
Quorum-Sensing Inhibition by Gram-Positive Bacteria

 ,
,  , and
, and 
Abstract
:1. Introduction: Quorum-Sensing and Quorum-Quenching as Mechanisms of Interspecies Interactions
2. Role of QS in Pathogenicity and Antimicrobial Resistance
2.1. QS of Gram-Negative Bacteria
2.1.1. QS in P. aeruginosa
2.1.2. QS in Enterohemorrhagic Escherichia coli O157:H7
2.2. QS in Gram-Positive Bacteria
2.2.1. QS in S. aureus
2.2.2. QS in Bacillus cereus
2.2.3. QS in Streptococcus pneumoniae
2.3. Regulation of Microbial Resistance with QS
2.3.1. Efflux Pumps
2.3.2. Biofilms
3. Mechanisms and Metabolites That Are Responsible for QQ in Different Groups
3.1. Firmicutes
3.1.1. Bacillus
3.1.2. Lactic Acid Bacteria as Quorum-Quenching Agents
3.2. Actinobacteria
4. The Evolutionary and Ecological Role of QQ
5. Prospects for Use: The Fight against Pathogens, the Improvement of Probiotics’ Efficiency, and Plant Protection
5.1. The Use of Microorganisms and Substances with QQ Activity to Protect Animals
5.2. Symbiotic and Mutualistic Microorganisms Capable of Protecting Plants from Pathogens, Exhibiting QQ Properties
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Turovskiy, Y.; Kashtanov, D.; Paskhover, B.; Chikindas, M.L. Quorum Sensing: Fact, Fiction, and Everything in Between. In Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 2007; Volume 62, pp. 191–234. [Google Scholar] [CrossRef] [Green Version]
- Grandclément, C.; Tannières, M.; Moréra, S.; Dessaux, Y.; Faure, D. Quorum Quenching: Role in Nature and Applied Developments. FEMS Microbiol. Rev. 2016, 40, 86–116. [Google Scholar] [CrossRef]
- Moradi, F.; Hadi, N. Quorum-Quenching Activity of Some Iranian Medicinal Plants. New Microbes New Infect. 2021, 42, 100882. [Google Scholar] [CrossRef] [PubMed]
- Weiland-Bräuer, N. Friends or Foes—Microbial Interactions in Nature. Biology 2021, 10, 496. [Google Scholar] [CrossRef] [PubMed]
- Paluch, E.; Rewak-Soroczyńska, J.; Jędrusik, I.; Mazurkiewicz, E.; Jermakow, K. Prevention of Biofilm Formation by Quorum Quenching. Appl. Microbiol. Biotechnol. 2020, 104, 1871–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haque, S.; Yadav, D.K.; Bisht, S.C.; Yadav, N.; Singh, V.; Dubey, K.K.; Jawed, A.; Wahid, M.; Dar, S.A. Quorum Sensing Pathways in Gram-Positive and-Negative Bacteria: Potential of Their Interruption in Abating Drug Resistance. J. Chemother. 2019, 31, 161–187. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus aureus Infections: Epidemiology, Pathophysiology, Clinical Manifestations, and Management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deep, A.; Chaudhary, U.; Gupta, V. Quorum Sensing and Bacterial Pathogenicity: From Molecules to Disease. J. Lab. Physicians 2011, 3, 004–011. [Google Scholar] [CrossRef]
- Saxena, P.; Joshi, Y.; Rawat, K.; Bisht, R. Biofilms: Architecture, Resistance, Quorum Sensing and Control Mechanisms. Indian J. Microbiol. 2019, 59, 3–12. [Google Scholar] [CrossRef]
- Flavier, A.B.; Clough, S.J.; Schell, M.A.; Denny, T.P. Identification of 3-hydroxypalmitic acid methyl ester as a novel autoregulator controlling virulence in Ralstonia solanacearum. Mol. Microbiol. 1997, 26, 251–259. [Google Scholar] [CrossRef]
- Barber, C.E.; Tang, J.L.; Feng, J.X.; Pan, M.Q.; Wilson, T.J.G.; Slater, H.; Dow, J.M.; Williams, P.; Daniels, M.J. A Novel Regulatory System Required for Pathogenicity of Xanthomonas campestris Is Mediated by a Small Diffusible Signal Molecule. Mol. Microbiol. 1997, 24, 555–566. [Google Scholar] [CrossRef]
- Gallagher, L.A.; McKnight, S.L.; Kuznetsova, M.S.; Pesci, E.C.; Manoil, C. Functions Required for Extracellular Quinolone Signaling by Pseudomonas aeruginosa. J. Bacteriol. Res. 2002, 184, 6472–6480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascale, G.; Sauriol, F.; Benhamou, N.; Bélanger, R.R.; Paulitz, T.C. Novel Butyrolactones with Antifungal Activity Produced by Pseudomonas aureofaciens Strain 63-28. J. Antibiot. 1997, 50, 742–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passador, L.; Cook, J.M.; Gambello, M.J.; Rust, L.; Iglewski, B.H. Expression of Pseudomonas aeruginosa Virulence Genes Requires Cell-to-Cell Communication. Science 1993, 260, 1127–1130. [Google Scholar] [CrossRef] [PubMed]
- Ledgham, F.; Ventre, I.; Soscia, C.; Foglino, M.; Sturgis, J.N.; Lazdunski, A. Interactions of the Quorum Sensing Regulator QscR: Interaction with Itself and the Other Regulators of Pseudomonas aeruginosa LasR and RhlR: Interactions of QscR. Mol. Microbiol. 2003, 48, 199–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, H.B.; DiMango, E.; Bryan, R.; Gambello, M.; Iglewski, B.H.; Goldberg, J.B.; Prince, A. Contribution of Specific Pseudomonas aeruginosa Virulence Factors to Pathogenesis of Pneumonia in a Neonatal Mouse Model of Infection. Infect. Immun. 1996, 64, 37–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walters, M.; Sperandio, V. Quorum Sensing in Escherichia coli and Salmonella. Int. J. Med. Microbiol. 2006, 296, 125–131. [Google Scholar] [CrossRef]
- Zheng, F.; Ji, H.; Cao, M.; Wang, C.; Feng, Y.; Li, M.; Pan, X.; Wang, J.; Qin, Y.; Hu, F. Contribution of the Rgg Transcription Regulator to Metabolism and Virulence of Streptococcus Suis Serotype 2. Infect. Immun. 2011, 79, 1319–1328. [Google Scholar] [CrossRef] [Green Version]
- Rutherford, S.T.; Bassler, B.L. Bacterial Quorum Sensing: Its Role in Virulence and Possibilities for Its Control. Cold Spring Harb. Perspect. Med. 2012, 2, 1. [Google Scholar] [CrossRef]
- Kirchdoerfer, R.N.; Garner, A.L.; Flack, C.E.; Mee, J.M.; Horswill, A.R.; Janda, K.D.; Kaufmann, G.F.; Wilson, I.A. Structural Basis for Ligand Recognition and Discrimination of a Quorum-Quenching Antibody. J. Biol. Chem. 2011, 286, 17351–17358. [Google Scholar] [CrossRef] [Green Version]
- Kutty, S.K.; Barraud, N.; Pham, A.; Iskander, G.; Rice, S.A.; Black, D.S.; Kumar, N. Design, Synthesis, and Evaluation of Fimbrolide–Nitric Oxide Donor Hybrids as Antimicrobial Agents. J. Med. Chem. 2013, 56, 9517–9529. [Google Scholar] [CrossRef]
- Donelli, G. (Ed.) Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2016; Volume 901. [Google Scholar] [CrossRef]
- Slamti, L.; Lereclus, D. Specificity and Polymorphism of the PlcR-PapR Quorum-Sensing System in the Bacillus Cereus Group. J. Bacteriol. 2005, 187, 1182–1187. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, G.; Ray, A.K. Quorum-Sensing Network-Associated Gene Regulation in Gram-Positive Bacteria. Acta Microbiol. Immunol. Hung. 2017, 64, 439–453. [Google Scholar] [CrossRef] [Green Version]
- Suntharalingam, P.; Cvitkovitch, D.G. Quorum Sensing in Streptococcal Biofilm Formation. Trends Microbiol. 2005, 13, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Waters, C.M.; Bassler, B.L. Quorum Sensing: Cell-to-Cell Communication in Bacteria. Annu. Rev. Cell Dev. Biol. 2005, 21, 319–346. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Zhou, X.; Li, Y. Strategies for Streptococcus mutans biofilm dispersal through extracellular polymeric substances disruption. Mol. Oral Microbiol. 2021, 37, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yu, Z.; Ding, T. Quorum-Sensing Regulation of Antimicrobial Resistance in Bacteria. Microorganisms 2020, 8, 425. [Google Scholar] [CrossRef] [Green Version]
- Pumbwe, L.; Skilbeck, C.A.; Wexler, H.M. Presence of Quorum-Sensing Systems Associated with Multidrug Resistance and Biofilm Formation in Bacteroides fragilis. Microb. Ecol. 2008, 56, 412–419. [Google Scholar] [CrossRef]
- Maseda, H.; Sawada, I.; Saito, K.; Uchiyama, H.; Nakae, T.; Nomura, N. Enhancement of the MexAB-OprM Efflux Pump Expression by a Quorum-Sensing Autoinducer and Its Cancellation by a Regulator, MexT, of the MexEF-OprN Efflux Pump Operon in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2004, 48, 1320–1328. [Google Scholar] [CrossRef] [Green Version]
- Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance. Microbiol. Spectr. 2016, 4, 2–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cirou, A.; Diallo, S.; Kurt, C.; Latour, X.; Faure, D. Growth Promotion of Quorum-Quenching Bacteria in the Rhizosphere of Solanum tuberosum. Environ. Microbiol. 2007, 9, 1511–1522. [Google Scholar] [CrossRef]
- Jafra, S.; Przysowa, J.; Czajkowski, R.; Michta, A.; Garbeva, P.; Van Der Wolf, J.M. Detection and Characterization of Bacteria from the Potato Rhizosphere Degrading N-Acyl-Homoserine Lactone. Can. J. Microbiol. 2006, 52, 1006–1015. [Google Scholar] [CrossRef]
- Yavuztürk Gül, B.; Koyuncu, I. Assessment of New Environmental Quorum Quenching Bacteria as a Solution for Membrane Biofouling. Process. Biochem. 2017, 61, 137–146. [Google Scholar] [CrossRef]
- Dong, Y.-H.; Gusti, A.R.; Zhang, Q.; Xu, J.-L.; Zhang, L.-H. Identification of Quorum-Quenching N-Acyl Homoserine Lactonases from Bacillus Species. Appl. Environ. Microbiol. 2002, 68, 1754–1759. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Dubey, R.C.; Maheshwari, D.K. Bacillus Strains Isolated from Rhizosphere Showed Plant Growth Promoting and Antagonistic Activity against Phytopathogens. Microbiol. Res. 2012, 167, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Moore, T. Antagonistic Activity of Bacillus Bacteria against Food-Borne Pathogens. J. Probiotics Health 2013, 1, 110. [Google Scholar] [CrossRef] [Green Version]
- Olishevska, S.; Nickzad, A.; Déziel, E. Bacillus and Paenibacillus Secreted Polyketides and Peptides Involved in Controlling Human and Plant Pathogens. Appl. Microbiol. Biotechnol. 2019, 103, 1189–1215. [Google Scholar] [CrossRef] [PubMed]
- Anandan, K.; Vittal, R.R. Quorum Quenching Activity of AiiA Lactonase KMMI17 from Endophytic Bacillus Thuringiensis KMCL07 on AHL-Mediated Pathogenic Phenotype in Pseudomonas aeruginosa. Microb. Pathog. 2019, 132, 230–242. [Google Scholar] [CrossRef]
- Hartmann, A.; Rothballer, M. Role of Quorum Sensing Signals of Rhizobacteria for Plant Growth Promotion. In Rhizotrophs: Plant Growth Promotion to Bioremediation; Mehnaz, S., Ed.; Springer: Singapore, 2017. [Google Scholar] [CrossRef]
- Hartmann, A.; Rothballer, M.; Hense, B.A.; Schröder, P. Bacterial Quorum Sensing Compounds Are Important Modulators of Microbe-Plant Interactions. Front. Plant Sci. 2014, 5, 131. [Google Scholar] [CrossRef] [Green Version]
- Schikora, A.; Schenk, S.T.; Hartmann, A. Beneficial Effects of Bacteria-Plant Communication Based on Quorum Sensing Molecules of TheN-Acyl Homoserine Lactone Group. Plant Mol. Biol. 2016, 90, 605–612. [Google Scholar] [CrossRef]
- Schneider, J.; Yepes, A.; Garcia-Betancur, J.C.; Westedt, I.; Mielich, B.; López, D. Streptomycin-Induced Expression in Bacillus subtilis of YtnP, a Lactonase-Homologous Protein That Inhibits Development and Streptomycin Production in Streptomyces griseus. Appl. Environ. Microbiol. 2012, 78, 599–603. [Google Scholar] [CrossRef] [Green Version]
- McBride, S.G.; Strickland, M.S. Quorum Sensing Modulates Microbial Efficiency by Regulating Bacterial Investment in Nutrient Acquisition Enzymes. Soil Biol. Biochem. 2019, 136, 107514. [Google Scholar] [CrossRef]
- Nithya, C.; Aravindraja, C.; Pandian, S.K. Bacillus pumilus of Palk Bay Origin Inhibits Quorum-Sensing-Mediated Virulence Factors in Gram-Negative Bacteria. Res. Microbiol. 2010, 161, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.; Lu, F.; Zhu, W.; Kang, C. Isolation and Characterization of New Potential Probiotic Bacteria Based on Quorum-Sensing System: AHLs-Degrading Bacteria as Probiotic Candidate Isolated from Fish Gut. J. Appl. Microbiol. 2011, 110, 202–208. [Google Scholar] [CrossRef]
- Piewngam, P.; Zheng, Y.; Nguyen, T.H.; Dickey, S.W.; Joo, H.-S.; Villaruz, A.E.; Glose, K.A.; Fisher, E.L.; Hunt, R.L.; Li, B.; et al. Pathogen Elimination by Probiotic Bacillus via Signalling Interference. Nature 2018, 562, 532–537. [Google Scholar] [CrossRef]
- Gunaratnam, S.; Millette, M.; McFarland, L.V.; DuPont, H.L.; Lacroix, M. Potential Role of Probiotics in Reducing Clostridioides difficile Virulence: Interference with Quorum Sensing Systems. Microb. Pathog. 2021, 153, 104798. [Google Scholar] [CrossRef]
- Algburi, A.; Zehm, S.; Netrebov, V.; Bren, A.B.; Chistyakov, V.; Chikindas, M.L. Subtilosin Prevents Biofilm Formation by Inhibiting Bacterial Quorum Sensing. Probiotics Antimicrob. Proteins 2017, 9, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Colegio, O.R.; Chu, N.-Q.; Szabo, A.L.; Chu, T.; Rhebergen, A.M.; Jairam, V.; Cyrus, N.; Brokowski, C.E.; Eisenbarth, S.C.; Phillips, G.M.; et al. Functional Polarization of Tumour-Associated Macrophages by Tumour-Derived Lactic Acid. Nature 2014, 513, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, J.; Kim, Y.; Oh, S.; Song, M.; Choe, J.H.; Whang, K.-Y.; Kim, K.H.; Oh, S. Influences of Quorum-Quenching Probiotic Bacteria on the Gut Microbial Community and Immune Function in Weaning Pigs. Anim. Sci. J. 2018, 89, 412–422. [Google Scholar] [CrossRef]
- Kim, A.-L.; Park, S.-Y.; Lee, C.-H.; Lee, C.-H.; Lee, J.-K. Quorum Quenching Bacteria Isolated from the Sludge of a Wastewater Treatment Plant and Their Application for Controlling Biofilm Formation. J. Microbiol. Biotechnol. 2014, 24, 1574–1582. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Lee, J.W.; Kang, S.-G.; Oh, S.; Griffiths, M.W. Bifidobacterium Spp. Influences the Production of Autoinducer-2 and Biofilm Formation by Escherichia coli O157:H7. Anaerobe 2012, 18, 539–545. [Google Scholar] [CrossRef]
- Kampouris, I.D.; Karayannakidis, P.D.; Banti, D.C.; Sakoula, D.; Konstantinidis, D.; Yiangou, M.; Samaras, P.E. Evaluation of a Novel Quorum Quenching Strain for MBR Biofouling Mitigation. Water Res. 2018, 143, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, W.; Xu, S.X.; Magarvey, N.A.; McCormick, J.K. Lactobacillus reuteri-Produced Cyclic Dipeptides Quench Agr-Mediated Expression of Toxic Shock Syndrome Toxin-1 in Staphylococci. Proc. Natl. Acad Sci. USA 2011, 108, 3360–3365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Cui, F.; Bai, F.; Zhao, G.; Li, J. Involvement of Acylated Homoserine Lactones (AHLs) of Aeromonas sobria in Spoilage of Refrigerated Turbot (Scophthalmus maximus L.). Sensors 2016, 16, 1083. [Google Scholar] [CrossRef] [PubMed]
- Morohoshi, T.; Fukamachi, K.; Kato, M.; Kato, N.; Ikeda, T. Regulation of the Violacein Biosynthetic Gene Cluster by Acylhomoserine Lactone-Mediated Quorum Sensing in Chromobacterium violaceum ATCC 12472. Biosci. Biotechnol. Biochem. 2010, 74, 2116–2119. [Google Scholar] [CrossRef]
- Cui, T.; Bai, F.; Sun, M.; Lv, X.; Li, X.; Zhang, D.; Du, H. Lactobacillus crustorum ZHG 2–1 as Novel Quorum-Quenching Bacteria Reducing Virulence Factors and Biofilms Formation of Pseudomonas aeruginosa. LWT 2020, 117, 108696. [Google Scholar] [CrossRef]
- Rana, S.; Bhawal, S.; Kumari, A.; Kapila, S.; Kapila, R. PH-Dependent Inhibition of AHL-Mediated Quorum Sensing by Cell-Free Supernatant of Lactic Acid Bacteria in Pseudomonas aeruginosa PAO1. Microb. Pathog. 2020, 142, 104105. [Google Scholar] [CrossRef]
- Dong, W.; Zhu, J.; Guo, X.; Kong, D.; Zhang, Q.; Zhou, Y.; Liu, X.; Zhao, S.; Ruan, Z. Characterization of AiiK, an AHL Lactonase, from Kurthia huakui LAM0618T and Its Application in Quorum Quenching on Pseudomonas aeruginosa PAO1. Sci. Rep. 2018, 8, 6013. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.-H.; Weng, L.-X.; Dong, Y.-H.; Zhang, L.-H. Specificity and Enzyme Kinetics of the Quorum-Quenching N-Acyl Homoserine Lactone Lactonase (AHL-Lactonase). J. Biol. Chem. 2004, 279, 13645–13651. [Google Scholar] [CrossRef] [Green Version]
- Yates, E.A.; Philipp, B.; Buckley, C.; Atkinson, S.; Chhabra, S.R.; Sockett, R.E.; Goldner, M.; Dessaux, Y.; Cámara, M.; Smith, H.; et al. N-Acylhomoserine Lactones Undergo Lactonolysis in a PH-, Temperature-, and Acyl Chain Length-Dependent Manner during Growth of Yersinia pseudotuberculosis and Pseudomonas aeruginosa. Infect. Immun. 2002, 70, 5635–5646. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-Z.; Plésiat, P.; Nikaido, H. The Challenge of Efflux-Mediated Antibiotic Resistance in Gram-Negative Bacteria. Clin. Microbiol. Rev. 2015, 28, 337–418. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Gu, S.; Cui, X.; Shi, Y.; Wen, S.; Chen, H.; Ge, J. Antimicrobial, Anti-Adhesive and Anti-Biofilm Potential of Biosurfactants Isolated from Pediococcus acidilactici and Lactobacillus plantarum against Staphylococcus Aureus CMCC26003. Microb. Pathog. 2019, 127, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.I.; Mizan, M.F.R.; Roy, P.K.; Nahar, S.; Toushik, S.H.; Ashrafudoulla, M.; Jahid, I.K.; Lee, J.; Ha, S.-D. Listeria monocytogenes Biofilm Inhibition on Food Contact Surfaces by Application of Postbiotics from Lactobacillus curvatus B.67 and Lactobacillus plantarum M.2. Food Res. Int. 2021, 148, 110595. [Google Scholar] [CrossRef]
- Yong, C.C.; Lim, J.; Kim, B.-K.; Park, D.-J.; Oh, S. Suppressive Effect of Lactobacillus fermentum Lim2 on Clostridioides Difficile 027 Toxin Production. Lett. Appl. Microbiol. 2019, 68, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Cui, T.; Du, H.; Sun, M.; Bai, F.; Li, J.; Zhang, D. Lactobacillus plantarum CY 1–1: A Novel Quorum Quenching Bacteria and Anti-Biofilm Agent against Aeromonas sobria. LWT 2021, 137, 110439. [Google Scholar] [CrossRef]
- Selvakumar, G.; Panneerselvam, P.; Ganeshamurthy, A.N. Diversity Utility and Potential of Actinobacteria in the Agro-Ecosystem. In Bacterial Diversity in Sustainable Agriculture; Sustainable Development and Biodiversity; Maheshwari, D.K., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2014; pp. 23–40. [Google Scholar] [CrossRef]
- Recio, E.; Colinas, Á.; Rumbero, Á.; Aparicio, J.F.; Martín, J.F. PI Factor, a Novel Type Quorum-Sensing Inducer Elicits Pimaricin Production in Streptomyces natalensis. J. Biol. Chem. 2004, 279, 41586–41593. [Google Scholar] [CrossRef] [Green Version]
- Vesuna, A.P.; Nerurkar, A.S. Biocontrol Impact of AHL Degrading Actinobacteria on Quorum Sensing Regulated Virulence of Phytopathogen Pectobacterium carotovorum subsp. carotovorum BR1. Plant Soil 2020, 453, 371–388. [Google Scholar] [CrossRef]
- Weber, T.; Welzel, K.; Pelzer, S.; Vente, A.; Wohlleben, W. Exploiting the Genetic Potential of Polyketide Producing Streptomycetes. J. Biotechnol. 2003, 106, 221–232. [Google Scholar] [CrossRef]
- Wijaya, M.; Delicia, D.; Waturangi, D.E. Screening and Quantification of Antiquorum-Sensing Activity of Actinobacteria Isolates Against Gram-Positive and Gram-Negative Biofilm Associated Bacteria. Res. Sq. 2021, 1–2, 1. [Google Scholar] [CrossRef]
- Mulya, E.; Waturangi, D.E. Screening and Quantification of Anti-Quorum Sensing and Antibiofilm Activity of Actinomycetes Isolates against Food Spoilage Biofilm-Forming Bacteria. BMC Microbiol. 2021, 21, 52. [Google Scholar] [CrossRef] [PubMed]
- Chankhamhaengdecha, S.; Hongvijit, S.; Srichaisupakit, A.; Charnchai, P.; Panbangred, W. Endophytic Actinomycetes: A Novel Source of Potential Acyl Homoserine Lactone Degrading Enzymes. BioMed Res. Int. 2013, 2013, e782847. [Google Scholar] [CrossRef]
- Park, S.-Y.; Kang, H.-O.; Jang, H.-S.; Lee, J.-K.; Koo, B.-T.; Yum, D.-Y. Identification of Extracellular N-Acylhomoserine Lactone Acylase from a Streptomyces sp. and Its Application to Quorum Quenching. Appl. Environ. Microbiol. 2005, 71, 2632–2641. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.-Z.; Morohoshi, T.; Ikenoya, M.; Someya, N.; Ikeda, T. AiiM, a Novel Class of N-Acylhomoserine Lactonase from the Leaf-Associated Bacterium Microbacterium testaceum. Appl. Environ. Microbiol. 2010, 76, 2524–2530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devaraj, K.; Tan, G.Y.A.; Chan, K.-G. Quorum Quenching Properties of Actinobacteria Isolated from Malaysian Tropical Soils. Arch. Microbiol. 2017, 199, 897–906. [Google Scholar] [CrossRef]
- Junji, I.; Hideo, T.; Haruo, I.; Satoshi, O. Cloning and Sequencing of the Aculeacin a Acylase-Encoding Gene from Actinoplanes Utahensis and Expression in Streptomyces lividans. Gene 1992, 119, 29–35. [Google Scholar] [CrossRef]
- Park, S.-Y.; Lee, S.J.; Oh, T.-K.; Oh, J.-W.; Koo, B.-T.; Yum, D.-Y.; Lee, J.-K. AhlD, an N-Acylhomoserine Lactonase in Arthrobacter sp., and Predicted Homologues in Other Bacteria. Microbiology 2003, 149, 1541–1550. [Google Scholar] [CrossRef] [Green Version]
- Naik, D.N.; Wahidullah, S.; Meena, R.M. Attenuation of Pseudomonas aeruginosa Virulence by Marine Invertebrate–Derived Streptomyces sp. Lett. Appl. Microbiol. 2013, 56, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Koul, S.; Kalia, V.C. Multiplicity of Quorum Quenching Enzymes: A Potential Mechanism to Limit Quorum Sensing Bacterial Population. Indian J. Microbiol. 2017, 57, 100–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cirou, A.; Mondy, S.; An, S.; Charrier, A.; Sarrazin, A.; Thoison, O.; DuBow, M.; Faure, D. Efficient Biostimulation of Native and Introduced Quorum-Quenching Rhodococcus erythropolis Populations Is Revealed by a Combination of Analytical Chemistry, Microbiology, and Pyrosequencing. Appl. Environ. Microbiol. 2012, 78, 481–492. [Google Scholar] [CrossRef] [Green Version]
- Uroz, S.; Oger, P.M.; Chapelle, E.; Adeline, M.-T.; Faure, D.; Dessaux, Y. A Rhodococcus QsdA-Encoded Enzyme Defines a Novel Class of Large-Spectrum Quorum-Quenching Lactonases. Appl. Environ. Microbiol. 2008, 74, 1357–1366. [Google Scholar] [CrossRef] [Green Version]
- Uroz, S.; Chhabra, S.R.; Cámara, M.; Williams, P.; Oger, P.; Dessaux, Y. N-Acylhomoserine Lactone Quorum-Sensing Molecules Are Modified and Degraded by Rhodococcus erythropolis W2 by Both Amidolytic and Novel Oxidoreductase Activities. Microbiology 2005, 151, 3313–3322. [Google Scholar] [CrossRef] [Green Version]
- Barbey, C.; Chane, A.; Burini, J.-F.; Maillot, O.; Merieau, A.; Gallique, M.; Beury-Cirou, A.; Konto-Ghiorghi, Y.; Feuilloley, M.; Gobert, V.; et al. A Rhodococcal Transcriptional Regulatory Mechanism Detects the Common Lactone Ring of AHL Quorum-Sensing Signals and Triggers the Quorum-Quenching Response. Front. Microbiol. 2018, 9, 2800. [Google Scholar] [CrossRef] [PubMed]
- Harper, D.R.; Parracho, H.M.R.T.; Walker, J.; Sharp, R.; Hughes, G.; Werthén, M.; Lehman, S.; Morales, S. Bacteriophages and Biofilms. Antibiotics 2014, 3, 270–284. [Google Scholar] [CrossRef]
- Drucker, V.V.; Gorshkova, A.S. Bacteriophages and their functioning in the biofilms. Bull. Irkutsk. State Univ. 2012, 5, 8–16. [Google Scholar]
- Zhou, L.; Zhang, Y.; Ge, Y.; Zhu, X.; Pan, J. Regulatory Mechanisms and Promising Applications of Quorum Sensing-Inhibiting Agents in Control of Bacterial Biofilm Formation. Front. Microbiol. 2020, 11, 589640. [Google Scholar] [CrossRef] [PubMed]
- Villa, T.G.; Feijoo-Siota, L.; Rama, J.R.; Sánchez-Pérez, A.; Viñas, M. Horizontal Gene Transfer Between Bacteriophages and Bacteria: Antibiotic Resistances and Toxin Production. In Horizontal Gene Transfer; Villa, T.G., Viñas, M., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 97–142. [Google Scholar] [CrossRef]
- Chu, Y.-Y.; Nega, M.; Wölfle, M.; Plener, L.; Grond, S.; Jung, K.; Götz, F. A New Class of Quorum Quenching Molecules from Staphylococcus Species Affects Communication and Growth of Gram-Negative Bacteria. PLoS Pathog. 2013, 9, e1003654. [Google Scholar] [CrossRef] [PubMed]
- Kalia, V.C.; Raju, S.C.; Purohit, H.J. Genomic analysis reveals versatile organisms for quorum quenching enzymes: Acyl-homoserine lactone-acylase and -lactonase. Open Microbiol. J. 2011, 5, 1–13. [Google Scholar] [CrossRef]
- Goh, S.Y.; Khan, S.A.; Tee, K.K.; Kasim, N.H.A.; Yin, W.F.; Chan, K.G. Quorum sensing activity of Citrobacter amalonaticus L8A, a bacterium isolated from dental plaque. Sci. Rep. 2016, 6, 20702. [Google Scholar] [CrossRef]
- Li, Y.-S.; Tian, T.; Li, B.-B.; Yu, H.-Q. Longer Persistence of Quorum Quenching Bacteria over Quorum Sensing Bacteria in Aerobic Granules. Water Res. 2020, 179, 115904. [Google Scholar] [CrossRef]
- Dong, Y.-H.; Wang, L.-H.; Xu, J.-L.; Zhang, H.-B.; Zhang, X.-F.; Zhang, L.-H. Quenching Quorum-Sensing-Dependent Bacterial Infection by an N-Acyl Homoserine Lactonase. Nature 2001, 411, 813–817. [Google Scholar] [CrossRef]
- Cao, Y.; He, S.; Zhou, Z.; Zhang, M.; Mao, W.; Zhang, H.; Yao, B. Orally Administered Thermostable N-Acyl Homoserine Lactonase from Bacillus Sp. Strain AI96 Attenuates Aeromonas hydrophila Infection in Zebrafish. Appl. Environ. Microbiol. 2012, 78, 1899–1908. [Google Scholar] [CrossRef] [Green Version]
- Fong, J.; Zhang, C.; Yang, R.; Boo, Z.Z.; Tan, S.K.; Nielsen, T.E.; Givskov, M.; Liu, X.-W.; Bin, W.; Su, H.; et al. Combination Therapy Strategy of Quorum Quenching Enzyme and Quorum Sensing Inhibitor in Suppressing Multiple Quorum Sensing Pathways of P. aeruginosa. Sci. Rep. 2018, 8, 1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalia, V.C.; Kumar, P.; Pandian, S.K.T.; Sharma, P. Biofouling Co 15. Biofouling Control by Quorum Quenching; Springer: Berlin/Heidelberg, Germany, 2015; pp. 431–440. [Google Scholar] [CrossRef]
- Algburi, A.; Comito, N.; Kashtanov, D.; Dicks, L.M.T.; Chikindas, M.L. Control of Biofilm Formation: Antibiotics and Beyond. Appl. Environ. Microbiol. 2017, 83, e02508-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, A.; Nehra, K.; Rana, J.S.; Dahiya, T. Antibiotic resistance in agriculture: Perspectives on upcoming strategies to overcome upsurge in resistance. Curr. Res. Microb. Sci. 2021, 2, 100030. [Google Scholar] [CrossRef] [PubMed]
- Gorovtsov, A.V.; Sazykin, I.S.; Sazykina, M.A. The influence of heavy metals, polyaromatic hydrocarbons, and polychlorinated biphenyls pollution on the development of antibiotic resistance in soils. Environ. Sci. Pollut. Res. 2018, 25, 9283–9292. [Google Scholar] [CrossRef]
- Garge, S.S.; Nerurkar, A.S. Evaluation of Quorum Quenching Bacillus Spp. for Their Biocontrol Traits against Pectobacterium carotovorum Subsp. Carotovorum Causing Soft Rot. Biocatal. Agric. Biotechnol. 2017, 9, 48–57. [Google Scholar] [CrossRef]
- Beckmann, B.E.; Knoester, D.B.; Connelly, B.D.; Waters, C.M.; McKinley, P.K. Evolution of Resistance to Quorum Quenching in Digital Organisms. Artif. Life 2012, 18, 291–310. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S. The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Demin, K.A.; Refeld, A.G.; Bogdanova, A.A.; Prazdnova, E.V.; Popov, I.V.; Kutsevalova, O.Y.; Ermakov, A.M.; Bren, A.B.; Rudoy, D.V.; Chistyakov, V.A.; et al. Mechanisms of Candida Resistance to Antimycotics and Promising Ways to Overcome It: The Role of Probiotics. Probiotics Antimicrob. Proteins 2021, 13, 926–948. [Google Scholar] [CrossRef]
- Dong, Y.-H.; Wang, L.-H.; Zhang, L.-H. Quorum-Quenching Microbial Infections: Mechanisms and Implications. Philos. Trans. R. Soc. B 2007, 362, 1201–1211. [Google Scholar] [CrossRef]
- Zhou, S.; Yu, Z.; Chu, W. Effect of Quorum-Quenching Bacterium Bacillus sp. QSI-1 on Protein Profiles and Extracellular Enzymatic Activities of Aeromonas hydrophila YJ-1. BMC Microbiol. 2019, 19, 135. [Google Scholar] [CrossRef]
- Chu, W.; Zhou, S.; Zhu, W.; Zhuang, X. Quorum Quenching Bacteria Bacillus sp. QSI-1 Protect Zebrafish (Danio Rerio) from Aeromonas hydrophila Infection. Sci. Rep. 2014, 4, 5446. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Peng, M.; Tong, W.; Zhang, Q.; Song, Z. The Quorum Quenching Bacterium Bacillus licheniformis T-1 Protects Zebrafish against Aeromonas hydrophila Infection. Probiotics Antimicrob. Proteins 2020, 12, 160–171. [Google Scholar] [CrossRef]
- Ghanei-Motlagh, R.; Mohammadian, T.; Gharibi, D.; Menanteau-Ledouble, S.; Mahmoudi, E.; Khosravi, M.; Zarea, M.; El-Matbouli, M. Quorum Quenching Properties and Probiotic Potentials of Intestinal Associated Bacteria in Asian Sea Bass Lates calcarifer. Mar. Drugs 2019, 18, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghanei-Motlagh, R.; Gharibi, D.; Mohammadian, T.; Khosravi, M.; Mahmoudi, E.; Zarea, M.; Menanteau-Ledouble, S.; El-Matbouli, M. Feed Supplementation with Quorum Quenching Probiotics with Anti-Virulence Potential Improved Innate Immune Responses, Antioxidant Capacity and Disease Resistance in Asian Seabass (Lates calcarifer). Aquaculture 2021, 535, 736345. [Google Scholar] [CrossRef]
- Kiarood, S.L.A.; Rahnama, K.; Golmohammadi, M.; Nasrollanejad, S. Quorum-Quenching Endophytic Bacteria Inhibit Disease Caused by Pseudomonas syringae Pv. Syringae in Citrus Cultivars. J. Basic Microbiol. 2020, 1, 12. [Google Scholar] [CrossRef]
- Valdéz, J.C.; Peral, M.C.; Rachid, M.; Santana, M.; Perdigón, G. Interference of Lactobacillus Plantarum with Pseudomonas aeruginosa in Vitro and in Infected Burns: The Potential Use of Probiotics in Wound Treatment. Clin. Microbiol. Infect. 2005, 11, 472–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vadassery, D.H.; Pillai, D. Quorum Quenching Potential of Enterococcus faecium QQ12 Isolated from Gastrointestinal Tract of Oreochromis Niloticus and Its Application as a Probiotic for the Control of Aeromonas hydrophila Infection in Goldfish Carassius auratus (Linnaeus 1758). Braz. J. Microbiol. 2020, 51, 1333–1343. [Google Scholar] [CrossRef] [PubMed]
- Wasfi, R.; Abd El-Rahman, O.A.; Zafer, M.M.; Ashour, H.M. Probiotic Lactobacillus sp. Inhibit Growth, Biofilm Formation and Gene Expression of Caries-Inducing Streptococcus mutans. J. Cell. Mol. Med. 2018, 22, 1972–1983. [Google Scholar] [CrossRef] [Green Version]
- Méndez-Vilas, A. Industrial, Medical and Environmental Applications of Microorganisms: Current Status and Trends; Wageningen Academic Publishers: Wageningen, The Netherlands, 2014; Volume 698. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.-H.; Xu, J.-L.; Li, X.-Z.; Zhang, L.-H. AiiA, an Enzyme That Inactivates the Acylhomoserine Lactone Quorum-Sensing Signal and Attenuates the Virulence of Erwinia carotovora. Proc. Natl. Acad. Sci. USA 2000, 97, 3526–3531. [Google Scholar] [CrossRef]
- Carlier, A.; Uroz, S.; Smadja, B.; Fray, R.; Latour, X.; Dessaux, Y.; Faure, D. The Ti Plasmid of Agrobacterium tumefaciens Harbors an AttM-Paralogous Gene, AiiB, Also Encoding N-Acyl Homoserine Lactonase Activity. Appl. Environ. Microbiol. 2003, 69, 4989–4993. [Google Scholar] [CrossRef] [Green Version]
- Molina, L.; Rezzonico, F.; Défago, G.; Duffy, B. Autoinduction in Erwinia amylovora: Evidence of an Acyl-Homoserine Lactone Signal in the Fire Blight Pathogen. J. Bacteriol. 2005, 187, 3206–3213. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.-H.; Zhang, X.-F.; Xu, J.-L.; Zhang, L.-H. Insecticidal Bacillus Thuringiensis Silences Erwinia carotovora Virulence by a New Form of Microbial Antagonism, Signal Interference. Appl. Environ. Microbiol. 2004, 70, 954–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alinejad, F.; Shahryari, F.; Eini, O.; Sarafraz-Niko, F.; Shekari, A.; Setareh, M. Screening of Quorum-Quenching Bacteria Associated with Rhizosphere as Biocontrol Agents of Pectobacterium carotovorum subsp. Carotovorum. Arch. Phytopathol 2020, 53, 509–523. [Google Scholar] [CrossRef]
- Zamani, M.; Behboudi, K.; Ahmadzadeh, M. Quorum Quenching by Bacillus cereus U92: A Double-Edged Sword in Biological Control of Plant Diseases. Biocontrol Sci. Technol. 2013, 23, 555–573. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
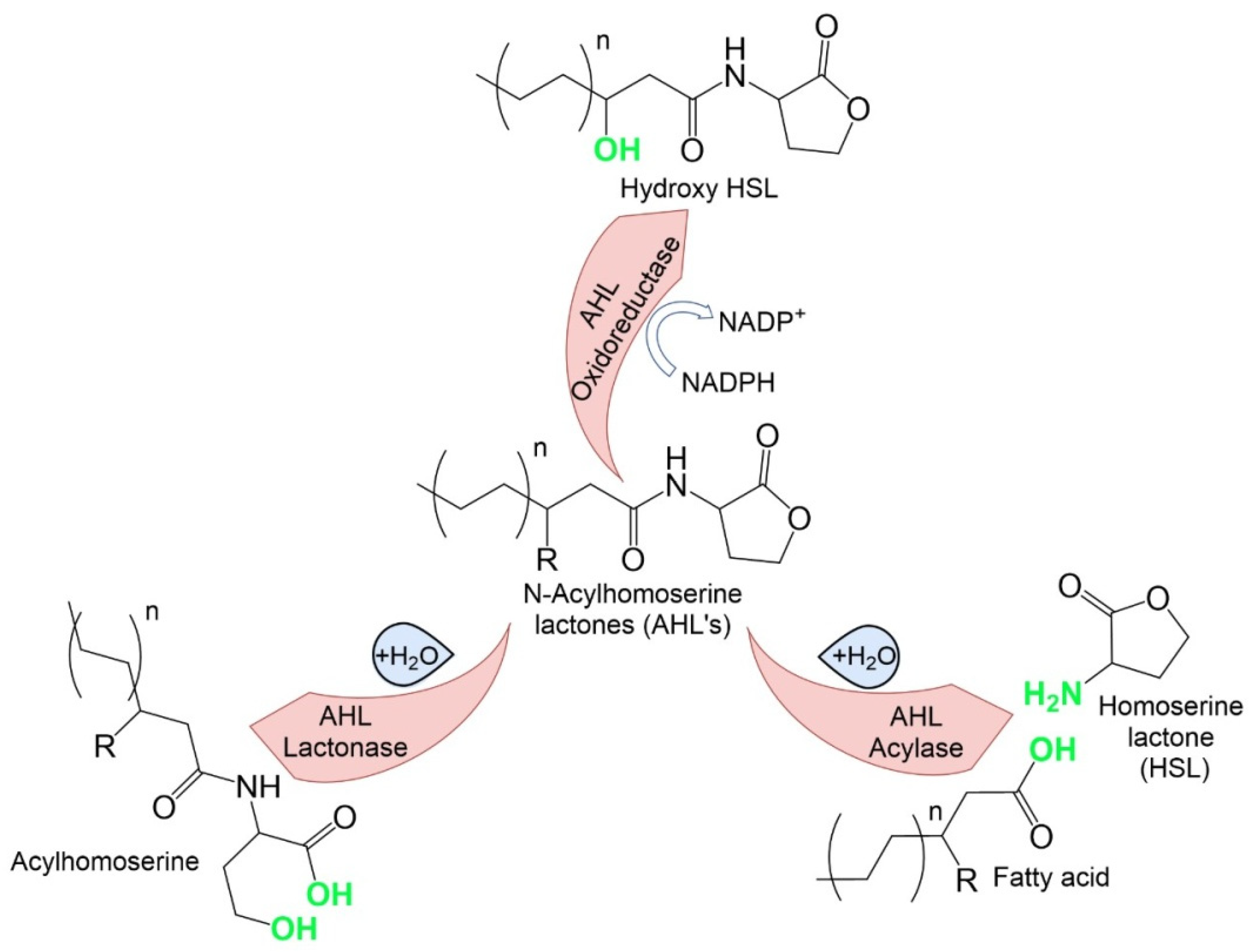
| Enzyme Categories | Mechanism of AHL Degradation |
|---|---|
| AHL lactonases | Hydrolysis of the AHL lactone ring to form the corresponding N-acyl homoserine. This process can also occur spontaneously in the presence of an alkaline pH and can be reversed when the pH is acidified |
| AHL acylases | Degradation of the AHL amide bond and generation of the corresponding free fatty acid and a lactone ring |
| AHL oxidoreductases | Modification (oxidation) and subsequent degradation of signal QS molecules |
| Strain | Mechanism | Against | References |
|---|---|---|---|
| L. acidophilus 30SC | AI-2 Inhibition | E. coli O157:H7 | [51] |
| L. plantarum SBR04MA | N-Hexanoyl-L-homoserine lactone (6-HSL) inhibition | Microbiota of activated sludge | [54] |
| L. plantarum CY-1 | HSL degradation (without details). | A. sobria | [67] |
| C. crustorum ZHG 2-1 | Dose-dependent degradation C4- and 3-oxo-C12- HSL | P. aeruginosa | [58] |
| Cell-free acidic supernatants L. lactis NCDC 309, L. rhamnosus MTCC 5897, L. rhamnosus MTCC 5857, L. fermentum MTCC 5898, L. acidophilus NCDC 15, L. delbrueckii subsp. lactis, L. plantarum NCDC 372, | Inhibition of biofilm formation, elastase, and expression of lasI and rhlI | P. aeruginosa | [59] |
| Cell-free neutral supernatants of some of the strains described above | Depending on the strain, both the lack of effect and the stimulation of some QS processes (an increase in the expression level of QS-related genes, elastase activity) were observed | P. aeruginosa | [59] |
| L. reuteri RC-14 | Inhibition of agr gene expression by cyclic dipeptides cyclo(L-Phe-L-Pro) and cyclo(L-Tyr-L-Pro) | S. aureus | [55] |
| L. plantarum, P. acidilactici | Reduce expression of genes cidA, icaA, dltB, agrA, sortaseA, and sarA involved in biofilm formation | S. aureus | [65] |
| L. plantarum M.2, L. curvatus B.67 | Inhibition of agr genes | Listeria monocytogenes | [65] |
| Heat-treated supernatant L. fermentum Lim2 | Inhibition of agr genes | C. difficile | [66] |
| Strain | Mechanism | Against | References |
|---|---|---|---|
| Rhodococcus pyridinivorans AI4 | 3-oxo-C6- N-acyl homoserine lactone degradation | Pectobacterium carotovorum | [71] |
| Glutamicibacter nicotianae AI5a | 3-oxo-C6- N-acyl homoserine lactone degradation | Pectobacterium carotovorum | [71] |
| Arthrobacter sp. IBN110 | 3-oxo-C6- N-acyl homoserine lactone degradation | Pectobacterium carotovorum N98 | [79] |
| Microbacterium testaceum StLB037 | C10-HSL degradation | Pectobacterium carotovorum | [76] |
| Micromonospora sp. TAV14 | Disruption of rhl-regulated QS | P. aeruginosa PAO1 | [77] |
| Streptomyces sp. NIO 10068, Streptomyces sp. NIO 10058 | Disruption of rhl-regulated QS | P. aeruginosa ATCC 27853 | [80] |
| Rhodococcus erythropolis R138 | 3 types of AHL degradation enzymes, AHL lactonase, acylase, and oxidoreductase | Pectobacterium atrosepticum CFBP 6276 | [81,82] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prazdnova, E.V.; Gorovtsov, A.V.; Vasilchenko, N.G.; Kulikov, M.P.; Statsenko, V.N.; Bogdanova, A.A.; Refeld, A.G.; Brislavskiy, Y.A.; Chistyakov, V.A.; Chikindas, M.L. Quorum-Sensing Inhibition by Gram-Positive Bacteria. Microorganisms 2022, 10, 350. https://doi.org/10.3390/microorganisms10020350
Prazdnova EV, Gorovtsov AV, Vasilchenko NG, Kulikov MP, Statsenko VN, Bogdanova AA, Refeld AG, Brislavskiy YA, Chistyakov VA, Chikindas ML. Quorum-Sensing Inhibition by Gram-Positive Bacteria. Microorganisms. 2022; 10(2):350. https://doi.org/10.3390/microorganisms10020350
Chicago/Turabian StylePrazdnova, Evgeniya V., Andrey V. Gorovtsov, Nikita G. Vasilchenko, Maxim P. Kulikov, Varvara N. Statsenko, Anna A. Bogdanova, Alexandr G. Refeld, Yaroslav A. Brislavskiy, Vladimir A. Chistyakov, and Michael L. Chikindas. 2022. "Quorum-Sensing Inhibition by Gram-Positive Bacteria" Microorganisms 10, no. 2: 350. https://doi.org/10.3390/microorganisms10020350