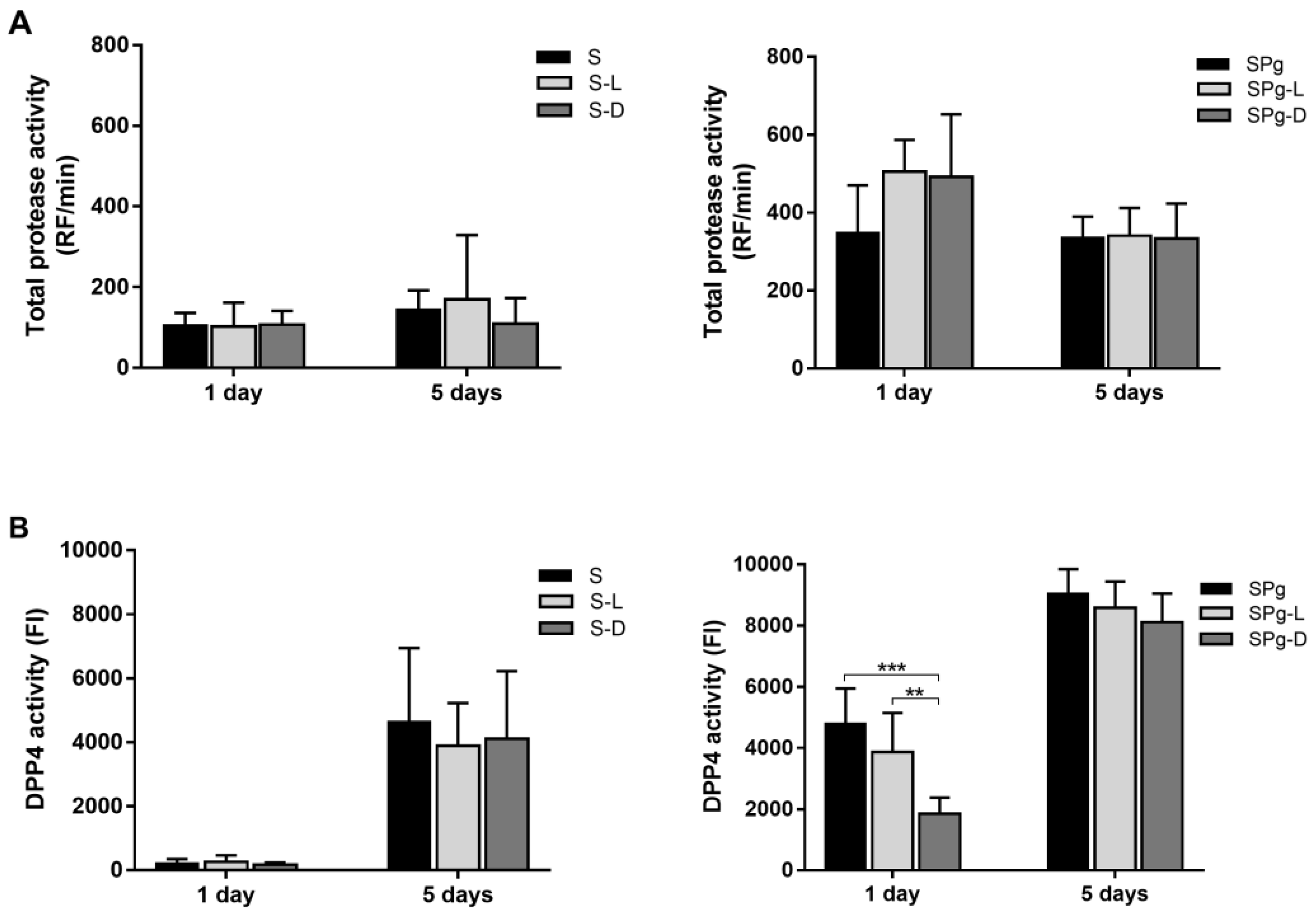
3.3. Total Protease Activity and DPP4 Activity of the Biofilms
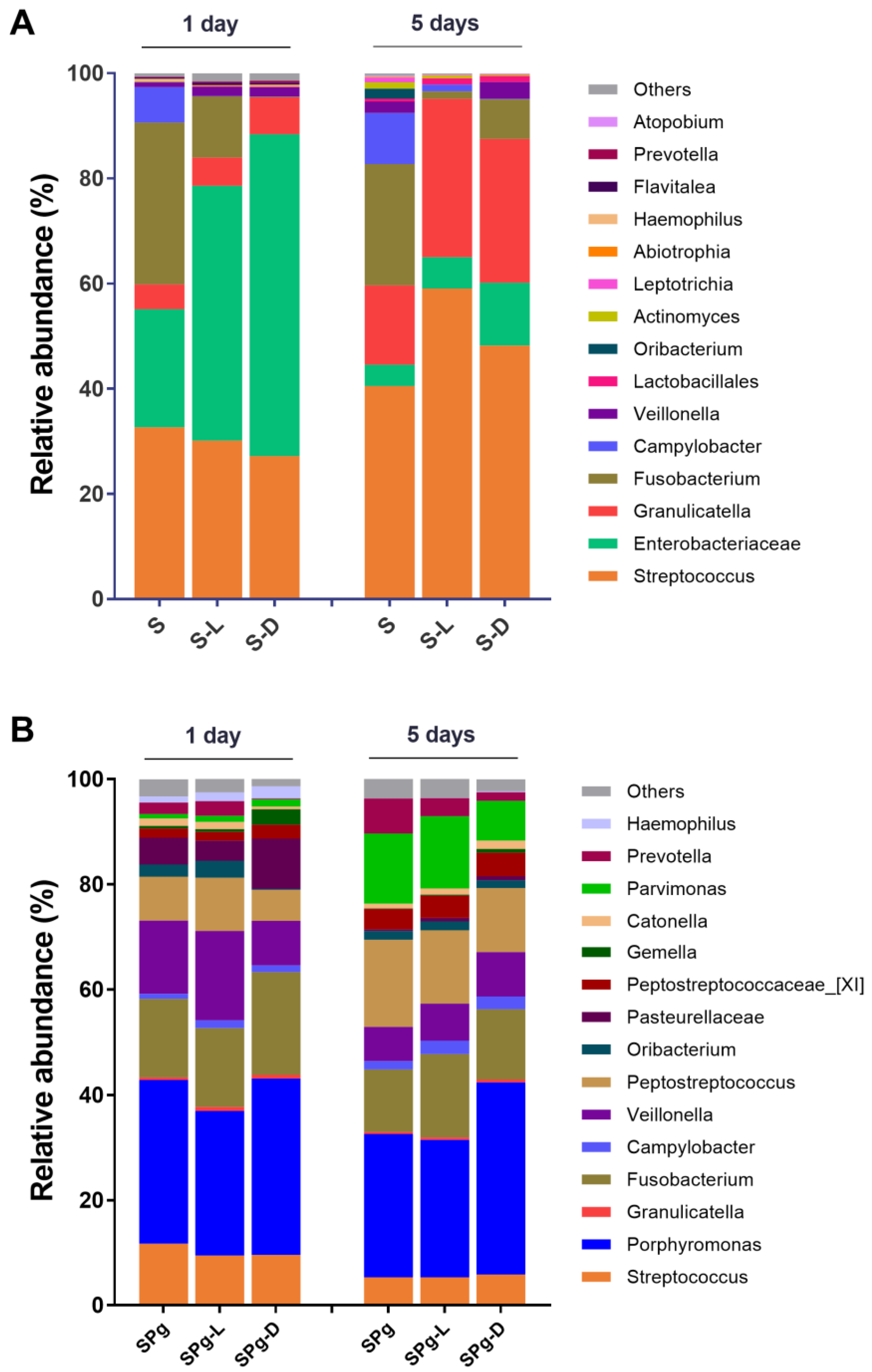
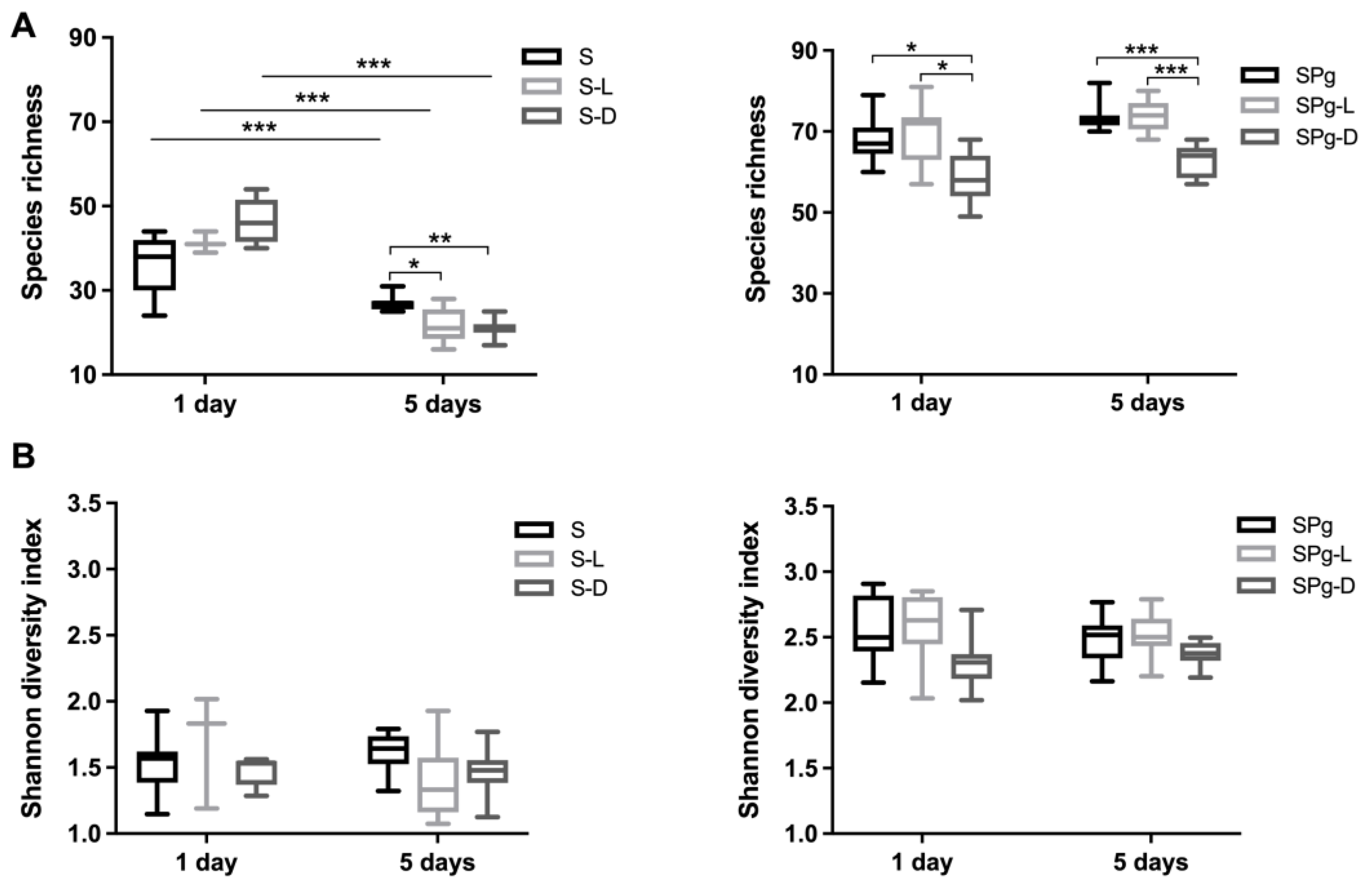
3.4. Major Bacterial Genera and α-Diversity of the Biofilms
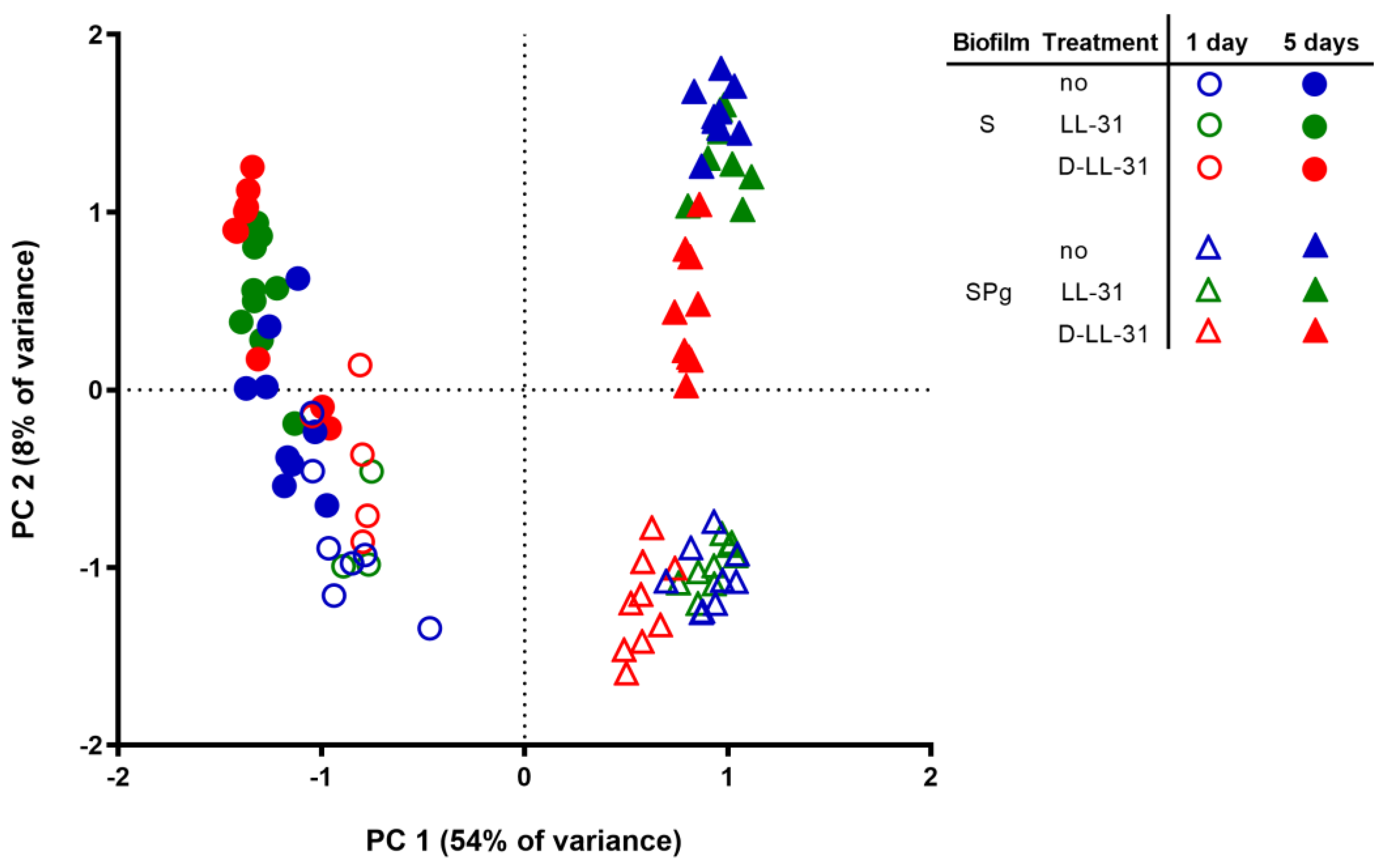
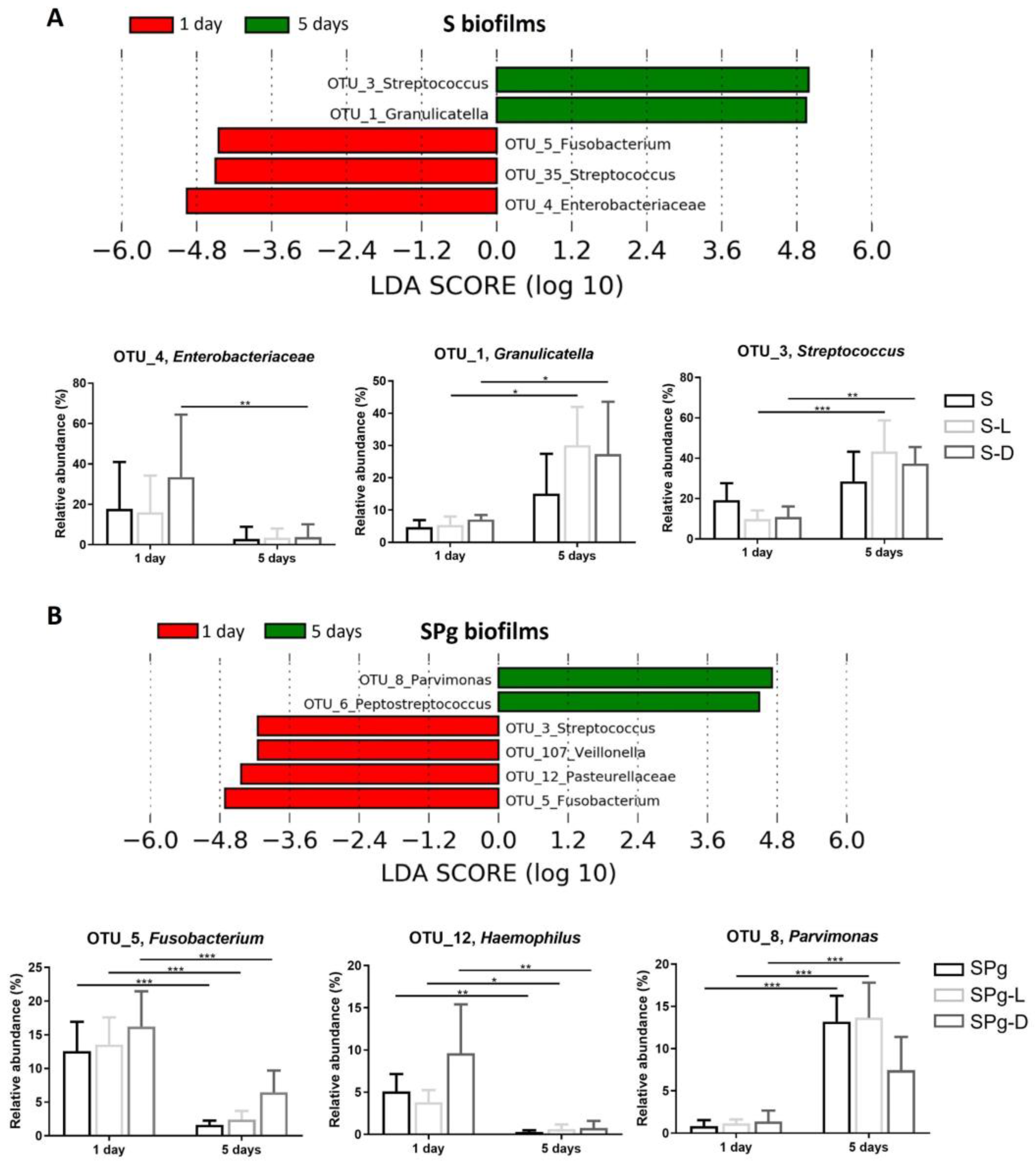
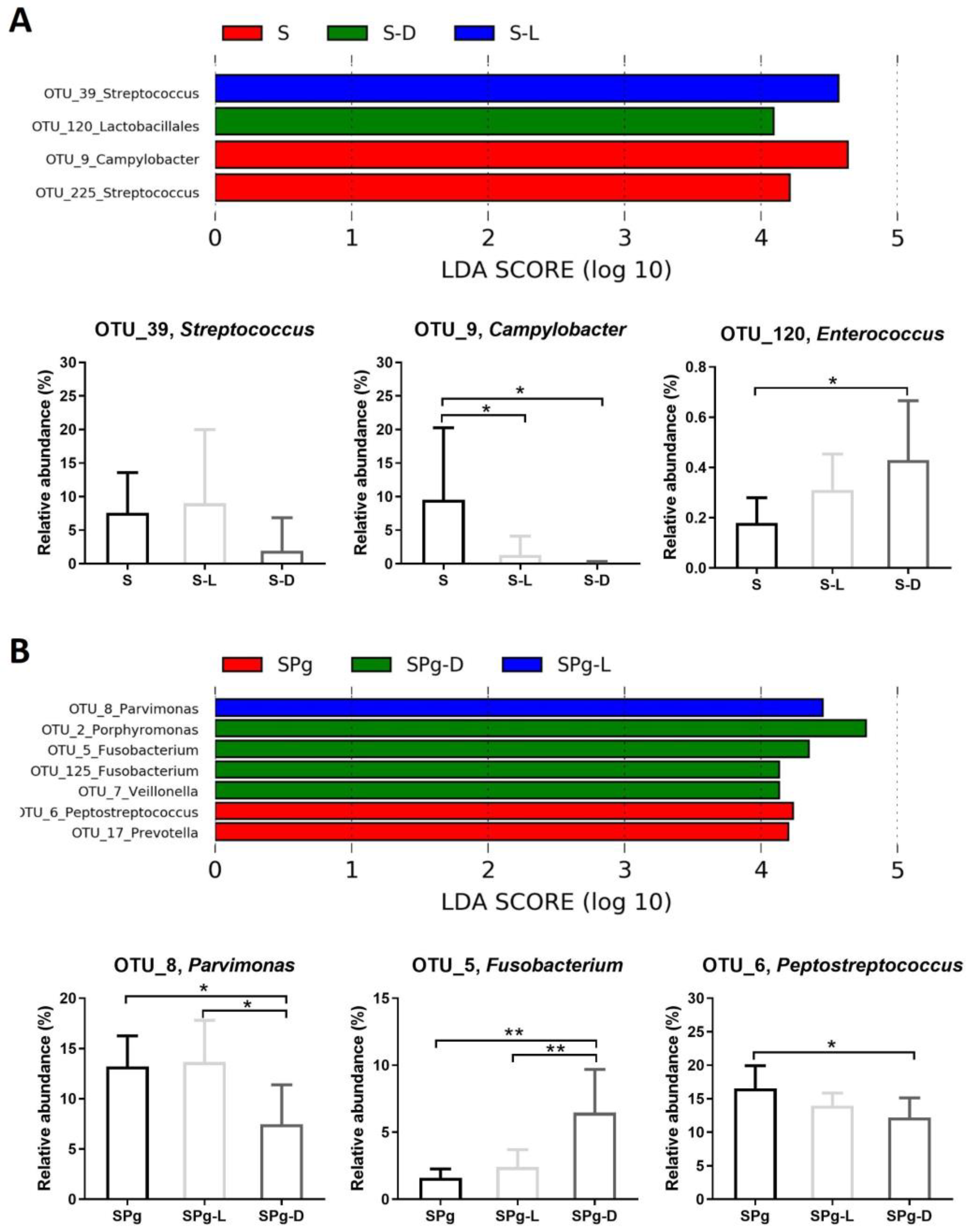
3.5. Microbial Composition of the Biofilms
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Trindade, D.; Carvalho, R.; Machado, V.; Chambrone, L.; Mendes, J.J.; Botelho, J. Prevalence of periodontitis in dentate people between 2011 and 2020: A systematic review and meta-analysis of epidemiological studies. J. Clin. Periodontol. 2023, 50, 604–626. [Google Scholar] [CrossRef] [PubMed]
- Caffesse, R.G.; Echeverría, J.J. Treatment trends in periodontics. J. Clin. Periodontol. 2019, 79, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Alassy, H.; Pizarek, J.A.; Kormas, I.; Pedercini, A.; Wolff, L.F. Antimicrobial adjuncts in the management of periodontal and peri-implant diseases and conditions: A narrative review. Front. Oral. Maxillofac. Med. 2021, 3, 16. [Google Scholar] [CrossRef]
- Khattri, S.; Kumbargere Nagraj, S.; Arora, A.; Eachempati, P.; Kusum, C.K.; Bhat, K.G.; Johnson, T.M.; Lodi, G. Adjunctive systemic antimicrobials for the non-surgical treatment of periodontitis. Cochrane Database Syst. Rev. 2020, 11, Cd012568. [Google Scholar] [PubMed]
- Ramanauskaite, E.; Moraschini, V.; Machiulskiene, V.; Sculean, A. Clinical efficacy of single and multiple applications of antimicrobial photodynamic therapy in periodontal maintenance: A systematic review and network meta-analysis. Photodiagnosis Photodyn. Ther. 2021, 36, 102435. [Google Scholar] [CrossRef]
- Balan, P.; Belibasakis, G.; Ivanovski, S.; Bostanci, N.; Seneviratne, C.J. Community dynamics of subgingival microbiome in periodontitis and targets for microbiome modulation therapy. Crit. Rev. Microbiol. 2022, 49, 726–738. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Darveau, R.P.; Curtis, M.A. The keystone-pathogen hypothesis. Nat. Rev. Microbiol. 2012, 10, 717–725. [Google Scholar]
- Hajishengallis, G.; Lamont, R.J. Beyond the red complex and into more complexity: The polymicrobial synergy and dysbiosis (PSD) model of periodontal disease etiology. Mol. Oral. Microbiol. 2012, 27, 409–419. [Google Scholar] [CrossRef]
- Horiuchi, A.; Kokubu, E.; Warita, T.; Ishihara, K. Synergistic biofilm formation by Parvimonas micra and Fusobacterium nucleatum. Anaerobe 2020, 62, 102100. [Google Scholar] [CrossRef]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar]
- Gubatan, J.; Holman, D.R.; Puntasecca, C.J.; Polevoi, D.; Rubin, S.J.; Rogalla, S. Antimicrobial peptides and the gut microbiome in inflammatory bowel disease. World J. Gastroenterol. 2021, 27, 7402–7422. [Google Scholar]
- Salzman, N.H.; Hung, K.; Haribhai, D.; Chu, H.; Karlsson-Sjöberg, J.; Amir, E.; Teggatz, P.; Barman, M.; Hayward, M.; Eastwood, D.; et al. Enteric defensins are essential regulators of intestinal microbial ecology. Nat. Immunol. 2010, 11, 76–83. [Google Scholar] [PubMed]
- Bevins, C.L.; Salzman, N.H. Paneth cells, antimicrobial peptides and maintenance of intestinal homeostasis. Nat. Rev. Microbiol. 2011, 9, 356–368. [Google Scholar]
- Wang, G.; Narayana, J.L.; Mishra, B.; Zhang, Y.; Wang, F.; Wang, C.; Zarena, D.; Lushnikova, T.; Wang, X. Design of Antimicrobial Peptides: Progress Made with Human Cathelicidin LL-37. In Antimicrobial Peptides: Basics for Clinical Application; Matsuzaki, K., Ed.; Springer: Singapore, 2019; pp. 215–240. [Google Scholar]
- Liang, W.; Enée, E.; Andre-Vallee, C.; Falcone, M.; Sun, J.; Diana, J. Intestinal Cathelicidin Antimicrobial Peptide Shapes a Protective Neonatal Gut Microbiota Against Pancreatic Autoimmunity. Gastroenterology 2022, 162, 1288–1302.e1216. [Google Scholar]
- Pütsep, K.; Carlsson, G.; Boman, H.G.; Andersson, M. Deficiency of antibacterial peptides in patients with morbus Kostmann: An observation study. Lancet 2002, 360, 1144–1149. [Google Scholar]
- Nilsson, B.-O. What can we learn about functional importance of human antimicrobial peptide LL-37 in the oral environment from severe congenital neutropenia (Kostmann disease)? Peptides 2020, 128, 170311. [Google Scholar] [PubMed]
- Zasloff, M. Innate immunity, antimicrobial peptides, and protection of the oral cavity. Lancet 2002, 360, 1116–1117. [Google Scholar]
- Kanthawong, S.; Bolscher, J.G.M.; Veerman, E.C.I.; van Marle, J.; Nazmi, K.; Wongratanacheewin, S.; Taweechaisupapong, S. Antimicrobial activities of LL-37 and its truncated variants against Burkholderia thailandensis. Int. J. Antimicrob. Agents 2010, 36, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Kaman, W.E.; Voskamp-Visser, I.; de Jongh, D.M.C.; Endtz, H.P.; van Belkum, A.; Hays, J.P.; Bikker, F.J. Evaluation of a D-amino-acid-containing fluorescence resonance energy transfer peptide library for profiling prokaryotic proteases. Anal. Biochem. 2013, 441, 38–43. [Google Scholar] [CrossRef]
- Wongkaewkhiaw, S.; Taweechaisupapong, S.; Anutrakunchai, C.; Nazmi, K.; Bolscher, J.G.M.; Wongratanacheewin, S.; Kanthawong, S. D-LL-31 in combination with ceftazidime synergistically enhances bactericidal activity and biofilm destruction in Burkholderia pseudomallei. Biofouling 2019, 35, 573–584. [Google Scholar]
- Jiang, Y.; Brandt, B.W.; Buijs, M.J.; Cheng, L.; Exterkate, R.A.M.; Crielaard, W.; Deng, D.M. Manipulation of Saliva-Derived Microcosm Biofilms to Resemble Dysbiotic Subgingival Microbiota. Appl. Environ. Microbiol. 2021, 87, e02371-20. [Google Scholar] [CrossRef] [PubMed]
- Wongkaewkhiaw, S.; Taweechaisupapong, S.; Thanaviratananich, S.; Bolscher, J.G.M.; Nazmi, K.; Anutrakunchai, C.; Chareonsudjai, S.; Kanthawong, S. D-LL-31 enhances biofilm-eradicating effect of currently used antibiotics for chronic rhinosinusitis and its immunomodulatory activity on human lung epithelial cells. PLoS ONE 2020, 15, e0243315. [Google Scholar]
- Puknun, A.; Bolscher, J.G.M.; Nazmi, K.; Veerman, E.C.I.; Tungpradabkul, S.; Wongratanacheewin, S.; Kanthawong, S.; Taweechaisupapong, S. A heterodimer comprised of two bovine lactoferrin antimicrobial peptides exhibits powerful bactericidal activity against Burkholderia pseudomallei. World J. Microbiol. Biotechnol. 2013, 29, 1217–1224. [Google Scholar] [PubMed]
- Thompson, H.; Rybalka, A.; Moazzez, R.; Dewhirst, F.E.; Wade, W.G. In vitro culture of previously uncultured oral bacterial phylotypes. Appl. Environ. Microbiol. 2015, 81, 8307–8314. [Google Scholar] [CrossRef]
- Janus, M.M.; Keijser, B.J.; Bikker, F.J.; Exterkate, R.A.; Crielaard, W.; Krom, B.P. In vitro phenotypic differentiation towards commensal and pathogenic oral biofilms. Biofouling 2015, 31, 503–510. [Google Scholar] [CrossRef]
- Boutaga, K.; van Winkelhoff, A.J.; Vandenbroucke-Grauls, C.M.J.E.; Savelkoul, P.H.M. Comparison of real-time PCR and culture for detection of Porphyromonas gingivalis in subgingival plaque samples. J. Clin. Microbiol. 2003, 41, 4950–4954. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261. [Google Scholar]
- Chen, T.; Yu, W.-H.; Izard, J.; Baranova, O.V.; Lakshmanan, A.; Dewhirst, F.E. The Human Oral Microbiome Database: A web accessible resource for investigating oral microbe taxonomic and genomic information. Database 2010, 2010, baq013. [Google Scholar] [PubMed]
- Hammer, Ø.; Harper, D.; Ryan, P. PAST-palaeontological statistics, ver. 1.89. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Aemaimanan, P.; Sattayasai, N.; Wara-aswapati, N.; Pitiphat, W.; Suwannarong, W.; Prajaneh, S.; Taweechaisupapong, S. Alanine Aminopeptidase and Dipeptidyl Peptidase IV in Saliva of Chronic Periodontitis Patients. J. Periodontol. 2009, 80, 1809–1814. [Google Scholar] [CrossRef]
- Eley, B.M.; Cox, S.W. Correlation between gingival crevicular fluid dipeptidyl peptidase II and IV activity and periodontal attachment loss. A 2–year longitudinal study in chronic periodontitis patients. Oral. Dis. 1995, 1, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Fjell, C.D.; Hiss, J.A.; Hancock, R.E.W.; Schneider, G. Designing antimicrobial peptides: Form follows function. Nat. Rev. Drug Discov. 2012, 11, 37–51. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, M.; Qiu, S.; Wang, J.; Peng, J.; Zhao, P.; Zhu, R.; Wang, H.; Li, Y.; Wang, K.; et al. Antimicrobial activity and stability of the D-amino acid substituted derivatives of antimicrobial peptide polybia-MPI. AMB Express 2016, 6, 122. [Google Scholar] [CrossRef]
- Zhong, C.; Zhu, N.; Zhu, Y.; Liu, T.; Gou, S.; Xie, J.; Yao, J.; Ni, J. Antimicrobial peptides conjugated with fatty acids on the side chain of D-amino acid promises antimicrobial potency against multidrug-resistant bacteria. Eur. J. Pharm. Sci. 2020, 141, 105123. [Google Scholar] [CrossRef]
- Huang, L.; van Loveren, C.; Ling, J.; Wei, X.; Crielaard, W.; Deng, D.M. Epithelial cell detachment by Porphyromonas gingivalis biofilm and planktonic cultures. Biofouling 2016, 32, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liang, W.; Gong, W.; Yoshimura, T.; Chen, K.; Wang, J.M. The Critical Role of the Antimicrobial Peptide LL-37/CRAMP in Protection of Colon Microbiota Balance, Mucosal Homeostasis, Anti-Inflammatory Responses, and Resistance to Carcinogenesis. Crit. Rev. Immunol. 2019, 39, 83–92. [Google Scholar] [CrossRef]
- Radaic, A.; Ye, C.; Parks, B.; Gao, L.; Kuraji, R.; Malone, E.; Kamarajan, P.; Zhan, L.; Kapila, Y.L. Modulation of pathogenic oral biofilms towards health with nisin probiotic. J. Oral. Microbiol. 2020, 12, 1809302. [Google Scholar] [CrossRef]
- Olsen, I.; Lambris, J.D.; Hajishengallis, G. Porphyromonas gingivalis disturbs host-commensal homeostasis by changing complement function. J. Oral. Microbiol. 2017, 9, 1340085. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G.; Liang, S.; Payne, M.A.; Hashim, A.; Jotwani, R.; Eskan, M.A.; McIntosh, M.L.; Alsam, A.; Kirkwood, K.L.; Lambris, J.D.; et al. Low-abundance biofilm species orchestrates inflammatory periodontal disease through the commensal microbiota and complement. Cell Host Microbe 2011, 10, 497–506. [Google Scholar] [CrossRef]
- Payne, M.A.; Hashim, A.; Alsam, A.; Joseph, S.; Aduse-Opoku, J.; Wade, W.G.; Curtis, M.A. Horizontal and Vertical Transfer of Oral Microbial Dysbiosis and Periodontal Disease. J. Dent. Res. 2019, 98, 1503–1510. [Google Scholar] [CrossRef] [PubMed]
- Kremer, B.H.A.; van Steenbergen, T.J.M. Peptostreptococcus micros coaggregates with Fusobacterium nucleatum and non-encapsulated Porphyromonas gingivalis. FEMS Microbiol. Lett. 2000, 182, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Neilands, J.; Davies, J.R.; Bikker, F.J.; Svensäter, G. Parvimonas micra stimulates expression of gingipains from Porphyromonas gingivalis in multi-species communities. Anaerobe 2019, 55, 54–60. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biofilm | Age | Treatment | F Value 1 | p Value 1 | ||
|---|---|---|---|---|---|---|
| S | 1 day | no | vs. | LL-31 | 1.0 | 1.000 |
| no | vs. | D-LL-31 | 2.2 | 0.074 | ||
| LL-31 | vs. | D-LL-31 | 0.6 | 1.000 | ||
| 5 days | no | vs. | LL-31 | 3.5 | 0.008 | |
| no | vs. | D-LL-31 | 5.4 | 0.002 | ||
| LL-31 | vs. | D-LL-31 | 0.8 | 0.575 | ||
| SPg | 1 day | no | vs. | LL-31 | 2.0 | 0.071 |
| no | vs. | D-LL-31 | 15.0 | 0.0003 | ||
| LL-31 | vs. | D-LL-31 | 12.3 | 0.0003 | ||
| 5 days | no | vs. | LL-31 | 1.9 | 0.072 | |
| no | vs. | D-LL-31 | 8.8 | 0.001 | ||
| LL-31 | vs. | D-LL-31 | 5.6 | 0.001 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soldati, K.R.; Jiang, Y.; Brandt, B.W.; Exterkate, R.A.M.; Buijs, M.J.; Nazmi, K.; Kaman, W.E.; Cheng, L.; Bikker, F.J.; Crielaard, W.; et al. Differential Modulation of Saliva-Derived Microcosm Biofilms by Antimicrobial Peptide LL-31 and D-LL-31. Pathogens 2023, 12, 1295. https://doi.org/10.3390/pathogens12111295
Soldati KR, Jiang Y, Brandt BW, Exterkate RAM, Buijs MJ, Nazmi K, Kaman WE, Cheng L, Bikker FJ, Crielaard W, et al. Differential Modulation of Saliva-Derived Microcosm Biofilms by Antimicrobial Peptide LL-31 and D-LL-31. Pathogens. 2023; 12(11):1295. https://doi.org/10.3390/pathogens12111295
Chicago/Turabian StyleSoldati, Kahena R., Yaling Jiang, Bernd W. Brandt, Rob A. M. Exterkate, Mark J. Buijs, Kamran Nazmi, Wendy E. Kaman, Lei Cheng, Floris J. Bikker, Wim Crielaard, and et al. 2023. "Differential Modulation of Saliva-Derived Microcosm Biofilms by Antimicrobial Peptide LL-31 and D-LL-31" Pathogens 12, no. 11: 1295. https://doi.org/10.3390/pathogens12111295
APA StyleSoldati, K. R., Jiang, Y., Brandt, B. W., Exterkate, R. A. M., Buijs, M. J., Nazmi, K., Kaman, W. E., Cheng, L., Bikker, F. J., Crielaard, W., Zandim-Barcelos, D. L., & Deng, D. M. (2023). Differential Modulation of Saliva-Derived Microcosm Biofilms by Antimicrobial Peptide LL-31 and D-LL-31. Pathogens, 12(11), 1295. https://doi.org/10.3390/pathogens12111295