

The Effect of Exposure to SARS-CoV-2 Vaccination and Infection on Humoral and Cellular Immunity in a Cohort of Patients with Immune-Mediated Diseases: A Pilot Study

 , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Materials and Methods
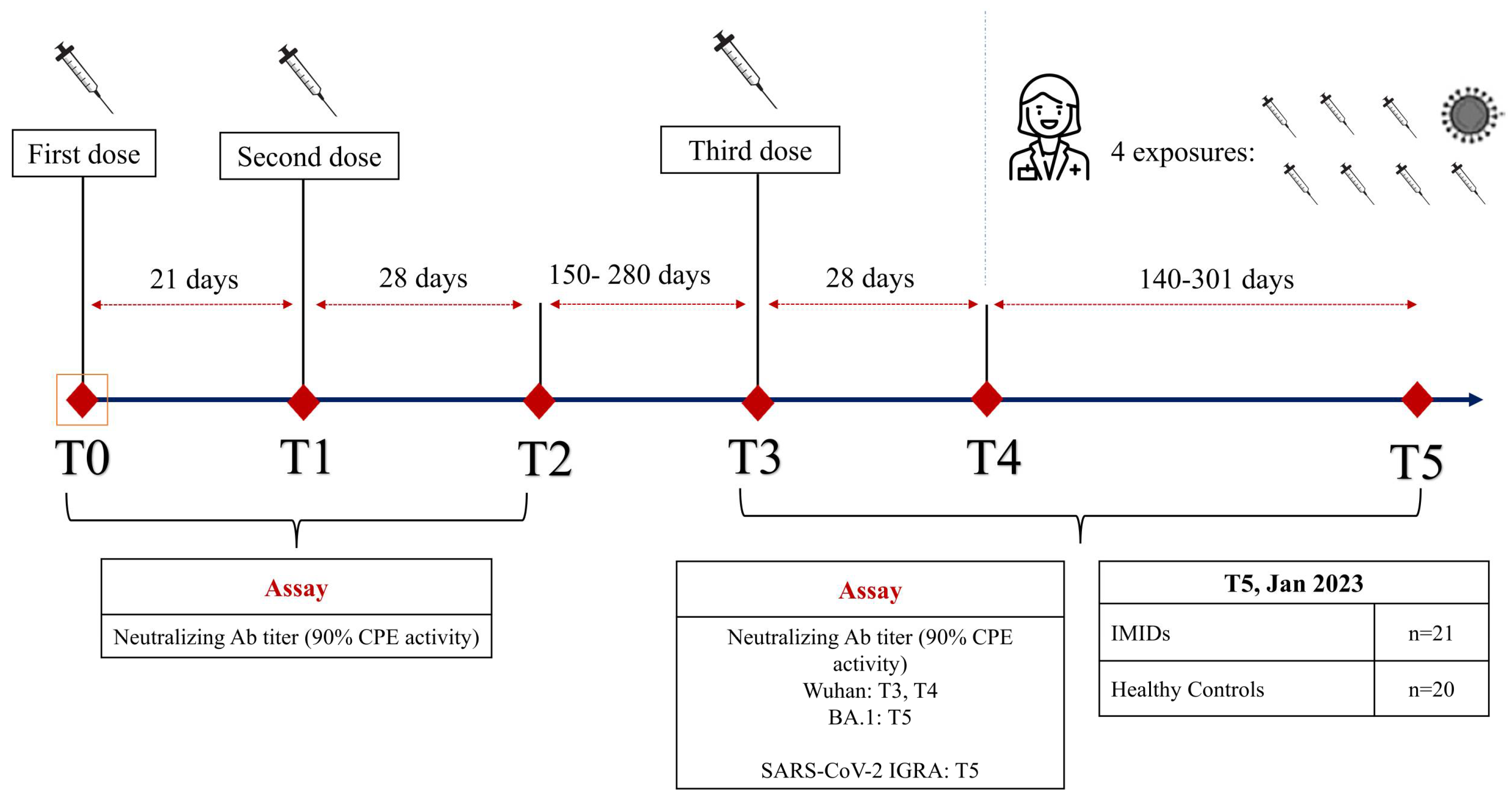
2.1. Study Design
2.2. Patient Enrolment
2.3. Sample Collection and Storing
2.4. Outcomes
2.5. SARS-CoV-2 Microneutralization Assay (MNA)
2.6. SARS-CoV-2 Specific Cellular Immunity
2.7. Statistical Analysis
2.8. Ethical Aspects
3. Results
3.1. Humoral Response
3.2. Quantitative IGRA Response
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Sahin, U.; Muik, A.; Derhovanessian, E.; Vogler, I.; Kranz, L.M.; Vormehr, M.; Baum, A.; Pascal, K.; Quandt, J.; Maurus, D.; et al. COVID-19 vaccine BNT162b1 elicits human antibody and TH1 T cell responses. Nature 2020, 586, 594–599. [Google Scholar] [CrossRef]
- Goel, R.R.; Painter, M.M.; Apostolidis, S.A.; Mathew, D.; Meng, W.; Rosenfeld, A.M.; Lundgreen, K.A.; Reynaldi, A.; Khoury, D.S.; Pattekar, A.; et al. mRNA vaccines induce durable immune memory to SARS-CoV-2 and variants of concern. Science 2021, 374, abm0829. [Google Scholar] [CrossRef] [PubMed]
- Bedekar, P.; Kearsley, A.J.; Patrone, P.N. Prevalence estimation and optimal classification methods to account for time dependence in antibody levels. J. Theor. Biol. 2023, 559, 111375. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.P.; Meyers, J.A. Immune-mediated inflammatory disorders (I.M.I.D.s): The economic and clinical costs. Am. J. Manag. Care 2002, 8, S664–S681, quiz S682–S685. [Google Scholar] [PubMed]
- Otero-Romero, S.; Ascherio, A.; Lebrun-Frénay, C. Vaccinations in multiple sclerosis patients receiving disease-modifying drugs. Curr. Opin. Neurol. 2021, 34, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Glück, T.; Kiefmann, B.; Grohmann, M.; Falk, W.; Straub, R.H.; Schölmerich, J. Immune status and risk for infection in patients receiving chronic immunosuppressive therapy. J. Rheumatol. 2005, 32, 1473–1480. [Google Scholar]
- Curtis, J.R.; Johnson, S.R.; Anthony, D.D.; Arasaratnam, R.J.; Baden, L.R.; Bass, A.R.; Calabrese, C.; Gravallese, E.M.; Harpaz, R.; Sadun, R.E.; et al. American College of Rheumatology Guidance for COVID-19 Vaccination in Patients With Rheumatic and Musculoskeletal Diseases: Version 1. Arthritis Rheumatol. 2021, 73, 1093–1107. [Google Scholar] [CrossRef] [PubMed]
- Jena, A.; Mishra, S.; Deepak, P.; Kumar-M, P.; Sharma, A.; Patel, Y.I.; Kennedy, N.A.; Kim, A.H.J.; Sharma, V.; Sebastian, S. Response to SARS-CoV-2 vaccination in immune mediated inflammatory diseases: Systematic review and meta-analysis. Autoimmun. Rev. 2022, 21, 102927. [Google Scholar] [CrossRef] [PubMed]
- Albach, F.N.; Burmester, G.R.; Biesen, R. Successful BNT162b2 booster vaccinations in a patient with rheumatoid arthritis and initially negative antibody response. Ann. Rheum. Dis. 2021, 80, 1361–1362. [Google Scholar] [CrossRef]
- Malato, J.; Ribeiro, R.M.; Fernandes, E.; Leite, P.P.; Casaca, P.; Antunes, C.; Fonseca, V.R.; Gomes, M.C.; Graca, L. Stability of hybrid versus vaccine immunity against BA.5 infection over 8 months. Lancet Infect. Dis. 2023, 23, 148–150. [Google Scholar] [CrossRef]
- Gao, Y.; Cai, C.; Grifoni, A.; Müller, T.R.; Niessl, J.; Olofsson, A.; Humbert, M.; Hansson, L.; Österborg, A.; Bergman, P.; et al. Ancestral SARS-CoV-2-specific T cells cross-recognize the Omicron variant. Nat. Med. 2022, 28, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Costanzo, G.A.M.L.; Deiana, C.M.; Sanna, G.; Perra, A.; Campagna, M.; Ledda, A.G.; Coghe, F.; Palmas, V.; Cappai, R.; Manzin, A.; et al. Impact of Exposure to Vaccination and Infection on Cellular and Antibody Response to SARS-CoV-2 in CVID Patients Through COVID-19 Pandemic. J. Clin. Immunol. 2023, 44, 12. [Google Scholar] [CrossRef] [PubMed]
- Firinu, D.; Perra, A.; Campagna, M.; Littera, R.; Fenu, G.; Meloni, F.; Cipri, S.; Sedda, F.; Conti, M.; Miglianti, M.; et al. Evaluation of antibody response to BNT162b2 mRNA COVID-19 vaccine in patients affected by immune-mediated inflammatory diseases up to 5 months after vaccination. Clin. Exp. Med. 2022, 22, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Padoan, A.; Bonfante, F.; Cosma, C.; Di Chiara, C.; Sciacovelli, L.; Pagliari, M.; Bortolami, A.; Costenaro, P.; Musso, G.; Basso, D.; et al. Analytical and clinical performances of a SARS-CoV-2 S-RBD IgG assay: Comparison with neutralization titers. Clin. Chem. Lab. Med. 2021, 59, 1444–1452. [Google Scholar] [CrossRef] [PubMed]
- Sanna, G.; Marongiu, A.; Firinu, D.; Piras, C.; Franci, G.; Galdiero, M.; Pala, G.; Palmas, V.; Angius, F.; Littera, R.; et al. Neutralizing Antibodies Responses against SARS-CoV-2 in a Sardinian Cohort Group up to 9 Months after BNT162b2 Vaccination. Vaccines 2022, 10, 531. [Google Scholar] [CrossRef] [PubMed]
- Azzolini, E.; Pozzi, C.; Germagnoli, L.; Oresta, B.; Carriglio, N.; Calleri, M.; Selmi, C.; De Santis, M.; Finazzi, S.; Carlo-Stella, C.; et al. mRNA COVID-19 vaccine booster fosters B- and T-cell responses in immunocompromised patients. Life Sci. Alliance 2022, 5, e202201381. [Google Scholar] [CrossRef] [PubMed]
- Weigang, S.; Fuchs, J.; Zimmer, G.; Schnepf, D.; Kern, L.; Beer, J.; Luxenburger, H.; Ankerhold, J.; Falcone, V.; Kemming, J.; et al. Within-host evolution of SARS-CoV-2 in an immunosuppressed COVID-19 patient as a source of immune escape variants. Nat. Commun. 2021, 12, 6405. [Google Scholar] [CrossRef] [PubMed]
- Cervia, C.; Zurbuchen, Y.; Taeschler, P.; Ballouz, T.; Menges, D.; Hasler, S.; Adamo, S.; Raeber, M.E.; Bächli, E.; Rudiger, A.; et al. Immunoglobulin signature predicts risk of post-acute COVID-19 syndrome. Nat. Commun. 2022, 13, 446. [Google Scholar] [CrossRef] [PubMed]
- Strukcinskiene, B.; Valotkiene, Z.; Jurgaitis, J.; Grigoliene, R.; Genowska, A. Immune Response to COVID-19 Vaccination in Frontline Healthcare Workers. Vaccines 2024, 12, 199. [Google Scholar] [CrossRef]
- Paniskaki, K.; Anft, M.; Meister, T.L.; Marheinecke, C.; Pfaender, S.; Skrzypczyk, S.; Seibert, F.S.; Thieme, C.J.; Konik, M.J.; Dolff, S.; et al. Immune Response in Moderate to Critical Breakthrough COVID-19 Infection After mRNA Vaccination. Front. Immunol. 2022, 13, 816220. [Google Scholar] [CrossRef]
- Merad, M.; Blish, C.A.; Sallusto, F.; Iwasaki, A. The immunology and immunopathology of COVID-19. Science 2022, 375, 1122–1127. [Google Scholar] [CrossRef] [PubMed]
- Stalman, E.W.; Wieske, L.; van Dam, K.P.J.; Kummer, L.Y.; van Kempen, Z.L.E.; Killestein, J.; Volkers, A.G.; Tas, S.W.; Boekel, L.; Wolbink, G.J.; et al. Breakthrough infections with the SARS-CoV-2 omicron (B.1.1.529) variant in patients with immune-mediated inflammatory diseases. Ann. Rheum. Dis. 2022, 81, 1757–1766. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.S.; Tan, H.-X.; Reynaldi, A.; Esterbauer, R.; Koutsakos, M.; Nguyen, J.; Amarasena, T.; Kent, H.E.; Aggarwal, A.; Turville, S.G.; et al. Durable reprogramming of neutralizing antibody responses following Omicron breakthrough infection. Sci. Adv. 2023, 9, eadg5301. [Google Scholar] [CrossRef] [PubMed]
- Barros-Martins, J.; Hammerschmidt, S.I.; Morillas Ramos, G.; Cossmann, A.; Hetzel, L.; Odak, I.; Köhler, M.; Stankov, M.V.; Ritter, C.; Friedrichsen, M.; et al. Omicron infection-associated T- and B-cell immunity in antigen-naive and triple-COVID-19-vaccinated individuals. Front. Immunol. 2023, 14, 1166589. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chandrashekar, A.; Sellers, D.; Barrett, J.; Jacob-Dolan, C.; Lifton, M.; McMahan, K.; Sciacca, M.; VanWyk, H.; Wu, C.; et al. Vaccines elicit highly conserved cellular immunity to SARS-CoV-2 Omicron. Nature 2022, 603, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Al-Dury, S.; Waldenström, J.; Ringlander, J.; Einarsdottir, S.; Andersson, M.; Hamah Saed, H.; Waern, J.; Martner, A.; Hellstrand, K.; Lagging, M. Catch-up antibody responses and hybrid immunity in mRNA vaccinated patients at risk of severe COVID-19. Infect. Dis. 2023, 55, 744–750. [Google Scholar] [CrossRef] [PubMed]
- Goldblatt, D.; Alter, G.; Crotty, S.; Plotkin, S.A. Correlates of protection against SARS-CoV-2 infection and COVID-19 disease. Immunol. Rev. 2022, 310, 6–26. [Google Scholar] [CrossRef]
- Geldof, J.; Truyens, M.; Sabino, J.; Ferrante, M.; Lambert, J.; Lapeere, H.; Hillary, T.; Van Laethem, A.; de Vlam, K.; Verschueren, P.; et al. SARS-CoV-2 infection and COVID19 vaccination across eight immune-mediated inflammatory disorders: A prospective, real-life Belgian cohort study—The BELCOMID study. Front. Immunol. 2023, 14, 1126351. [Google Scholar] [CrossRef]
- Pappa, M.; Panagiotopoulos, A.; Thomas, K.; Fanouriakis, A. Systemic Lupus Erythematosus and COVID-19. Curr. Rheumatol. Rep. 2023, 25, 192–203. [Google Scholar] [CrossRef]

{kind=link}
| Subject Characteristics | IMIDs n = 20 | HCWs n = 21 | p-Value |
|---|---|---|---|
| Age (years) median (IQR) | 51 (20.2) | 45 (23.6) | 0.02 |
| Female % | 67% | 55% | 0.44 |
| AZA or immunosuppression ongoing, % | 94% | 0% | <0.001 |
| Median dose of daily oral prednisone | 5 mg | 0 | n.a. |
| Total events (vaccine and/or infection), median (IQR) | 4 (1.6) | 4 (0.8) | 0.21 |
| Infection after 3rd dose | 10.5% | 63% | 0.0008 |
| Infection after 4th dose | 23.5% | 0% | 0.13 |
| Last event type before sampling at T5 | 25% infection 75% vaccine | 47% infection 52% vaccine | 0.21 |
| Days since the last event before sampling at T5 (IQR) | 301 | 141 | 0.043 |
| Hospital admission for COVID-19 | 1/10 | 0/16 | 0.32 |
| Duration of swab positivity, median (IQR) | 7 (12.7) | 7 (10.5) | 0.75 |
| Duration of COVID-19 symptoms, median (IQR) | 1 (6.3) | 4 (4.5) | 0.57 |
| Residual Response after Additional Exposures (SARS-CoV-2 Infection or Booster), in January 2023 (T5) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| # Of Exposures | Last Event to Sampling (Days) | nAb Titer [1:X] BA.1 | IGRA OSP IU/mL | IGRA VSP IU/mL | Rate of Residual Responders IGRA | Double Positive to nAb and IGRA | Rate of Isolated nAb Positivity | Rate of Double Negative to nAb and IGRA | |
| IMID group | 4 (1.6) | (301) | 40 (0) | 0.47 (0.33) | 0.44 (0.71) | 43% | 43% | 43% | 9.5% |
| HC group | 4 (0.8) | (141) | 40 (150) | 1.18 (2.74) | 1.14 (2.58) | 90% | 90% | 10% | 0% |
| p value | 0.21 | 0.043 | 0.43 | 0.024 | 0.03 | 0.0015 | 0.001 | 0.017 | 0.157 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costanzo, G.A.M.L.; Sanna, G.; Pes, F.; Deiana, C.M.; Ledda, A.G.; Perra, A.; Palmas, V.; Manca, V.; Miglianti, M.; Coghe, F.; et al. The Effect of Exposure to SARS-CoV-2 Vaccination and Infection on Humoral and Cellular Immunity in a Cohort of Patients with Immune-Mediated Diseases: A Pilot Study. Pathogens 2024, 13, 506. https://doi.org/10.3390/pathogens13060506
Costanzo GAML, Sanna G, Pes F, Deiana CM, Ledda AG, Perra A, Palmas V, Manca V, Miglianti M, Coghe F, et al. The Effect of Exposure to SARS-CoV-2 Vaccination and Infection on Humoral and Cellular Immunity in a Cohort of Patients with Immune-Mediated Diseases: A Pilot Study. Pathogens. 2024; 13(6):506. https://doi.org/10.3390/pathogens13060506
Chicago/Turabian StyleCostanzo, Giulia Anna Maria Luigia, Giuseppina Sanna, Francesco Pes, Carla Maria Deiana, Andrea Giovanni Ledda, Andrea Perra, Vanessa Palmas, Valeria Manca, Michela Miglianti, Ferdinando Coghe, and et al. 2024. "The Effect of Exposure to SARS-CoV-2 Vaccination and Infection on Humoral and Cellular Immunity in a Cohort of Patients with Immune-Mediated Diseases: A Pilot Study" Pathogens 13, no. 6: 506. https://doi.org/10.3390/pathogens13060506
APA StyleCostanzo, G. A. M. L., Sanna, G., Pes, F., Deiana, C. M., Ledda, A. G., Perra, A., Palmas, V., Manca, V., Miglianti, M., Coghe, F., Manzin, A., Del Giacco, S., Chessa, L., & Firinu, D. (2024). The Effect of Exposure to SARS-CoV-2 Vaccination and Infection on Humoral and Cellular Immunity in a Cohort of Patients with Immune-Mediated Diseases: A Pilot Study. Pathogens, 13(6), 506. https://doi.org/10.3390/pathogens13060506