
Protozoan Parasites of Iranian Freshwater Fishes: Review, Composition, Classification, and Modeling Distribution



Abstract
1. Introduction
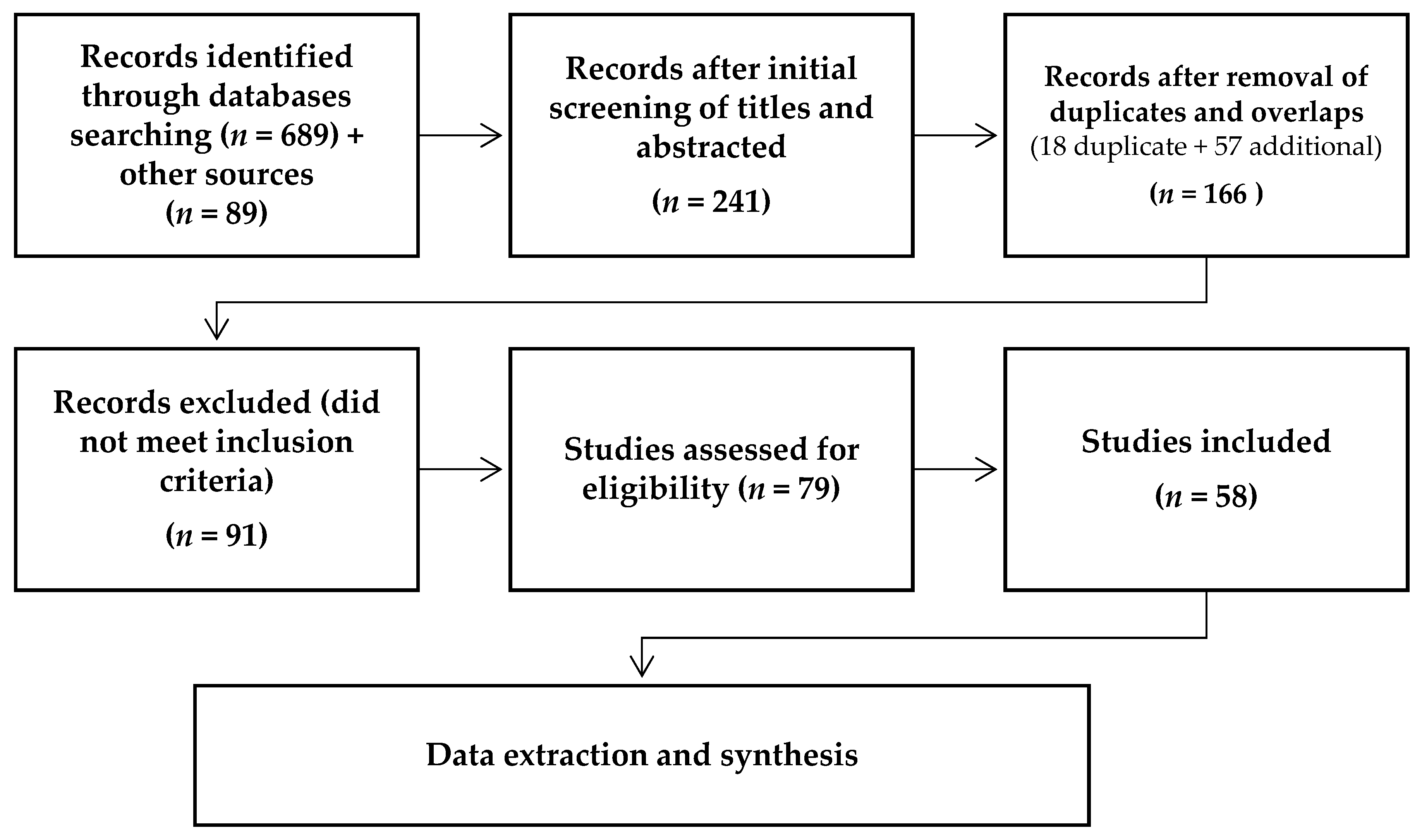
2. Materials and Methods
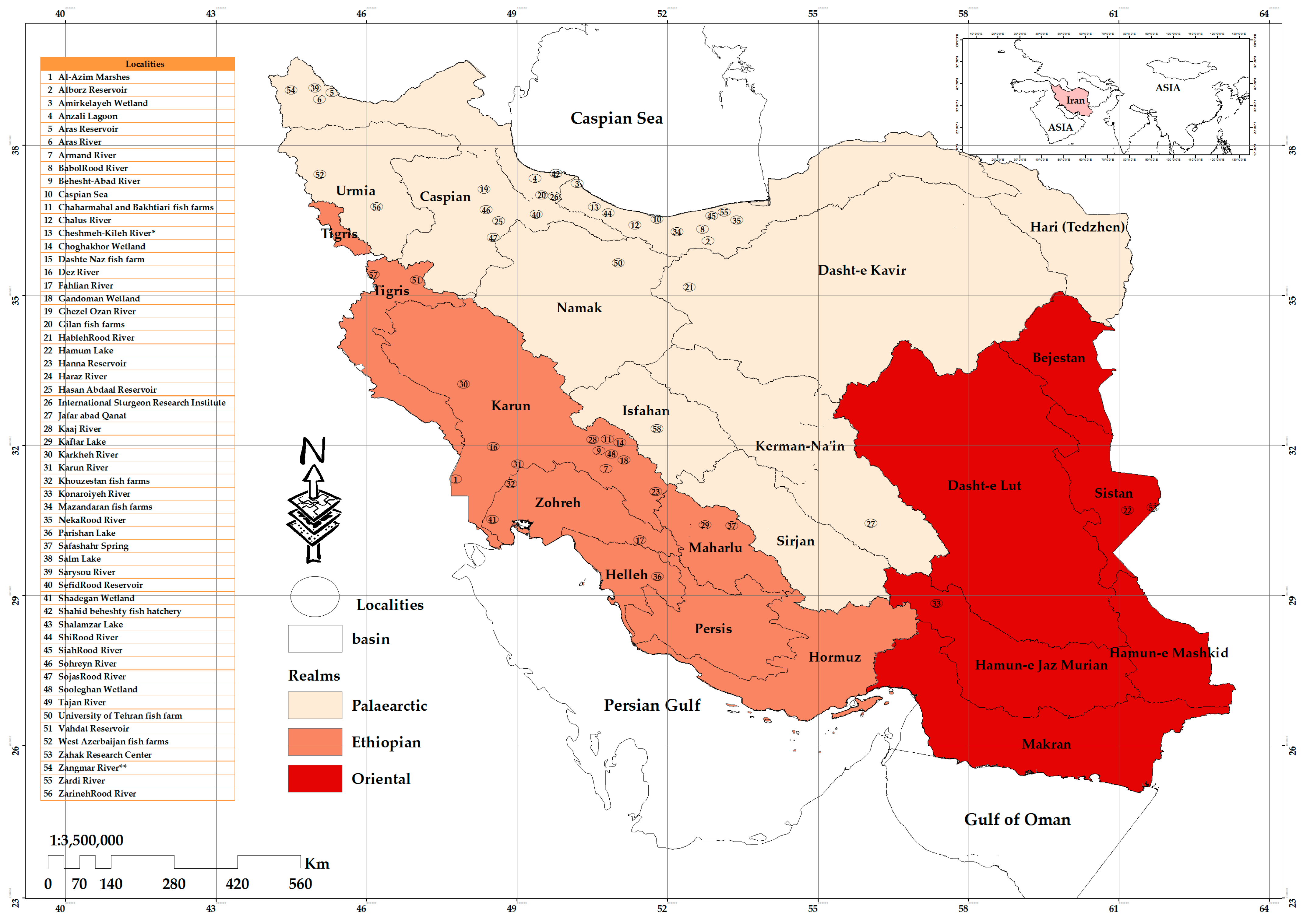
2.1. Study Area
2.2. Search Strategy
2.3. Data Extraction
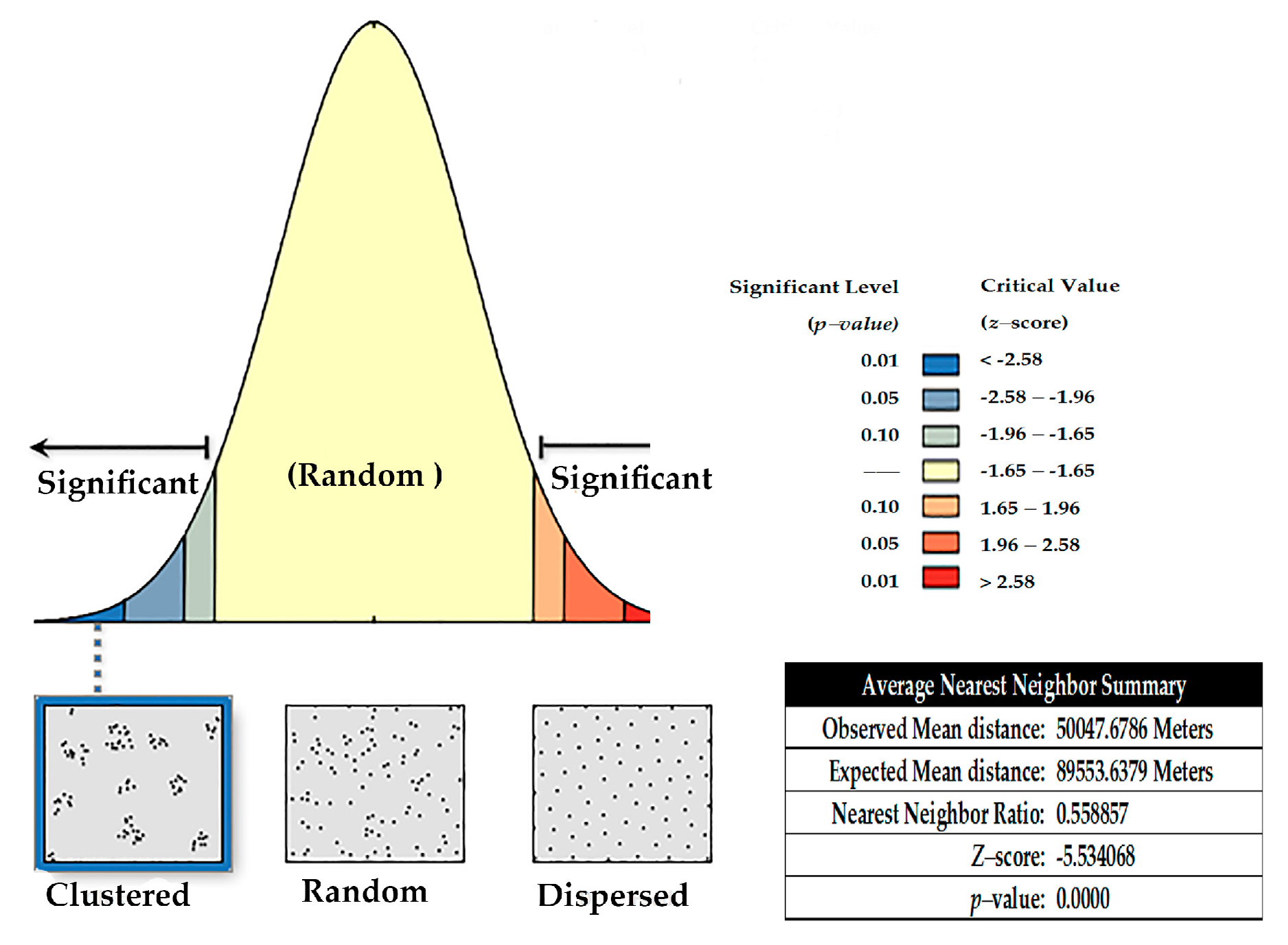
2.4. Spatial Analysis
2.4.1. Occurrence Record
2.4.2. Protozoan Occurrences Map
- i = 1, …, n are the input points. Only points that are in the radius distance of the (x,y) location come into account.
- speciesi is the species field value (in this case, the number of individual protozoan species) of point i.
- disti is the distance between the (x,y) location and point i.
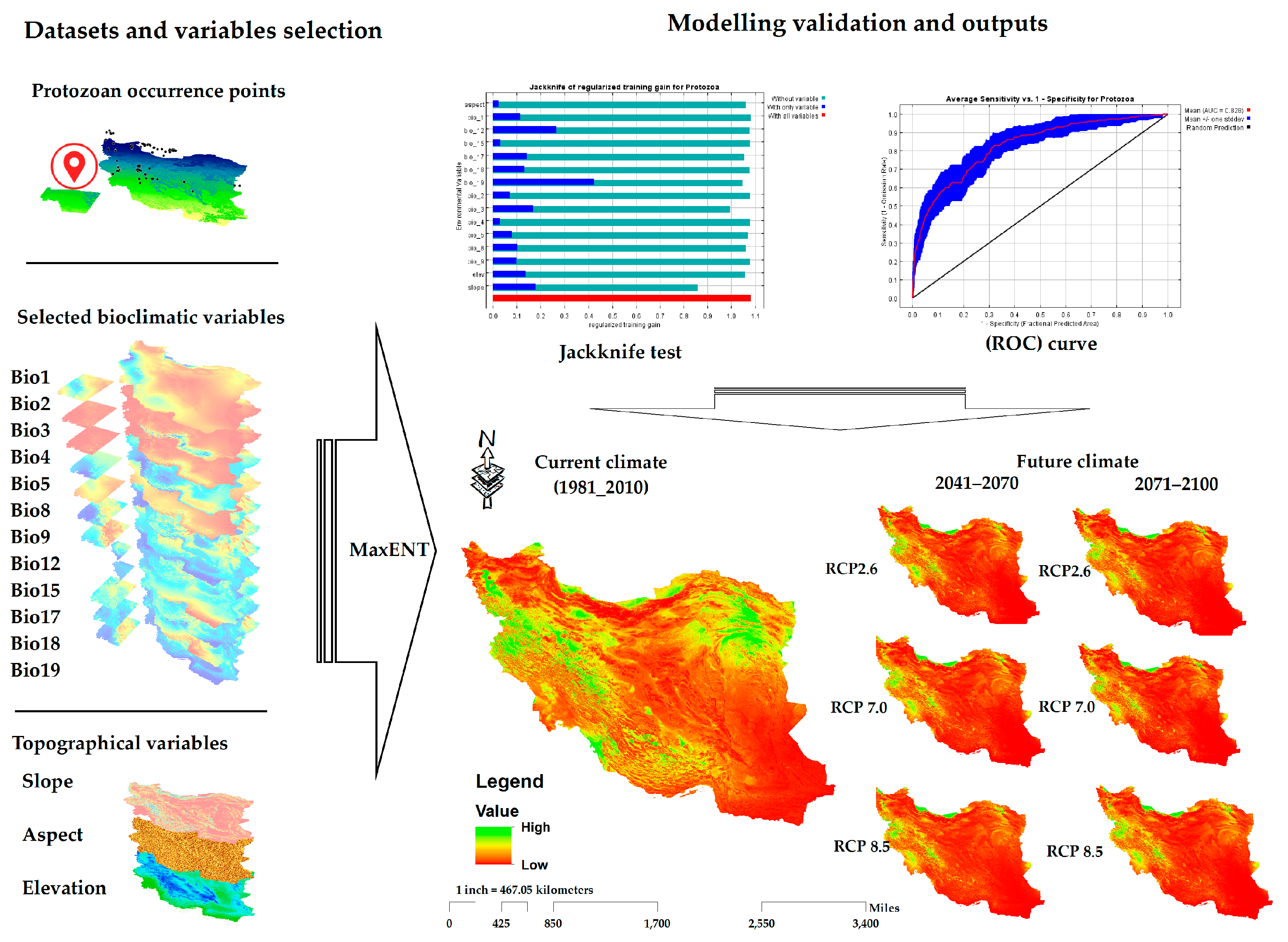
2.5. Distribution Modeling
2.5.1. Environmental Variables
2.5.2. Species Distribution Modeling (SDM) and Statistical Analysis
3. Results
3.1. Analysis of Published Reports
3.2. Spatial Analysis
Protozoan Occurrence Map
3.3. Distribution Modeling
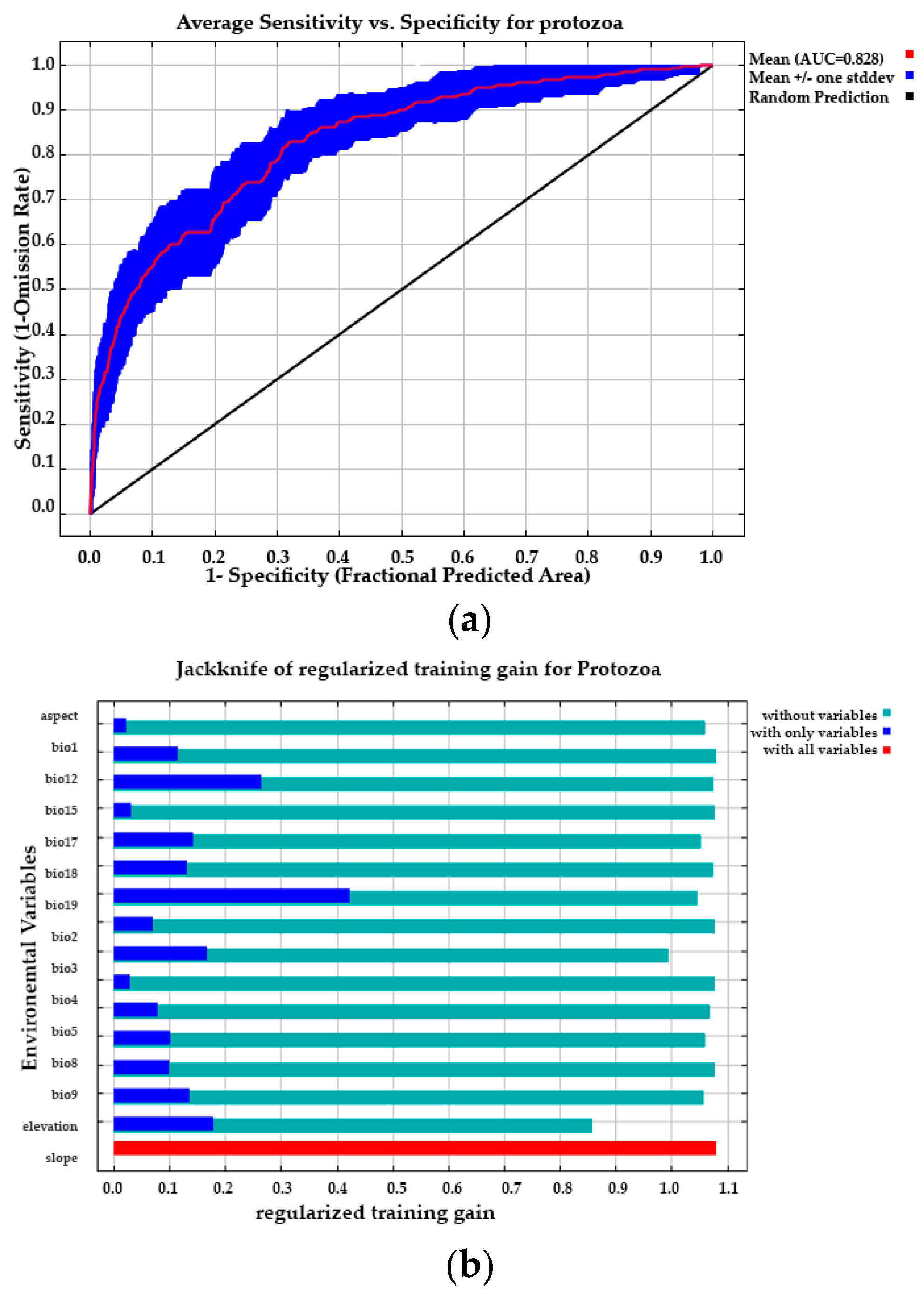
3.3.1. Model Performance and Contribution of Environmental Variables
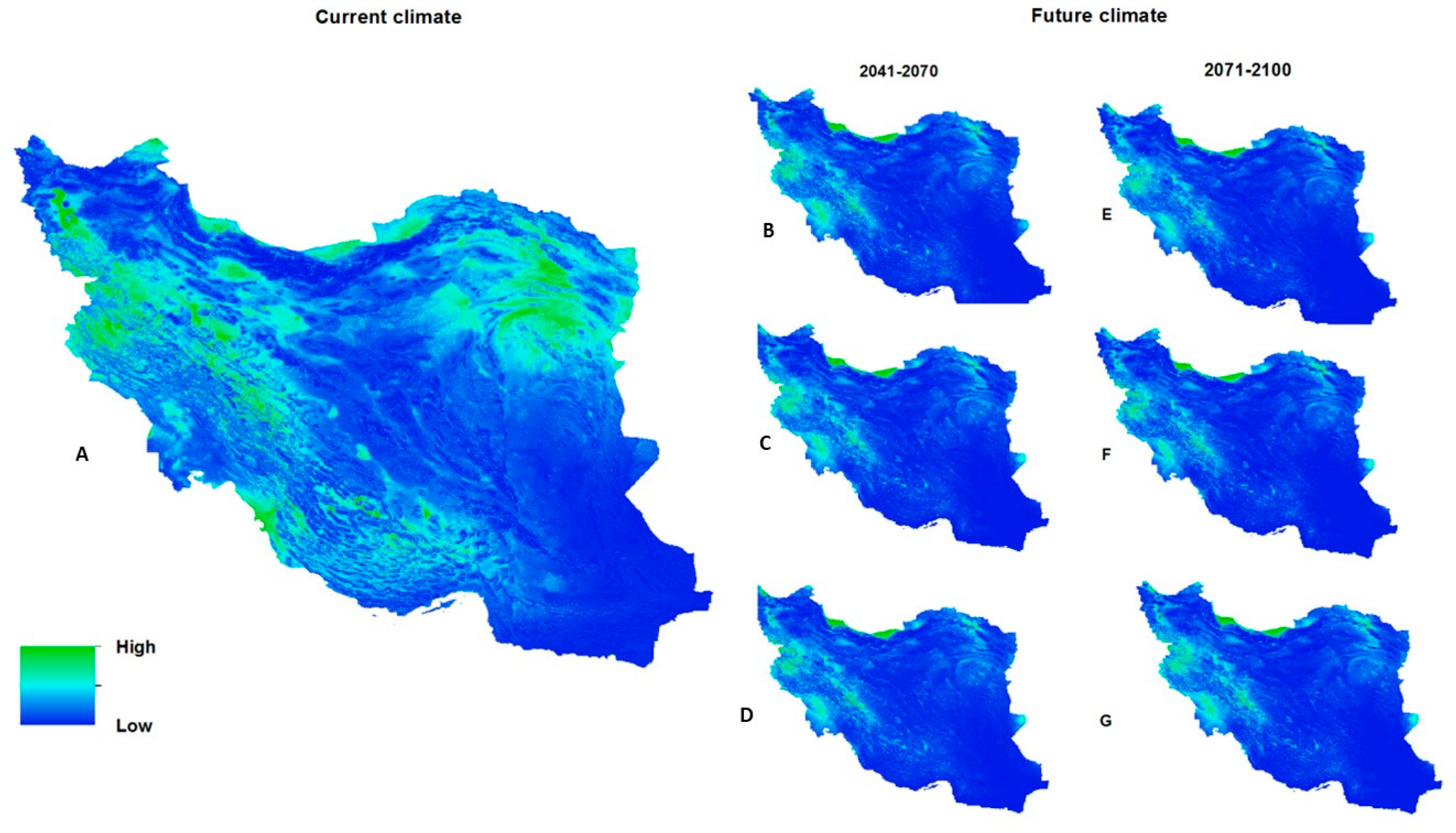
3.3.2. Predicted Current Potential Distribution
3.3.3. Future Suitable Climate Spaces
4. Discussion
4.1. Host-Parasite List
4.2. Mapping Distribution
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buchmann, K. Impact and control of protozoan parasites in maricultured fishes. Parasitology 2015, 142, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Sibley, L.D. Invasion and intracellular survival by protozoan parasites. Immunol. Rev. 2011, 240, 72–91. [Google Scholar] [CrossRef] [PubMed]
- Bermingham, M.L.; Mulcahy, M.F. Microfauna associated with amoebic gill disease in sea-farmed Atlantic salmon, Salmo salar L., smolts. J. Fish Dis. 2006, 29, 455–465. [Google Scholar] [CrossRef]
- Zilberg, D. Amoebic gill disease of marine fish caused by Neoparamoeba pemaquidensis. Acta Zool. Sin. 2005, 51, 554–556. [Google Scholar]
- Marino, F. Damage due to parasites in Mediterranean teleosts. Parassitologia 2006, 48, 19–21. [Google Scholar]
- Barzegar, M.; Bozorgnia, A.; Raissy, M. Prevalence and mean intensity of ectoparasite Infections in Caspian shemaya (Alburnus chalcoides) in southeastern Caspian Sea Basin. J. Anim. Environ. 2022, 11, 50–61. [Google Scholar]
- Jalali, B. Parasites and Parasitic Diseases in the Freshwater Fishes of Iran, 1st ed.; Iranian Fisheries Company, Aquaculture Department: Tehran, Iran, 1998; 564p. (In Persian) [Google Scholar]
- Masoumian, M.; Pazouki, J.; Yahyazadeh, M.; Teymornezhad, A. Protozoan from freshwater fishes from north west of Iran. Iran. J. Fish. Sci. 2005, 4, 31–42. [Google Scholar]
- Mokhayer, B. Survey on the parasites of Sephid-Roud River. J. Vet. Med. Fac. Tehran. Univ. 1981, 38, 61–75. [Google Scholar]
- Pazooki, J.; Masoumian, M. Synopsis of the parasites in Iranian freshwater fishes. Iran. J. Fish. Sci. 2012, 11, 570–589. [Google Scholar]
- Taghavi, M.; Mokhayer, B.; Saeedi, A.; Mosavi, H. Parasitic infection in Hemiculter lucisculus, Liza auratus and Gasterosteus aculeatus of the Zardi River (Mazandaran). Iran. Sci. Fish. J. 2013, 21, 151–156. [Google Scholar]
- Zargar, A.; Rahimi Afzal, Z.; Taheri Mirghaed, A.; Soltani, M.; Ebrahimzadeh Mousavi, H.A.; Mollaeian, H. Study of ectoparasite contamination of rainbow trout Oncorhynchus mykiss (Walbaum, 1792) in Aquatic Animal Health Research Center’s farm, Faculty of Veterinary, University of Tehran. J. Appl. Ichthyol. Res. 2017, 5, 153–166. [Google Scholar]
- Scherer, R.W.; Saldanha, I.J. How should systematic reviewers handle conference abstracts? A view from the trenches. Syst. Rev. 2019, 8, 264. [Google Scholar] [CrossRef] [PubMed]
- Lom, J.; Dykova, I. Myxozoan genera: Definition and notes on taxonomy, life-cycle terminology and pathogenic species. Folia Parasitol. 2006, 53, 1–36. [Google Scholar] [CrossRef]
- Lom, J.; Dyková, I. (Eds.) Protozoan Parasites of Fishes; Elsevier Science Publishers: Amsterdam, The Netherlands, 1992; Volume 26, 315p. [Google Scholar]
- Tawfik, G.M.; Dila, K.A.S.; Mohamed, M.Y.F.; Tam, D.N.H.; Kien, N.D.; Ahmed, A.M.; Huy, N.T. A step by step guide for conducting a systematic review and meta-analysis with simulation data. Trop. Med. Health 2019, 47, 46. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.; Griffin, L.S. The ESRI Guide to GIS Analysis; ESRI Press: Redlands, CA, USA, 2005; Volume 2, 288p. [Google Scholar]
- Silverman, B.W. Density Estimation for Statistics and Data Analysis; CRC Press: Boca Raton, FL, USA, 1986; Volume 26. [Google Scholar]
- Biler, L.; Germeç, E. Inverse Distance Weighted (IDW) Distribution Mapping of Phlebotomus papatasi, Phlebotomus tobbi, and Phlebotomus sergenti in Çukurova Region, Turkey. Commagene J. Biol. 2018, 2, 23–27. [Google Scholar] [CrossRef]
- Emadodin, I.; Reinsch, T.; Taube, F. Drought and desertification in Iran. Hydrology 2019, 6, 66. [Google Scholar] [CrossRef]
- Karger, D.N.; Saladin, B.; Wüest-Karpati, R.O.; Graham, C.H.; Zurell, D.; Mo, L.; Zimmermann, N.E. Interannual climate variability data improves niche estimates in species distribution models. bioRxiv 2021. [Google Scholar] [CrossRef]
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef]
- Boucher, O.; Servonnat, J.; Albright, A.L.; Aumont, O.; Balkanski, Y.; Bastrikov, V.; Bekki, S.; Bonnet, R.; Bony, S.; Bopp, L. Presentation and evaluation of the IPSL-CM6A-LR climate model. J. Adv. Model. Earth Syst. 2020, 12, e2019MS002010. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, Z.; Zhu, B.; Cheng, X.; Yang, L.; Gao, M.; Kong, R. MaxEnt modeling based on CMIP6 models to project potential suitable zones for Cunninghamia lanceolata in China. Forests 2021, 12, 752. [Google Scholar] [CrossRef]
- O’Neill, B.C.; Tebaldi, C.; Van Vuuren, D.P.; Eyring, V.; Friedlingstein, P.; Hurtt, G.; Knutti, R.; Kriegler, E.; Lamarque, J.-F.; Lowe, J. The scenario model intercomparison project (ScenarioMIP) for CMIP6. Geosci. Model Dev. 2016, 9, 3461–3482. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.r-project.org (accessed on 31 January 2023).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Pearson, R.G. Species’ distribution modeling for conservation educators and practitioners. Synthesis. Am. Mus. Nat. Hist. 2007, 50, 54–89. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Masomian, M.; Mehdizadeh, A.; Yahyazadeh, M. Protozoans;(Coccidia and Myxosporea;) infections in some fishes of Aras and Mahabad dams (Northwest of Iran). Iran. Sci. Fish. J. 2002, 11, 79–90. [Google Scholar]
- Molnar, K. Report of the Fish Pathology Course Held in Iran, 24 November–21 December; Fisheries Co. of Iran: Tehran, Iran, 1993. [Google Scholar]
- Pazooki, J.; Masoumian, M. Cryptobia acipenseris and Haemogregarina acipenseris infections in Acipenser guldenstadti and A. persicus in the Southern part of the Caspian Sea. J. Agric. Sci. Technol. 2004, 6, 95–101. [Google Scholar]
- Sharif Rohani, M. Survey on parasites and parasitic diseases in Sistan region. In Proceedings of the 2nd Symposium of Iranian Veterinary Clinics, Tehran, Iran, 19–21 November 1994; p. 109. [Google Scholar]
- Tavakol, S.; Motalebi, A.; Bahmani, M.; Halajian, A.; Jalali, B. Survey on gill parasites of acipenserid fishes of Caspian Sea and freshwater in Iran. Iran. Vet. Sci. 2009, 6, 763–770. [Google Scholar]
- Tavakol, S. Survey on Gill Parasites of Sturgeon Fishes (Acipenseridae) in Southern Part of Caspian Sea with Emphasis on Nitzschia sturionis Ablidgaard, 1794; Science and Research Branch, Islamic Azad University: Tehran, Iran, 2008; 115p. [Google Scholar]
- Seyed Mortezaei, R.; Pazoki, J.; Masomiay, M.; Koor, N. Identification of myxozoa and protozoa parasites of Barboid fishes of water resources in Khouzestan Province. Iran. Sci. Fish. J. 2008, 17, 63–78. [Google Scholar]
- Raissy, M.; Ansari, M. Parasites of some freshwater fish from Armand river, chaharmahal va Bakhtyari province, Iran. Iran. J. Parasitol. 2012, 7, 73–79. [Google Scholar]
- Mehdipoor, M.; Ranjbar Bahadori, S.; Masoumi, M.; Ojrati, N.; Barzegar, M.; Ghasemi, H. Identification of ectoparasites of fishes Hablerud river, Garmsar (Semnan province). J. Vet. Microbiol. Garmsar Vet. Res. J. 2010, 6, 95–100. (In Persian) [Google Scholar]
- Mehdipour, M. Parasites of Endemic and Introduced Fishes of Zaianderud River of the Esfahan; Science and Research Branch, Islamic Azad University: Tehran, Iran, 2007; 153p. [Google Scholar]
- Fadaeifard, F.; Mokhayer, B.; Ghorbani, H.; Fadaei Fard, F.; Mokhayer, B.; Ghorbani, H. Survey and identification of parasites from fishes of Choghakhor Wetland in Chaharmahal and Bakhtiari. J. Fac. Vet. Med. Univ. Tehran 2001, 56, 109–114. (In Persian) [Google Scholar]
- Raissy, M.; Azizi, H.; Fadaeifard, F.; Pour, S.Y. Parasites of some native fish from Kaaj river, Chaharmahal va Bakhtiari province, Iran. World J. Fish Mar. Sci. 2013, 5, 84–87. [Google Scholar]
- Golchin, M.A.; Ataei, A.; Sadeghi, L.R. Survey on skin and gill’s helminths and protozoa of Fahlian River’ fish. Iran. Vet. J. 2018, 14, 58–67. [Google Scholar]
- Jalali, B.; Barzegar, M.; Asadolah, S.; Mehdipour, M.; Maghsoudloo, E.; Gheshlaghi, P.; Abdolahi, F.; Mansouri, F.; Fakhri, Z. Identification of the parasites from some fishes of Zayandeh-Rud River and first record of Allocreadium laymani Bychowsky, 1962 from Iran. Vet. Res. Bulliten 2007, 4, 63–70. [Google Scholar]
- Pazouki, J.; Ghobadian, M.; Masoumian, M. Identification of some parasites of freshwater fishes of Zanjan Province, northwest Iran. Iran. Sci. Fish. J. 2005, 14, 23–40. [Google Scholar]
- Gholami, M.H.; Mokhayer, B.; Bozorgnia, A.; Hosseinzadeh Sahafi, H. Investigation on the c and intenstiy c parasitic infection of Leuciscus cephalus and Capoeta capoeta gracilis of the Neka River. J. Mar. Sci. Technol. Res. 2009, 4, 50–60. (In Persian) [Google Scholar]
- Nazari Chamak, F.; Pazooki, J.; Masoumianm, M. Identification of parasitic fauna of fishes in some water resources of Jiroft city, Kerman. In Proceedings of the 17th National & 5th International Iranian Biology Conference, Kerman, Iran, 4–6 September 2012; p. 5. [Google Scholar]
- Peyghan, R.; Razi, J.M.H.; Mohamadian, B. Study on parasitic contamination of Botak (Cyprinion macrostomum), Toeini (Barbus barbulus) and Kaputa (Capoeta trutta) in Dez River. J. Wetl. Ecobiol. 2018, 10, 15–28. [Google Scholar]
- Manshadi, A.; Masoumian, M.; Jafari, B.J.; Dowlatabadi, M.B. Protozoan and myxozoan infections in some fishes of Parishan Lake. Asian J. Anim. Vet. Adv. 2012, 7, 842–850. [Google Scholar] [CrossRef][Green Version]
- Moghainemi, S.R.; Abbasi, S.; Amiri, F. Survey on parasitic infections in the endemic fishes of Hoor-Alazim Lagoon. Iran. Fish. Res. Organ. Fish. Sci. 2004, 4, 31–42. (In Persian) [Google Scholar]
- Jalali, B.; Mahbobi Soofiani, N.; Asadollah, S.; Barzegar, M. An investigation on fish parasites in Hanna Wetland, Semirom, Isfahan Province. Iran. Sci. Fish. J. 2012, 21, 25–38. [Google Scholar]
- Rasouli, S.; Purghasem, S. Survey of external parasites infestation of cyprinidea in Zarine-rud river in West Azerbaijan province. Vet. Clin. Pathol. Q. Sci. J. 2016, 10, 155–164. [Google Scholar]
- Daghigh Roohi, J.; Sattari, M.; Nezamabadi, H.; Ghorbanpour, N. Occurrence and intensity of parasites in Prussian carp, Carassius gibelio from Anzali wetland, Southwest Caspian Sea. Iran. J. Fish. Sci. 2014, 13, 276–288. [Google Scholar]
- Raeisi, M.; Ansari, M.; Lashkari, A.; Jalali, B. Occurrence of parasites in selected fish species in Gandoman Lagoon, Iran. Iran. J. Fish. Sci. 2010, 9, 464–471. [Google Scholar]
- Raissy, M.; Fadaeifard, F.; Ansari, M.; Tajizadegan, H.; Hosseini, S.R. Study on parasites of fishes in Sooleghan Lagoon, Chaharmahal & Bakhtyari Province. Vet. J. Azad Univ. Garmsar 2009, 1, 15–19. [Google Scholar]
- Daghigh Roohi, J.; Mirhasheminasab, S.; Asgharnia, M.; Ghorbanpour, N.; Nahrvar, M.; Rufchaei, R.; Ramezani, B.; Musavi, A.; Abbasi, K.; Rastin, R. Study of Parasites Occurrence and Intensity in Fishes of Anzali Lagoon; Final Report Iranian Fisheries Science Research Institute; Iranian Fisheries Science Research Institute: Tehran, Iran, 2016; Available online: http://hdl.handle.net/1834/13770 (accessed on 21 November 2022).
- Ranjbar Bahadori, S.; Jafari Cherati, A.; Mehdipour, M.; Jalali, B. A survey on ectoparvasit in freshwater and warm water culture fishes in Mazandaran province. Vet. J. 2009, 84, 18–22. [Google Scholar]
- Ehsanfar, A.; Bozorgnia, A.; Barzegar, M.; Sadeghloo, S. Investigation on the prevalence and intensity of protozoan and metazoan parasites of some of the warm- water cultured fish species in Mazandaran Province. New Technol. Aquac. Dev. 2021, 4, 16–28. [Google Scholar]
- Bozorgnia, A.; Youssefi, M.; Barzegar, M.; Hosseinifard, S.; Ebrahimpour, S. Biodiversity of parasites of fishes in Gheshlagh (Vahdat) Reservoir, Kurdistan Province, Iran. World J. Fish Mar. Sci. 2012, 4, 249–253. [Google Scholar]
- Elahi, M.E.; Soltani, M.; Nokhbe, Z.D.; Ghafari, M.; Naderi, M. Study of parasites in skin and gill of (Schizothorax zarudnyi) in Sistan region. J. Aquat. Anim. Fish. 2014, 18, 1–11. [Google Scholar]
- Barzegar, M.; Jalali, B. Parasites of fishes of Kuftar Lake, distribution and economically importance of them. Iran. Vet. J. Shahid Chamran Ahvaz Univ. 2002, 5, 52–64. (In Persian) [Google Scholar]
- Mirghaed, A.T.; Barzegar, M.; Mousavi, H.E.; Rahmati-holasoo, H.; Bozorgnia, A. A comparative study of parasite communities of some endemic fish species in River Babol and Alborz Dam in the Southern Caspian Sea basin in Mazandaran province. Int. J. Aquat. Biol. 2017, 5, 401–407. [Google Scholar]
- Jalali, B.; Barzegar, M. Fish parasites in Zarivar lake. J. Agric. Sci. Technol. 2006, 8, 47–58. [Google Scholar]
- Keivany, Y.; Aalipour, M.; Khalagy, M.; Asaollah, S.; Siami, M. Parasitic infestation in Chondrostoma regium and Aphanius vladykovi of the Behesht Abad River (Chaharmahal-va-Bakhtiary). Iran. Sci. Fish. J. 2017, 25, 101–108. [Google Scholar]
- Barzegar, M. A survey on ectoparasite of fishes in some rivers in southwest of Caspian Basin. Ph.D. Thesis, Faculty of Veterinary Medicine, University of Tehran, Tehran, Iran, 2018; 124p. [Google Scholar]
- Mirghaed, A.T.; Mousavi, H.E.; Jazani, M.M.; Rahmati-Holasoo, H.; Bozorgnia, A. Parasite fauna of Chub, Squalius turcicus De Filippi, 1865 (Teleostei: Cyprinidae) from some rivers of the Southern Caspian Sea basin in Iran. Iran. J. Ichthyol. 2018, 5, 109–117. [Google Scholar]
- Rahanandeh, M.; Tizkar, B. Morphometric study and identification of external parasites in Hypophthalmichthys molitrix breeders in Guilan, Iran. Sustain. Aquac. Health Manag. J. 2021, 7, 1–9. [Google Scholar] [CrossRef]
- Gholami, Z.; Youssefi, M.R.; Marhaba, Z.; Alizadeh, A.; Rahimi, M.T. Aphanius sophiae (Actinoptrygii, Cyprinodontidae), a new host for Ichthyophthirius multifiliis (Ciliophora) reported from Iran. J. Parasit. Dis. 2016, 40, 1030–1032. [Google Scholar] [CrossRef][Green Version]
- Raissy, M.; Pirali Kheirabadi, E.; Mohseni, P. Identification of parasites of Aphanius vladykovi from Shalamzar and Salm Lakes, Chaharmahal va Bakhtiari Province, Iran. Iran. Sci. Fish. J. 2018, 27, 159–163. [Google Scholar]
- Mirhashemi Nasab, S.; Firouzbakhsh, F.; Sattari, M.; Ghasemi, M. Prevalence and intensity of parasites in pike (Esox lucius) from Anzali Wetland and evaluation of their impacts on biometric characticties in host fish. Iran. Sci. Fish. J. 2019, 1, 69–87. [Google Scholar]
- Barzegar, M.; Raeisi, M.; Bozorgnia, A.; Jalali, B. Parasites of the eyes of fresh and brackish water fishes in Iran. Iran. J. Vet. Res. 2008, 9, 256–261. [Google Scholar]
- Miariar, A.; Bozorgnia, A.; Pazoki, J.; Barzegar, M.; Masomian, M.; Jalali, B. Fish parasites in Valasht Lake and Chalns River. Iran. Sci. Fish. J. 2008, 17, 133–138. [Google Scholar]
- Mehdizadeh Mood, S.; Shohreh, P.; Sahandi, J. A survey on ectoparasite fauna of cold water fish farms in Mazandaran Province, Iran. Hum. Vet. Med. 2011, 3, 246–251. [Google Scholar]
- Azadikhah, D. Survey of Parasitic Infection of Sander lucioperca (Linnaeus, 1758) and Silurus glanis (Linnaeus, 1758) in Aras Reservoir with Emphasizes of Monogenean Parasites; Science and Research Branch, Islamic Azad University: Tehran, Iran, 2008; 155p. [Google Scholar]
- Pazooki, J.; Goorabzarmakhi, F.T.; Masoumian, M. Parasitic infection of an endemic fish (Blicca bjoerkna) and an exotic fish (Hemiculter beucisculus) in Anzali Lagoon, Caspian Sea, Iran. Iran. J. Parasitol. 2011, 6, 66–73. [Google Scholar] [PubMed]
- Bazari Moghaddam, S.; Mokhayer, B.; Masoumian, M.; Shenavar Masouleh, A.; Jalilpour, J.; Masoumzadeh, M.; Alizadeh, M. Parasitic infection among larvae and fingerlings of the Persian sturgeon (Acipenser persicus) in Vniro tanks and earthen ponds. Iran. J. Fish. Sci. 2010, 9, 342–351. [Google Scholar]
- Mirnategh, B.; Shabanipour, N.; Sattari, M. Occurrence and intensity of parasites in Chelon aurata (Risso, 1810) and Neogobius caspius (Eichwald, 1831) (Teleostei: Perciformes) from southern Caspian Sea. Int. J. Aquat. Biol. 2017, 5, 310–320. [Google Scholar]
- Khojasateh-Khah, M. Survey on Protozoan Parasites Infections of Cultured Trout in Chaharmahal and Bakhtiari Province. Ph.D. Thesis, Faculty of Veterinary Medicine, University of Tehran, Tehran, Iran, 1999; 87p. (In Persian). [Google Scholar]
- Rahanandeh, M.; Tizkar, B.; Abedi, M. Identification of Chalcalburnus chalcoides internal parasites in Sefidrood river, Guilan, Iran. Iran. J. Aquat. Anim. Health 2021, 7, 19–29. [Google Scholar] [CrossRef]
- Khara, H.; Nezami, A.; Satari, M.; Mousavi, A.; Mousapour, M.; Hajipour, A. Occuranee and intensity of parasites from perch (Perca fluviatilis, L. 1785) in Amirkelaieh wetland of Lahijan (Caspian basin-Iran). Pajouhesh Sazandegi 2005, 67, 92–103. [Google Scholar]
- Esmaeili, H.R.; Sayyadzadeh, G.; Eagderi, S.; Abbasi, K. Checklist of Freshwater Fishes of Iran. FishTaxa 2018, 3, 1–95. [Google Scholar]
- Jalali, B.; Molnar, K. Occurrence of monogeneans on fresh-water fishes in Iran—Dactylogyrus spp. on cultured Iranian fishes. Acta Vet. Hung. 1990, 38, 239–242. [Google Scholar]
- Didier, E.S.; Weiss, L.M. Microsporidiosis: Not just in AIDS patients. Curr. Opin. Infect. Dis. 2011, 24, 490–495. [Google Scholar] [CrossRef]
- Lom, J.; Dykova, I. Developments in Aquaculture and Fisheries. In Protozoan Parasites of Fishes, 1st ed.; Elsevier Science Publishers: Amsterdam, The Netherlands, 1992; p. 316. [Google Scholar]
- Jouladeh-Roudbar, A.; Eagderi, S.; Ghanavi, H.; Doadrio, I. A new species of the genus Capoeta Valenciennes, 1842 from the Caspian Sea basin in Iran (Teleostei, Cyprinidae). ZooKeys 2017, 682, 137–155. [Google Scholar] [CrossRef]
- Tange, E.Ø.; Mathiessen, H.; von Gersdorff Jørgensen, L. Effects of pH on free-living stages of a Nordic strain of the economically important freshwater fish parasite Ichthyophthirius multifiliis. Int. J. Parasitol. 2020, 50, 859–864. [Google Scholar] [CrossRef]
- Rohde, K. Marine Parasitology; Csiro Publishing: Clayton, Australia, 2005; p. 565. [Google Scholar]
- Fourcade, Y.; WallisDeVries, M.F.; Kuussaari, M.; van Swaay, C.A.; Heliölä, J.; Öckinger, E. Habitat amount and distribution modify community dynamics under climate change. Ecol. Lett. 2021, 24, 950–957. [Google Scholar] [CrossRef]
- Kołodziej-Sobocińska, M. Factors affecting the spread of parasites in populations of wild European terrestrial mammals. Mammal. Res. 2019, 64, 301–318. [Google Scholar] [CrossRef]
- Iyaji, F.; Etim, L.; Eyo, J. Parasite assemblages in fish hosts. Bio Res. 2009, 7, 570–581. [Google Scholar] [CrossRef]
- Esmaeili, H.R.; Gholamhosseini, A.; Mohammadian-Kalat, T.; Aliabadian, M. Predicted changes in climatic niche of Alburnus species (Teleostei: Cyprinidae) in Iran until 2050. Turk. J. Fish. Aquat. Sci. 2018, 18, 995–1003. [Google Scholar] [CrossRef]
- Vidal-Martínez, V.M.; Velázquez-Abunader, I.; Centeno-Chalé, O.A.; May-Tec, A.L.; Soler-Jiménez, L.C.; Pech, D.; Mariño-Tapia, I.; Enriquez, C.; Zapata-Pérez, O.; Herrera-Silveira, J. Metazoan parasite infracommunities of the dusky flounder (Syacium papillosum) as bioindicators of environmental conditions in the continental shelf of the Yucatan Peninsula, Mexico. Parasites Vectors 2019, 12, 277. [Google Scholar] [CrossRef]
- Kuhn, T.; Cunze, S.; Kochmann, J.; Klimpel, S. Environmental variables and definitive host distribution: A habitat suitability modelling for endohelminth parasites in the marine realm. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef]
- Moore, C.; Drazen, J.C.; Radford, B.T.; Kelley, C.; Newman, S.J. Improving essential fish habitat designation to support sustainable ecosystem-based fisheries management. Mar. Policy 2016, 69, 32–41. [Google Scholar] [CrossRef]
- Yousefi, M.; Kafash, A.; Khani, A.; Nabati, N. Applying species distribution models in public health research by predicting snakebite risk using venomous snakes’ habitat suitability as an indicating factor. Sci. Rep. 2020, 10, 18073. [Google Scholar] [CrossRef]
- Kim, Z.; Shim, T.; Ki, S.J.; An, K.-G.; Jung, J. Prediction of three-dimensional shift in the distribution of largemouth bass (Micropterus salmoides) under climate change in South Korea. Ecol. Indic. 2022, 137, 108731. [Google Scholar] [CrossRef]
- Alt, K.G.; Kochmann, J.; Klimpel, S.; Cunze, S. Improving species distribution models of zoonotic marine parasites. Sci. Rep. 2019, 9, 9851. [Google Scholar] [CrossRef]
- Kwon, Y.-S.; Bae, M.-J.; Hwang, S.-J.; Kim, S.-H.; Park, Y.-S. Predicting potential impacts of climate change on freshwater fish in Korea. Ecol. Inform. 2015, 29, 156–165. [Google Scholar] [CrossRef]
- Pandit, S.N.; Maitland, B.M.; Pandit, L.K.; Poesch, M.S.; Enders, E.C. Climate change risks, extinction debt, and conservation implications for a threatened freshwater fish: Carmine shiner (Notropis percobromus). Sci. Total Environ. 2017, 598, 1–11. [Google Scholar] [CrossRef]
- Lymbery, A.J.; Lymbery, S.J.; Beatty, S.J. Fish out of water: Aquatic parasites in a drying world. Int. J. Parasitol. Parasites Wildl. 2020, 12, 300–307. [Google Scholar] [CrossRef]
- Akoll, P.; Konecny, R.; Mwanja, W.W.; Nattabi, J.K.; Agoe, C.; Schiemer, F. Parasite fauna of farmed Nile tilapia (Oreochromis niloticus) and African catfish (Clarias gariepinus) in Uganda. Parasitol. Res. 2012, 110, 315–323. [Google Scholar] [CrossRef]
- Cao, Q.; Yu, D.; Georgescu, M.; Han, Z.; Wu, J. Impacts of land use and land cover change on regional climate: A case study in the agro-pastoral transitional zone of China. Environ. Res. Lett. 2015, 10, 124025. [Google Scholar] [CrossRef]
- Rahimi-Moghaddam, S.; Kambouzia, J.; Deihimfard, R. Adaptation strategies to lessen negative impact of climate change on grain maize under hot climatic conditions: A model-based assessment. Agric. For. Meteorol. 2018, 253, 1–14. [Google Scholar] [CrossRef]
- Burgin, L.; Ekström, M.; Dessai, S. Combining dispersion modelling with synoptic patterns to understand the wind-borne transport into the UK of the bluetongue disease vector. Int. J. Biometeorol. 2017, 61, 1233–1245. [Google Scholar] [CrossRef]
- Lin, Y.; Feng, Z.; Wu, W.; Yang, Y.; Zhou, Y.; Xu, C. Potential impacts of climate change and adaptation on maize in northeast China. Agron. J. 2017, 109, 1476–1490. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Climate Type | IDM Values |
|---|---|
| Arid | IDM < 10 |
| Semi-arid | 10 ≤ IDM < 20 |
| Mediterranean | 20 ≤ IDM < 24 |
| Semi-humid | 24 ≤ IDM < 28 |
| Humid | 28 ≤ IDM < 35 |
| Very humid | 35 ≤ IDM < 55 |
| Categories | Name of Variables | Unit |
|---|---|---|
| Bioclimatic variables | Annual mean temperature (BIO1) | °C |
| Mean diurnal range (mean of monthly max temp–min temp) (BIO2) | °C | |
| Isothermality (BIO2/BIO7) (×100) (BIO3) | °C | |
| Temperature seasonality (standard deviation ×100) (BIO4) | °C | |
| Max temperature of warmest month (BIO5) | °C | |
| Mean temperature of wettest quarter (BIO8) | °C | |
| Mean temperature of driest quarter (BIO9) | °C | |
| Annual precipitation (BIO12) | mm | |
| Precipitation seasonality (coefficient of variation) (BIO15) | mm | |
| Precipitation of driest quarter (BIO17) | mm | |
| Precipitation of warmest quarter (BIO18) | mm | |
| Precipitation of coldest quarter (BIO19) | mm | |
| Topography variables | Elevation (DEM) | m |
| Slope | % | |
| Aspect (Asp) | Degrees |
| Parasite | Host Taxonomy | Host Name | Infected Organ | Environment, Type | Basin | Locality | Ref. |
|---|---|---|---|---|---|---|---|
| Ph: Apicomplexa Cl: Conoidasida Or: Eucoccidiorida Fa: Eimeriidae | |||||||
| Goussia carpelli (Leger and Stankovich, 1921) Dykova and Lom, 1983 | OR: Cypriniformes FA: Cyprinidae | Cyprinus carpio | Intestine | FW, Re, Ri | Caspian | Aras, SefidRood | [31,32] |
| Goussia sinensis Chen, 1956 | OR: Cypriniformes FA: Xenocyprididae | Hypophthalmichthys molitrix | Intestine | FW, Ri | Caspian | SefidRood | [32] |
| Ph: Apicomplexa Cl: Conoidasida Or: Eucoccidiorida Fa: Haemogregarinidae | |||||||
| Haemogregarina acipenseris Navrotskii, 1914 | OR: Acipenseriformes FA: Acipenseridae | Acipenser gueldenstaedtii | Blood | BW, S | Caspian | Caspian | [33] |
| Acipenser persicus | Blood | BW, S | Caspian | Caspian | [33] | ||
| Ph: Choanozoa Cl: Ichthyosporea Or: Dermocystida Fa: Dermocystidae | |||||||
| Dermocystidium salmonis Davis 1947 | OR: Salmoniformes FA: Salmonidae | Oncorhynchus mykiss | Gills | FW, Fa | Tigris | Chaharmahal and Bakhtiari | [10] |
| Ph: Ciliophora Cl: Litostomatea Or: Pleurostomatida Fa: Amphileptidae | |||||||
| Amphileptus branchiarum Weinrich, 1924 | OR: Cypriniformes FA: Cyprinidae | Capoeta capoeta | Skin, fins | FW, Ri | Urmia | Zangmar | [8] |
| Ph: Ciliophora Cl: Litostomatea Or: Vestibuliferida Fa: Balantidiidae | |||||||
| Balantidium ctenopharyngodoni Chen, 1955 | OR: Cypriniformes FA: Xenocyprididae | Ctenopharyngodon idella | Intestine | FW, L | Sistan | Hamun | [34] |
| Ph: Ciliophora Cl: Oligohymenophorea Or: Hymenostomatida Fa: Ichthyophthiriidae | |||||||
| Ichthyophthirius multifiliis Fouquet, 1876 | OR: Acipenseriformes FA: Acipenseridae | Acipenser persicus | Gills | FW, Fa | Caspian | International Sturgeon Fa | [35] |
| OR: Acipenseriformes FA: Acipenseridae | Acipenser stellatus | Gills | FW, Fa | Caspian | International Sturgeon Fa | [35] | |
| OR: Acipenseriformes FA: Acipenseridae | Huso huso | Gills | FW, Fa | Caspian | International Sturgeon Fa | [36] | |
| OR: Cypriniformes FA: Cyprinidae | Arabibarbus grypus | Skin, gills | FW, Ri, W | Karun; Tigris | Karun, Karkheh, Shadegan; Armand | [37,38] | |
| Barbus lacerta | gills | FW, Ri | Kavir | HablehRood | [39] | ||
| Capoeta aculeata | Gills, skin | FW, Ri, W | Isfahan; Tigris; Kavir | ZayandehRood; Armand, Choghakhor; Kaaj; HablehRood | [38,39,40,41,42] | ||
| Capoeta barroisi | Gills, skin | FW, Ri | Kor | Fahlian | [43] | ||
| Capoeta capoeta | Gills, skin | FW, Ri | Caspian; Isfahan; Urmia | SefidRood, NekaRood, SojasRood; ZayandehRood; Sarysou, Zangmar | [8,9,44,45,46] | ||
| Capoeta damascina | Gills, skin | FW, Q, Ri, W | Kerman- Nain; Isfahan; Tigris, Urmia; Kavir | Jafar abad, Konaroiyeh; ZayandehRood; Armand, Kaaj; Choghakhor; ZarinehRood; HablehRood | [7,38,39,40,41,42,44,47] | ||
| Capoeta trutta | Gills, skin | FW, Ri | Karun | Dez | [48] | ||
| Carasobarbus luteus | Gills, skin | FW, L, W | Kor; Karun | Parishan; HoorAlazim | [49,50] | ||
| Carassius carassius | Gills, skin | FW, L, Re | Kor; Urmia | Parishan; Aras | [8,49] | ||
| Carassius auratus | Gills, skin | FW, Ri, W | Tigris; Caspian; Isfahan | Choghakhor; SefidRood; Hanna | [9,41,51] | ||
| Carassius gibelio | Gills, skin | FW, Ri, Lg, W | Urmia; Caspian; Tigris | ZarrinehRood; Anzali, SefidRood; Gandoman, | [9,52,53,54,55] | ||
| Sooleghan | |||||||
| Cyprinion macrostomum | Gills, skin | FW, Ri | Karun; Kor | Dez; Fahlian | [43,48] | ||
| Cyprinus carpio | Gills, skin, fin | FW, Ri, Re, Fa, Lg, W, L | Urmia; Caspian; Karun; Tigris; Kor; Isfahan | ZarrinehRood; Mazandaran, Anzali, Dashte Naz, SefidRood; HoorAlazim, Sooleghan, Vahdat, Gandoman; Parishan; ZayandehRood | [9,40,49,50,52,54,55,56,57,58,59] | ||
| Luciobarbus barbulus | Gills, skin | FW, Ri | Karun; Tigris | Dez; Armand | [38,48] | ||
| Luciobarbus capito | Skin | FW, Ri | Caspian | Aras | [8] | ||
| Luciobarbus esocinus | Skin | FW, Ri, W | Karun | Karun, Karkheh, Shadegan | [37] | ||
| Mesopotamichthys sharpeyi | Gills, skin | FW, Ri, W | Karun | Karun, Karkheh, Shadegan, HoorAlazim | [37,50] | ||
| Schizocypris altidorsalis | Skin | FW, L | Sistan | Hamun | [34] | ||
| Schizothorax pelzami | Skin | FW, L | Sistan | Hamun | [34] | ||
| Schizothorax zarudnyi | Gills, skin | FW, L, Fa | Sistan | Hamun, Zahak | [34,60] | ||
| OR: Cypriniformes FA: Leuciscidae | Abramis brama | Skin | FW, Re | Urmia | Aras | [8] | |
| Acanthobrama persidis | Gills | FW, L | Kor | Kuftar | [61] | ||
| Alburnoides eichwaldii | Skin | FW, Ri | Caspian | Aras | [8] | ||
| Alburnoides tabarestanensis | Gills | FW, Re, Ri | Caspian | Alborz, BabolRood | [62] | ||
| Alburnus chalcoides | Gills | FW, Ri | Isfahan; Caspian | ZayandehRood; Cheshmeh Kileh, ShiRood | [6,40] | ||
| Alburnus hohenackeri | Gills, skin | FW, L | Tigris | Zarivar | [63] | ||
| Alburnus mossulensis | Gills, skin | FW, Ri | Kor | Fahlian | [43] | ||
| Chondrostoma orientale | Gills, skin | FW, L, W | Kor; Tigris | Kuftar; Choghakhor | [41,61] | ||
| Chondrostoma regium | Gills, skin | FW, Ri | Tigris; Isfahan | Kaaj, Behesht Abad; ZayandehRood | [42,44,64] | ||
| Leuciscus vorax | Gills, skin | FW, W | Karun | HoorAlazim | [50] | ||
| Squalius cephalus | Gills | FW, Ri | Urmia; Caspian | ZarinehRood; | [7] | ||
| NekaRood, Chalus, Tajan, ShiRood, SiahRood | [46,65,66] | ||||||
| Vimba vimba | Gills | FW, Ri | Caspian | Cheshmeh Kileh | [65] | ||
| OR: Cypriniformes FA: Xenocyprididae | Ctenopharyngodon idella | Gills | FW, Ri | Isfahan; Caspian; Urmia | ZayandehRood; SefidRood; ZarrinehRood | [9,40,52] | |
| Hemiculter leucisculus | not stated | FW, Lg | Caspian | Anzali | [56] | ||
| Hypophthalmichthys molitrix | Gills, skin, fin | FW, Fa, Ri | Caspian | Gilan, SefidRood, Mazandaran | [9,57,67] | ||
| Hypophthalmichthys nobilis | Skin, fin | FW, Fa | Caspian | Mazandaran | [57] | ||
| OR: Cyprinodontiformes FA: Aphaniidae | Aphanius sophiae | Gills, skin | FW, Sp | Kor | Safashahr | [68] | |
| Aphanius vladykovi | Gills, skin | FW, Ri, L | Isfahan; Tigris | ZayandehRood; Behesht Abad, Shalamzar, Salm | [40,64,69] | ||
| OR: Esociformes FA: Esocidae | Esox lucius | Gills | FW, Lg, Ri | Caspian | Anzali, ShiRood | [56,65,70] | |
| OR: Mugiliformes FA: Mugilidae | Chelon auratus | Gills, skin | FW, Ri | Caspian | Zardi | [11] | |
| Planiliza abu | Gills, skin | FW, W | Karun | HoorAlazim | [50] | ||
| OR: Perciformes FA: Gasterosteidae | Gasterosteus aculeatus | Gills, skin | FW, Ri | Caspian | Zardi | [11] | |
| OR: Salmoniformes FA: Salmonidae | Oncorhynchus mykiss | Surface of eyes, gills, skin, fin | FW, Ri, Fa | Caspian | Haraz, Mazandaran, Chalus | [10,57,71,72,73] | |
| OR: Siluriformes FA: Sisoridae | Glyptothorax silviae | Gills, skin | FW, Ri | Tigris | Armand | [38] | |
| Silurus glanis | Gills, skin | FW, Lg, Re | Caspian; Urmia | Anzali; Aras | [60,74] | ||
| OR: Synbranchiformes FA: Mastacembelidae | Mastacembelus mastacembelus | Gills | FW, L | Tigris | Zarivar | [63] | |
| Ph: Ciliophora Cl: Oligohymenophorea Or: Hymenostomatida Fa: Tetrahymenidae | |||||||
| Tetrahymena pyriformis Ehrenberg, 1830 | OR: Cypriniformes FA: Cyprinidae | Capoeta capoeta | Skin | FW, Ri | Urmia | Zangmar | [8] |
| Ph: Ciliophora Cl: Oligohymenophorea Or: Mobilida Fa: Trichodinidae | |||||||
| Trichodina domerguei Wallengren, 1897 | OR: Cypriniformes FA: Cyprinidae | Barbus lacerta | Gills, skin | FW, Ri | Caspian | SefidRood | [9] |
| Capoeta capoeta | Gills, skin | FW, Ri | Caspian | SefidRood | [9] | ||
| Cyprinus carpio | Gills, skin | FW, Ri | Caspian | SefidRood | [9] | ||
| Luciobarbus capito | Gills, skin | FW, Ri | Caspian | SefidRood | [9] | ||
| OR: Cypriniformes FA: Leuciscidae | Abramis brama | Gills, skin | FW, Ri | Caspian | SefidRood | [9] | |
| Alburnoides eichwaldii | Gills, skin | FW, Ri | Caspian | SefidRood | [9] | ||
| Trichodina gracilis Polyanski, 1995 | OR: Cypriniformes FA: Cyprinidae | Capoeta razii | Gills | FW, Ri | Caspian | BabolRood | [62] |
| Trichodina perforata Lom, Golemansky and Grupcheva, 1976 | OR: Cypriniformes FA: Cyprinidae | Barbus lacerta | Gills, skin | FW, Ri | Urmia | SojasRood | [45] |
| Capoeta capoeta | Gills, skin | FW, Ri, Re | Urmia | Zangmar, Ghezel Ozan, SojasRood, Hasan Abdaal | [8,45] | ||
| Carassius auratus | Gills, skin, fins | FW, Re | Isfahan | Hanna | [51] | ||
| Luciobarbus capito | Gills, skin | FW, Ri | Urmia | Aras, Sarysou | [8] | ||
| OR: Cypriniformes FA: Leuciscidae | Abramis brama | Gills, skin | FW, Re | Urmia | Aras | [8] | |
| Alburnoides eichwaldii | Gills, skin | FW, Ri | Urmia | Aras, Sarysou | [8,45] | ||
| Alburnus filippii | Skin | FW, Ri | Urmia | SojasRood | [45] | ||
| Blicca bjoerkna | Gills, skin, fins | FW, Lg | Caspian | Anzali | [75] | ||
| OR: Cypriniformes FA: Xenocyprididae | Hemiculter leucisculus | Gills, skin, fins | FW, Lg | Caspian | Anzali | [75] | |
| OR: Perciformes FA: Percidae | Sander lucioperca | Gills, skin | FW, Re | Urmia | Aras | [8] | |
| OR: Siluriformes FA: Siluridae | Siluris glanis | Gills, skin | FW, Re | Urmia | Aras | [8] | |
| Trichodina pediculus Ehrenberg, 1831 | OR: Cypriniformes FA: Cyprinidae | Capoeta damascina | Gills | FW, L | Tigris | Zarivar | [63] |
| Cyprinus carpio | Gills | FW, L | Tigris | Zarivar | [63] | ||
| OR: Synbranchiformes FA: Mastacembelidae | Mastacembelus mastacembelus | Gills | FW, L | Tigris | Zarivar | [63] | |
| Trichodina truttae Mueller, 1937 | OR: Salmoniformes FA: Salmonidae | Oncorhynchus mykiss | Skin, fins | FW, Ri | Caspian | Chalus | [72] |
| Trichodina reticulata Hirschmann and Partsch, 1955 | OR: Acipenseriformes FA: Acipenseridae | Acipenser gueldenstaedtii | not stated | FW, Fa | Caspian | Shahid Beheshty | [76] |
| Acipenser persicus | not stated | FW, Fa | Caspian | Shahid Beheshty | [76] | ||
| Acipenser stellatus | not stated | FW, Fa | Caspian | Shahid Beheshty | [76] | ||
| OR: Mugiliformes FA: Mugilidae | Chelon auratus | not stated | BW, S | Caspian | Kiashahr, Anzali, Chamkhaleh | [77] | |
| Trichodinella subtilis Lom, 1959 | OR: Cypriniformes FA: Cyprinidae | Cyprinus carpio | Skin | FW, Fa | Urmia | West Azerbaijan | [10] |
| Tripartiella lata Lom 1963 | OR: Salmoniformes FA: Salmonidae | Oncorhynchus mykiss | Skin | FW, Fa | Urmia | West Azerbaijan | [10] |
| Ph: Ciliophora Cl: Phyllopharyngea Or: Chlamydodontida Fa: Chilodonellidae | |||||||
| Chilodonella cyprini (Moroff, 1902) Strand, 1928 | OR: Cyprinodontiformes FA: Aphaniidae | Squalius cephalus | Skin | FW, Ri | Caspian | Chalus | [72] |
| OR: Salmoniformes FA: Salmonidae | Oncorhynchus mykiss | Skin | FW, Fa, Ri | Tigris; Caspian | Chaharmahal and Bakhtiari; Chalus | [10,72] | |
| Chilodonella piscicola (Zacharias 1894) Jankowski 1980 | OR: Cypriniformes FA: Cyprinidae | Capoeta capoeta | Gills, skin | FW, Ri | Urmia | Ghezel Ozan | [45] |
| OR: Cypriniformes FA: Xenocyprididae | Hypophthalmichthys molitrix | Gills, skin, fins | FW, Fa | Caspian | Gilan | [67] | |
| Ph: Ciliophora Cl: Spirotrichea Or: Sporadotrichida Fa: Oxytrichidae | |||||||
| Stylonychia pustulata (Müller, 1786) Ehrenberg, 1835 | OR: Cypriniformes FA: Cyprinidae | Capoeta capoeta | Gills | FW, Ri | Urmia | Zangmar | [8] |
| Ph: Euglenozoa Cl: Kinetoplastea Or: Ichthyobodonidae Fa: Ichthyobodonidae | |||||||
| Ichthyobodo necator Henneguy, 1928 | OR: Cypriniformes FA: Cyprinidae | Arabibarbus grypus | Skin | FW, W | Karun | HoorAlazim | [7] |
| Capoeta capoeta | Gills | FW, Ri, Re | Urmia; Caspian | ZarinehRood; Sohreyn | [7,45] | ||
| Carassius auratus | Skin | FW, Lg | Caspian | Anzali | [10] | ||
| OR: Cypriniformes FA: Leuciscidae | Leuciscus vorax | Gills | FW, W | Karun | HoorAlazim | [7] | |
| OR: Cyprinodontiformes FA: Aphaniidae | Aphanius vladykovi | Skin | FW, L | Tigris | Shalamzar | [69] | |
| OR: Cypriniformes FA: Xenocyprididae | Hemiculter leucisculus | Skin | FW, Ri | Caspian | Zardi | [11] | |
| OR: Mugiliformes FA: Mugilidae | Chelon auratus | Skin | FW, Ri | Caspian | Zardi | [11] | |
| Planiliza abu | Skin, gills | FW, W, Ri | Karun | HoorAlazim, Karun | [7] | ||
| Ph: Euglenozoa Cl: Kinetoplastea Or: Parabodonida Fa: Cryptobiaceae | |||||||
| Cryptobia branchialis Nie in Chen, 1956 | OR: Cypriniformes FA: Xenocyprididae | Hypophthalmichthys molitrix | Gills, skin, fins | FW, Fa | Karun; Caspian | Khouzestan; Guilan | [7,67] |
| Ph: Euglenozoa Cl: Kinetoplastea Or: Parabodonida Fa: Cryptobiaceae | |||||||
| Trypanoplasma acipenseris Ioff, Lewashow, Boschenko, 1926 | OR: Acipenseriformes FA: Acipenseridae | Acipenser gueldenstaedtii | Blood | BW, S | Caspian | Caspian | [33] |
| OR: Acipenseriformes FA: Acipenseridae | Acipenser persicus | Blood | BW, S | Caspian | Caspian | [33] | |
| Trypanoplasma borelli Laveran et Mesnil, 1901 | OR: Salmoniformes FA: Salmonidae | Oncorhynchus mykiss | Blood | FW, Fa | Tigris | Chaharmahal and Bakhtiari | [78] |
| Ph: Euglenozoa Cl: Kinetoplastea Or: Trypanosomatida Fa: Trypanosomatidae | |||||||
| Trypanosoma percae Brumpt, 1906 | OR: Cypriniformes FA: Leuciscidae | Alburnus chalcoides | Blood | FW, Ri | Caspian | Sefidrood | [79] |
| OR: Perciformes FA: Percidae | Perca fluviatilis | Blood | FW, W, S | Caspian | Amirkelayeh, Caspian | [7,80] | |
| Ph: Fornicata Cl: Trepomonadea Or: Diplomonadida Fa: Hexamitidae | |||||||
| Hexamita salmonis (Moore, 1923) Wenyon, 1926 | OR: Salmoniformes FA: Salmonidae | Oncorhynchus mykiss | Intestine | FW, Fa | Tigris; Urmia | Chaharmahal and Bakhtiari; West Azerbaijan | [10,78] |
| Ph: Tubulinea Cl: Elardia Or: Arcellinida Fa: Arcellidae | |||||||
| Arcella vulgaris Ehrenberg, 1830 | OR: Cypriniformes FA: Cyprinidae | Capoeta capoeta | Skin | FW, Ri | Urmia | Zangmar | [8] |
| Host | Parasite Species |
|---|---|
| Class Actinopterygii | |
| Order Acipenseriformes | |
| Family Acipenseridae Bonaparte, 1831 | |
| Genus Acipenser Linnaeus, 1758 | |
| Species Acipenser gueldenstaedtii Brandt and Ratzeburg, 1833 | Haemogregarina acipenseris |
| Trichodina reticulata | |
| Trypanoplasma acipenseris | |
| Species Acipenser persicus Borodin, 1897 | Haemogregarina acipenseris |
| Ichthyophthirius multifiliis | |
| Trichodina reticulata | |
| Trypanoplasma acipenseris | |
| Species Acipenser stellatus Pallas, 1771 | Ichthyophthirius multifiliis |
| Trichodina reticulata | |
| Genus Huso Brandt and Ratzeburg, 1833 | |
| Species Huso huso Linnaeus, 1758 | Ichthyophthirius multifiliis |
| Order Cypriniformes | |
| Family Cyprinidae Rafinesque, 1815 | |
| Genus Arabibarbus Borkenhagen, 2014 | |
| Species Arabibarbus grypus Heckel, 1843 | |
| Synonym: Barbus grypus Heckel, 1843 | Ichthyobodo necator Ichthyophthirius multifiliis |
| Species Barbus lacerta Heckel, 1843 | |
| Synonym: Barbus lacerta cyri De Filippi, 1865 | Ichthyophthirius multifiliis |
| Trichodina domerguei | |
| Trichodina perforate | |
| Genus Capoeta Valenciennes, 1842 | |
| Species Capoeta aculeata Valenciennes, 1844 | Ichthyophthirius multifiliis |
| Species Capoeta barroisi Lortet, 1894 | Ichthyophthirius multifiliis |
| Species Capoeta capoeta Güldenstädt, 1773 | Arcella vulgaris Amphileptus branchiarum Chilodonella piscicola Ichthyobodo necator Ichthyophthirius multifiliis Stylonychia pustulata Tetrahymena pyriformis Trichodina domerguei Trichodina perforata |
| Comment: The subspecies, Capoeta capoeta gracilis Keyserling, 1861), which has been considered as an Iranian subspecies, is recognized as a full species [81] | |
| Species Capoeta damascina Valenciennes, 1842 | Ichthyophthirius multifiliis |
| Trichodina pediculus | |
| Species Capoeta razii Jouladeh-Roudbar, Eagderi, Ghanavi and Doadrio 2017 | Trichodina gracilis |
| Species Capoeta trutta Heckel, 1843 | Ichthyophthirius multifiliis |
| Genus Carasobarbus Karaman, 1971 | |
| Species Carasobarbus luteus Heckel, 1843 | Ichthyophthirius multifiliis |
| Synonym: Barbus luteus Heckel, 1843 | |
| Genus Carassius Jarocki, 1822 | Ichthyophthirius multifiliis Ichthyobodo necator Trichodina perforata |
| Species Carassius auratus Linnaeus, 1758 | |
| Synonym: Carassius auratus auratus Linnaeus, 1758 | |
| Species Carassius Carassius Linnaeus, 1758 | Ichthyophthirius multifiliis |
| Species Carassius gibelio Bloch, 1782 | Ichthyophthirius multifiliis |
| Synonym: Carassius auratus gibelio Bloch, 1782 | |
| Genus Cyprinion Heckel, 1843 | |
| Species Cyprinion macrostomum Heckel, 1843 | Ichthyophthirius multifiliis |
| Genus Cyprinus Linnaeus, 1758 | |
| Species Cyprinus carpio Linnaeus, 1758 | Goussia carpelli |
| Ichthyophthirius multifiliis | |
| Trichodina domerguei | |
| Trichodina pediculus | |
| Trichodinella subtilis | |
| Genus Luciobarbus Heckel, 1849 | |
| Species Luciobarbus barbulus Heckel, 1849 | Ichthyophthirius multifiliis |
| Synonym: Barbus barbulus Heckel, 1849 | |
| Species Luciobarbus brachycephalus Kessler, 1872 | Ichthyophthirius multifiliis |
| Synonym: Barbus brachycephalus Kessler, 1872 | |
| Species Luciobarbus capito Güldenstaedt, 1773 | Ichthyophthirius multifiliis |
| Synonym: Barbus capito Güldenstaedt, 1773 | Trichodina domerguei |
| Trichodina perforata | |
| Species Luciobarbus esocinus Heckel, 1843 | Ichthyophthirius multifiliis |
| Synonym: Barbus esocinus Heckel, 1843 | |
| Genus Mesopotamichthys Karaman, 1971 | |
| Species Mesopotamichthys sharpeyi Günther, 1874 | Ichthyophthirius multifiliis |
| Synonym: Barbus sharpeyi Günther, 1874 | |
| Genus Schizocypris Regan, 1914 | |
| Species Schizocypris altidorsalis Bianco and Banarescu, 1982 | Ichthyophthirius multifiliis |
| Comment: Schizocypris altidorsalis formerly identified as Schizocypris brucei Regan, 1914 (El-Dairi and House 2019) | |
| Genus Schizothorax Heckel, 1838 | |
| Species Schizothorax pelzami Kessler, 1870 | Ichthyophthirius multifiliis |
| Species Schizothorax zarudnyi Nikol’skii, 1897 | Ichthyophthirius multifiliis |
| Family Leuciscidae Bonaparte, 1835 | |
| Genus Abramis Cuvier, 1816 | |
| Species Abramis brama Linnaeus, 1758 | Ichthyophthirius multifiliis |
| Trichodina domerguei | |
| Trichodina perforata | |
| Genus Acanthobrama Heckel, 1843 | |
| Species Acanthobrama persidis Coad, 1981 | Ichthyophthirius multifiliis |
| Synonym: Leuciscus persidis Coad, 1981 | |
| Genus Blicca Heckel, 1843 | |
| Species Blicca bjoerkna Linnaeus, 1758 | Trichodina perforata |
| Genus Alburnoides Jeitteles, 1861 | |
| Species Alburnoides eichwaldii De Filippii, 1863 | Ichthyophthirius multifiliis Trichodina domerguei Trichodina perforata |
| Synonym: Alburnoides bipunctatus eichwaldi De Filippi, 1863 | |
| Species Alburnus chalcoides Güldenstaedt, 1772 | Ichthyophthirius multifiliis Trypanosoma percae |
| Synonym: Chalcalburnus chalcoides Güldenstädt, 1772 | |
| Species Alburnus filippii Kessler, 1877 | Trichodina perforata |
| Species Alburnus hohenackeri Kessler, 1877 | Ichthyophthirius multifiliis |
| Species Alburnus mossulensis Heckel, 1843 | |
| Synonym: Chalcalburnus mossulensis Heckel, 1843 | Ichthyophthirius multifiliis |
| Species Alburnoides tabarestanensis | Ichthyophthirius multifiliis |
| Genus Chondrostoma Agassiz, 1832 | |
| Species Chondrostoma regium Heckel, 1843 | Ichthyophthirius multifiliis |
| Species Chondrostoma orientale Bianco and Bănărescu, 1982 | Ichthyophthirius multifiliis |
| Species Leuciscus vorax Heckel, 1843 | Ichthyophthirius multifiliis |
| Synonym: Aspius vorax Heckel, 1843 | Ichthyobodo necator |
| Genus Squalius Bonaparte, 1837 | |
| Species Squalius cephalus Linnaeus, 1758 | Chilodonella cyprini |
| Synonym: Leuciscus cephalus Linnaeus, 1758 | Ichthyophthirius multifiliis |
| Genus Vimba Fitzinger, 1873 | |
| Species Vimba vimba Linnaeus 1758 | Ichthyophthirius multifiliis |
| Synonym: Vimba vimba persa Pallas, 1814 | |
| Family Xenocyprididae Günther, 1868 | |
| Genus Ctenopharyngodon Steindachner, 1866 | Balantidium ctenopharyngodoni Ichthyophthirius multifiliis |
| Species Ctenopharyngodon idella Valenciennes, 1844 | |
| Genus Hemiculter Bleeker, 1860 | |
| Species Hemiculter leucisculus Basilewsky, 1855 | Ichthyobodo necator Ichthyophthirius multifiliis Trichodina perforata |
| Genus Hypophthalmichthys Bleeker, 1859 | |
| Species Hypophthalmichthys molitrix Valenciennes, 1844 | Chilodonella piscicola Cryptobia branchialis Goussia sinensis Ichthyophthirius multifiliis |
| Species Hypophthalmichthys nobilis Valenciennes, 1844 | Ichthyophthirius multifiliis |
| Order Cyprinodontiformes | |
| Family Aphaniidae Hoedeman, 1949 | |
| Genus Aphanius Nardo, 1827 | Ichthyophthirius multifiliis |
| Species Aphanius vladykovi Coad, 1988 | Ichthyobodo necator |
| Species Aphanius sophiae Heckel, 1847 | Ichthyophthirius multifiliis |
| Order Esociformes | |
| Family Esocidae Rafinesque, 1815 | |
| Genus Esox Linnaeus, 1758 | |
| Species Esox Lucius Linnaeus, 1758 | Ichthyophthirius multifiliis |
| Order Gasterosteiformes | |
| Family Gasterosteidae Bonaparte, 1831 | |
| Genus Gasterosteus Linnaeus, 1758 | |
| Species Gasterosteus aculeatus Linnaeus, 1758 | Ichthyophthirius multifiliis |
| Order Mugiliformes | |
| Family Mugilidae Jarocki, 1822 | |
| Genus Planiliza Whitley, 1945 | |
| Species Planiliza abu Heckel, 1843 | |
| Synonym: Mugil abu Heckel, 1843; Liza abu Heckel, 1843 | Ichthyobodo necator Ichthyophthirius multifiliis |
| Genus Chelon Artedi, 1793 | |
| Species Chelon auratus Risso, 1810 | Ichthyobodo necator |
| Synonym: Mugil auratus Risso, 1810 | Ichthyophthirius multifiliis |
| Trichodina reticulata | |
| Order Perciformes | |
| Family Percidae Rafinesque, 1815 | |
| Genus Perca Linnaeus, 1758 | |
| Species Perca fluviatilis Linnaeus, 1758 | Trypanosoma percae |
| Genus Sander Oken, 1817 | |
| Species Sander lucioperca Linnaeus, 1758 | Trichodina perforata |
| Order Salmoniformes | |
| Family Salmonidae | |
| Genus Oncorhynchus Suckley, 1861 | |
| Species Oncorhynchus mykiss Walbaum, 1792 | Chilodonella cyprini |
| Synonym: Salmo gairdnerii Richardson, 1836 | Dermocystidium salmonis |
| Hexamita salmonis | |
| Ichthyophthirius multifiliis | |
| Trichodina truttae | |
| Tripartiella lata | |
| Trypanoplasma borelli | |
| Order Siluriformes | |
| Family Sisoridae Bleeker, 1858 | |
| Genus Glyptothorax Blyth, 1860 | |
| Species Glyptothorax silviae Coad, 1981 | Ichthyophthirius multifiliis |
| Family Siluridae Cuvier, 1816 | |
| Genus Silurus Linnaeus, 1758 | |
| Species Silurus glanis Linnaeus, 1758 | Ichthyophthirius multifiliis |
| Trichodina perforata | |
| Order Synbranchiformes | |
| Family Mastacembelidae Swainson, 1839 | |
| Genus Mastacembelus Scopoli, 1777 | |
| Species Mastacembelus mastacembelus Banks and Solander, 1794 | Ichthyophthirius multifiliis |
| Trichodina pediculus |
| Periods | AUCmean | AUCmean Standard Deviation | |
|---|---|---|---|
| 2041–2070 | RCP2.6 | 0.796 | 0.063 |
| RCP7.0 | 0.828 | 0.044 | |
| RCP8.5 | 0.797 | 0.040 | |
| 2071–2100 | RCP2.6 | 0.804 | 0.068 |
| RCP7.0 | 0.803 | 0.067 | |
| RCP8.5 | 0.823 | 0.037 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barzegar, M.; Raissy, M.; Shamsi, S. Protozoan Parasites of Iranian Freshwater Fishes: Review, Composition, Classification, and Modeling Distribution. Pathogens 2023, 12, 651. https://doi.org/10.3390/pathogens12050651
Barzegar M, Raissy M, Shamsi S. Protozoan Parasites of Iranian Freshwater Fishes: Review, Composition, Classification, and Modeling Distribution. Pathogens. 2023; 12(5):651. https://doi.org/10.3390/pathogens12050651
Chicago/Turabian StyleBarzegar, Maryam, Mehdi Raissy, and Shokoofeh Shamsi. 2023. "Protozoan Parasites of Iranian Freshwater Fishes: Review, Composition, Classification, and Modeling Distribution" Pathogens 12, no. 5: 651. https://doi.org/10.3390/pathogens12050651
APA StyleBarzegar, M., Raissy, M., & Shamsi, S. (2023). Protozoan Parasites of Iranian Freshwater Fishes: Review, Composition, Classification, and Modeling Distribution. Pathogens, 12(5), 651. https://doi.org/10.3390/pathogens12050651