
Development of Real-Time Molecular Assays for the Detection of Wesselsbron Virus in Africa

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Virus Stocks and Samples Collection
2.3. Primers Design
2.4. Samples Preparation and RNA Extraction
2.5. Real-Time RT-qPCR Conditions
2.6. RPA Assay Conditions
2.7. Specificity Assessment
2.8. Sensitivity Assessment
2.8.1. Analytical Sensitivity
2.8.2. Sensitivity in Human Serum and L-15 Medium
2.8.3. Sensitivity on Clinical and Veterinary Samples
2.9. Assessment of the Impact of Genetic Diversity on Assays
2.10. Statistical Analysis
3. Results
3.1. Primers Selection
3.2. Analytical Specificity
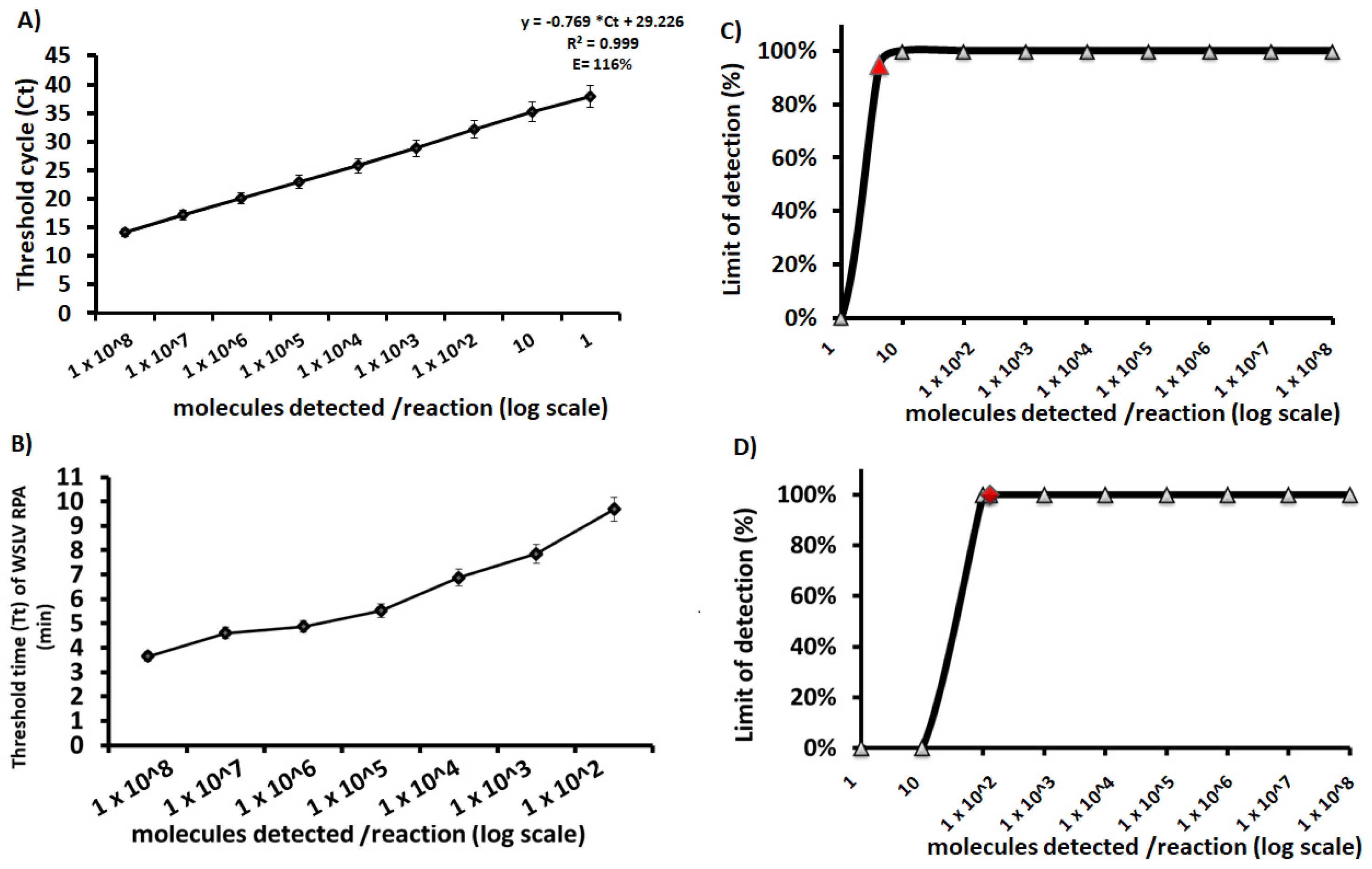
3.3. Analytical Sensitivity
3.4. Diagnostic Performances
3.5. Sensitivity in Human Serum and L-15 Medium
3.6. Sensitivity in Clinical and Veterinary Samples
3.7. In Silico Analysis of New Primers and Probes Sequences
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Strains | Virus | Place of Collection | Year of Collection | Species | Reference |
|---|---|---|---|---|---|
| ArD142157 | Wesselsbron | Senegal | 1995 | Aedes dalzieli | CRORA database |
| ArD142585 | Wesselsbron | Senegal | 1995 | Aedes dalzieli | CRORA database |
| ArD140166 | Wesselsbron | Mauritania | 1995 | Aedes vexans | CRORA database |
| ArD142730 | Wesselsbron | Senegal | 1995 | Aedes dalzieli | CRORA database |
| ArD142098 | Wesselsbron | Senegal | 1995 | Aedes dalzieli | CRORA database |
| ARA23495 | Wesselsbron | Côte d’Ivoire | 1987 | Culex perfuscus | CRORA database |
| ArD90431 | Wesselsbron | Senegal | 1988 | Aedes vittatus | CRORA database |
| ArD90535 | Wesselsbron | Senegal | 1988 | Aedes minutus | CRORA database |
| ArD142143 | Wesselsbron | Senegal | 1995 | Aedes dalzieli | CRORA database |
| ArD65233 | Wesselsbron | Senegal | 1995 | Aedes dalzieli | CRORA database |
| ArD142775 | Wesselsbron | Senegal | 1995 | Aedes dalzieli | CRORA database |
| ArD140179 | Wesselsbron | Mauritania | 1995 | Aedes vexans | CRORA database |
| ArD92269 | Wesselsbron | Senegal | 1988 | Cellia domicola | CRORA database |
| ArD142716 | Wesselsbron | Senegal | 1995 | Aedes dalzieli | CRORA database |
| ArD140184 | Wesselsbron | Mauritania | 1995 | Aedes vexans | CRORA database |
| ArA20858 | Wesselsbron | Côte d’Ivoire | 1982 | Culex perfuscus | CRORA database |
| SH88963 | Wesselsbron | Senegal | 1988 | Human | CRORA database |
| ArA22079B | Wesselsbron | Côte d’Ivoire | 1984 | Stegomyia africanus | CRORA database |
| ArD141023 | Wesselsbron | Senegal | 1995 | Aedes dalzieli | CRORA database |
| ArD285495 | Wesselsbron | Senegal | 2016 | Culex perfuscus | CRORA database |
| ArD90416 | Wesselsbron | Senegal | 1988 | Stegomyia luteocephalus | CRORA database |
| ArD140187 | Wesselsbron | Mauritania | 1995 | Aedes vexans | CRORA database |
| ArD90541 | Wesselsbron | Senegal | 1988 | Aedes minutus | CRORA database |
| ArD92276 | Wesselsbron | Senegal | 1988 | Cellia pharoensis | CRORA database |
| ArD140162 | Wesselsbron | Mauritania | 1995 | Aedes vexans | CRORA database |
| ArD140194 | Wesselsbron | Mauritania | 1995 | Aedes vexans | CRORA database |
| ArD85094 | Wesselsbron | Senegal | 1987 | Aedes dalzieli | CRORA database |
| ArA16523 | Wesselsbron | Côte d’Ivoire | 1985 | Culex perfuscus | CRORA database |
| ArA 23139 | Bagaza | Côte d’Ivoire | 1988 | Culex poicilipes | CRORA database |
| ArD76986 | WNV Lineage 1 | Senegal | 1990 | Culex poicilipes | KJ131500 |
| B956 | WNV Lineage 2 | Uganda | 1937 | Human | AY532665 |
| ArD96655 | WNV Koutango | Senegal | 1993 | Rhipicephalus guihoni | KJ131501 |
| ArD94343 | WNV new Lineage | Senegal | 1992 | Culex perfuscus | KJ131502 |
| New Guinea C | DENV2 | New Guinea | 1974 | Human | AF038403 |
| H-241 | DENV4 | Philippines | 1956 | Human | U18433 |
| ArAAMT/7 | Yellow fever | Côte d’Ivoire | 1973 | Aedes africanus | CRORA database |
| MR766 | Zika | Uganda | 1947 | Rhesus Monkey | AY632535 |
| SAAR1776 | Usutu | South Africa | 1959 | Culex neavei | AY453412 |
| ArB1803 | Usutu subtype | CAR | 1969 | Culex perfuscus | KC754958 |
| Assay | Primers | EU707555_ SAH177_SA_1955 | MK163943_SA999-10_SA_2010 | JN226796_SA_1997 | KY056258_ WSLV-IP259570/SEN/2013_SN_2013 | KY056256_ WSLV-IP248525/SEN/2013_SN_2013 | KY056257_ WSLV-IP262451/SEN/2014_SN_2014 |
|---|---|---|---|---|---|---|---|
| RT-qPCR | WSBFOW | 5% | 5% | 0% | 0% | 0% | 0% |
| WSBPROBE | 15% | 0% | 5% | 5% | 5% | 5% | |
| WSBREV | 5% | 0% | 0% | 0% | 0% | 0% | |
| RT-RPA | RF2 | 0% | 0% | 0% | 0% | 0% | 0% |
| exoProbe | 8% | 2% | 2% | 2% | 2% | 2% | |
| RR2 | 6% | 3% | 3% | 0% | 0% | 0% |

References
- Karabatsos, N. (Ed.) International Catalogue of Arboviruses, Including Certain Other Viruses of Vertebrates, 3rd ed.; The American Journal of Tropical Medicine and Hygiene: San Antonio, TX, USA, 1985; Volume 91, pp. 179–181. [Google Scholar]
- Weiss, K.E.; Haig, D.A.; Alexander, R.A.; Alexander, R.A.; Clark, R.; Louw, J.G.; De Kock, V.E. Wesselsbron virus-A virus not previously described, associated with abortion in domestic animals. Onderstepoort Vet. J. 1956, 27, 183–195. [Google Scholar]
- Coetzer, J.A.W.; Barnard, B.J.H. Hydrops amnii in sheep associated with hydranencephaly and arthrogryposis with Wesselsbron disease and Rift Valley fever viruses as ethological agents. Onderstepoort J. Vet. Res. 1977, 44, 119–126. [Google Scholar]
- Mushi, E.Z.; Binta, M.G.; Raborokgwe, M. Wesselsbron disease virus associated with abortions in goats in Botswana. J. Vet. Diagn. Invest. 1998, 10, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venter, M.M.; Human, S.; van Eeden, C.; van Niekerk, S.; Williams, J.; Steyl, J.; Gerdes, T.; Swanepoel, R. The role of zoonotic vector-borne viruses as neurological pathogens in horses and wildlife in South Africa. In Proceedings of the 9th Annual Congress of the South African Society for Veterinary Epidemiology and Preventive Medicine, Farm Inn, Pretoria, South Africa, 18–20 August 2010; pp. 85–89. [Google Scholar]
- Diagne, M.M.; Faye, M.; Faye, O.; Sow, A.; Balique, F.; Sembène, M.; Granjon, L.; Handschumacher, P.; Faye, O.; Diallo, M.; et al. Emergence of Wesselsbron virus among black rat and humans in Eastern Senegal in 2013. One Health 2017, 3, 23–28. [Google Scholar] [CrossRef]
- Swanepoel, R. Wesselsbron virus disease. In The Arboviruses Epidemiology and Ecology; Monath, T.P., Ed.; CRC Press: Boca Raton, FL, USA, 1989; Volume 5. [Google Scholar]
- Jupp, P. Mosquitoes of Southern Africa: Culicinae and Toxorhynchitinae; Ekogilde Publishers: Hartebeespoort, South Africa, 1996. [Google Scholar]
- Kokernot, R.H.; Smithburn, K.C.; Paterson, H.E.; De Meillon, B. Further isolations of Wesselsbron virus from mosquitoes. S. Afr. Med. J. 1960, 34, 871–874. [Google Scholar] [PubMed]
- Kokernot, R.H.; Paterson, H.E.; De Meillon, B. Studies on the transmission of Wesselsbron virus by Aedes (Ochlerotatus) caballus (Theo). S. Afr. Med. J. 1958, 32, 546–548. [Google Scholar] [PubMed]
- Diallo, M.; Nabeth, P.; Ba, K.; Sall, A.A.; Ba, Y.; Mondo, M.; Girault, L.; Abdalahi, M.O.; Mathiot, C. Mosquito vectors of the 1998-1999 outbreak of Rift Valley Fever and other arboviruses (Bagaza, Sanar, Wesselsbron and West Nile) in Mauritania and Senegal. Med. Vet. Entomol. 2005, 19, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Durand, J.-P.; de Camaret, X.; Zeller, H.; Sall, A.; Tock, F.; Czarnecki, E.; Régis, R.; Guesdon, S.; Tolou, H. First isolation of Wesselsbron virus in Chad. Virologie 2001, 5, 305–306. [Google Scholar]
- McIntosh, B.M. The Epidemiology of Arthropod-Borne Viruses in Southern Africa. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, 1980; pp. 144–159. [Google Scholar]
- Weyer, J.; Thomas, J.; Leman, P.A.; Grobbelaar, A.A.; Kemp, A.; Paweska, J.T. Human cases of Wesselsbron disease, South Africa 2010–2011. Vector-Borne Zoonotic Dis. 2013, 13, 330–336. [Google Scholar] [CrossRef]
- Baba, S.S.; Fagbami, A.H.; Ojeh, C.K.; Olaleye, O.D.; Omilabu, S.A. Wesselsbron virus antibody in domestic animals in Nigeria: Retrospective and prospective studies. New Microbiol. 1995, 18, 151–162. [Google Scholar]
- Barnard, B.J. Antibodies against some viruses of domestic animals in southern African wild animals. Onderstepoort J. Vet. Res. 1997, 64, 95–110. [Google Scholar] [PubMed]
- Kokernot, R.H.; Szlamp, E.L.; Levitt, J.; McIntosh, B.M. Survey for antibodies against arthropod-borne viruses in the sera of indigenous residents of the Caprivi strip and Bechuanaland protectorate. Trans. R. Soc. Trop. Med. Hyg. 1965, 59, 553–562. [Google Scholar] [CrossRef]
- Smithburn, K.C.; Kokernot, R.H.; Heymann, C.S.; Weinbren, M.P.; Zentkowsky, D. Neutralizing antibodies for certain viruses in the sera of human beings residing in northern Natal. S. Afr. Med. J. 1959, 33, 555–561. [Google Scholar] [PubMed]
- Gould, E.A.; de Lamballerie, X.; de A Zanotto, P.M.; Holmes, E.C. Origins, evolution, and vector/host coadaptations within the genus Flavivirus. Adv. Virus Res. 2003, 59, 277–314. [Google Scholar]
- Henchal, E.A.; Gentry, M.K.; McCown, J.M.; Brandt, W.E. Dengue virus-specific and flavivirus group determinants identified with monoclonal antibodies by indirect immunofluorescence. Am. J. Trop. Med. Hyg. 1982, 31, 830–836. [Google Scholar] [CrossRef]
- Huhtamo, E.; Putkuri, N.; Kurkela, S.; Manni, T.; Vaheri, A.; Vapalahti, O.; Uzcategui, N.Y. Characterization of a novel flavivirus from mosquitoes in northern Europe that is related to mosquito-borne flaviviruses of the tropics. J. Virol. 2009, 83, 9532–9540. [Google Scholar] [CrossRef] [Green Version]
- Moureau, G.; Temmam, S.; Gonzalez, J.P.; Charrel, R.N.; Grard, G.; de Lamballerie, X. A real-time RT-PCR method for the universal detection and identification of flaviviruses. Vector-Borne Zoonotic Dis. 2007, 7, 467–477. [Google Scholar] [CrossRef]
- Grard, G.; Moureau, G.; Charrel, R.N.; Lemasson, J.J.; Gonzalez, J.P.; Gallian, P.; Gritsun, T.S.; Holmes, E.C.; Gould, E.A.; de Lamballerie, X. Genetic characterization of tick-borne flaviviruses: New insights into evolution, pathogenetic determinants and taxonomy. Virol 2007, 361, 80–92. [Google Scholar] [CrossRef]
- Scaramozzino, N.; Crance, J.M.; Jouan, A.; DeBriel, D.A.; Stoll, F.; Garin, D. Comparison of flavivirus universal primer pairs and development of a rapid, highly sensitive heminested reverse transcription-PCR assay for detection of flaviviruses targeted to a conserved region of the NS5 gene sequences. J. Clin. Microbiol. 2001, 39, 1922–1927. [Google Scholar] [CrossRef] [Green Version]
- Kuno, G.; Chang, G.J.; Tsuchiya, K.R.; Karabatsos, N.; Cropp, C.B. Phylogeny of the genus Flavivirus. J. Virol. 1998, 72, 73–83. [Google Scholar] [CrossRef] [Green Version]
- University of Iowa Principles for the Care and Use of Laboratory Animals (IACUC). AVMA Guidelines for the Euthanasia of Animals, 2013 ed.; American Veterinary Medical Association: Schaumburg, IL, USA, 2013. [Google Scholar]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [Google Scholar]
- Digoutte, J.P.; Calvo-Wilson, M.A.; Mondo, M.; Traore-Lamizana, M.; Adam, F. Continuous cell lines and immune ascitic fluid pools in arbovirus detection. Res. Virol. 1992, 143, 417–422. [Google Scholar] [CrossRef]
- Patel, P.; Landt, O.; Kaiser, M.; Faye, O.; Koppe, T.; Lass, U.; Sall, A.A.; Niedrig, M. Development of one-step quantitative reverse transcription PCR for the rapid detection of flaviviruses. Virol. J. 2013, 10, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; The UGENE Team. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Nattanmai, S.; Kramer, L.D.; Bernard, K.A.; Tavakoli, N.P. A duplex real-time reverse transcriptase polymerase chain reaction assay for the detection of California serogroup and Cache Valley viruses. Diagn. Microbiol. Infect. Dis. 2009, 65, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Abd El Wahed, A.; Patel, P.; Faye, O.; Thaloengsok, S.; Heidenreich, D.; Matangkasombut, P.; Manopwisedjaroen, K.; Sakuntabhai, A.; Sall, A.A.; Hufert, F.T.; et al. Recombinase polymerase amplification assay for rapid diagnostics of dengue infection. PLoS ONE 2015, 10, e0129682. [Google Scholar] [CrossRef]
- Faye, M.; Abd El Wahed, A.; Faye, O.; Kissenkötter, J.; Hoffmann, B.; Sall, A.A.; Faye, O. A recombinase polymerase amplification assay for rapid detection of rabies virus. Sci. Rep. 2021, 11, 3131. [Google Scholar] [CrossRef]
- Faye, M.; Dacheux, L.; Weidmann, M.; Diop, S.A.; Loucoubar, C.; Bourhy, H.; Sall, A.A.; Faye, O. Development and validation of sensitive real-time RT-PCR assay for broad detection of rabies virus. J. Virol. Methods 2017, 243, 120–130. [Google Scholar] [CrossRef] [Green Version]
- Schlottau, K.; Freuling, C.M.; Müller, T.; Beer, M.; Hoffmann, B. Development of molecular confirmation tools for swift and easy rabies diagnostics. Virol. J. 2017, 14, 184. [Google Scholar] [CrossRef] [Green Version]
- Nagy, A.; Jiřinec, T.; Jiřincová, H.; Černíková, L.; Havlíčková, M. In silico re-assessment of a diagnostic RT-qPCR assay for universal detection of Influenza A viruses. Sci. Rep. 2019, 9, 1630. [Google Scholar] [CrossRef] [PubMed]
- Villinger, J.; Mbaya, M.K.; Ouso, D.; Kipanga, P.N.; Lutomiah, J.; Masiga, D.K. Arbovirus and insect-specific virus discovery in Kenya by novel six genera multiplex high-resolution melting analysis. Mol. Ecol. Resour. 2017, 17, 466–480. [Google Scholar] [CrossRef] [PubMed]
- Abd El Wahed, A.; El-Deeb, A.; El-Tholoth, M.; Abd El Kader, H.; Ahmed, A.; Hassan, S.; Hoffmann, B.; Haas, B.; Shalaby, M.A.; Hufert, F.T.; et al. A portable reverse transcription recombinase polymerase amplification assay for rapid detection of foot-and-mouth disease virus. PLoS ONE. 2013, 8, e71642. [Google Scholar]
- Oymans, J.; van Keulen, L.; Wichgers Schreur, P.J.; Kortekaas, J. Early Pathogenesis of Wesselsbron Disease in Pregnant Ewes. Pathogens 2020, 9, 373. [Google Scholar] [CrossRef]
| Name | Sequence 5′-3′ | Protein | Position a | GC% | Tm b | Product Size |
|---|---|---|---|---|---|---|
| WSBFOW | GAGGACCAACGGAAAGTGTT | NS3 | 6146–6165 | 50.00 | 59.04 | 231 |
| WSBREV c | ACTGCATACCCTGGTGTCAA | NS3 | 6357–6376 | 50.00 | 59.01 | |
| WSBPROBE c | 6FAM-TCGCAACCTGCCATGACAGC--BBQ | NS3 | 6202–6221 | 60.00 | 68.05 | |
| RF2 | GGAACAGCAGTGATGCAGTAAAAGTTACAAC | Envelope | 1895–1926 | 50.00 | 126 | |
| RR2 c | GACGCAGCAATAGGGTTGGTCGTGATGAGCT | Envelope | 1991–2022 | 50.00 | ||
| exoProbe c | CCCACGGTTCTCTGTTCCTGCCATGGAG dT-BHQ1-THF–dT-FAM GCTGCAATCACTGGA-Ph | Envelope | 1940–1990 | 60.00 |
| Strains | Virus | PAN-FLAVIVIRUS RT-qPCR | WSLV RT-qPCR | WSLV RT-RPA | |||
|---|---|---|---|---|---|---|---|
| Mean Ct Value a | SD | Mean Ct Value a | SD | Mean Tt Value b | SD | ||
| ArD142157 | Wesselsbron | 25.80 | 0.01 | 26.18 | 0.58 | 4.21 | 0.21 |
| ArD142585 | Wesselsbron | 21.81 | 0.02 | 23.79 | 0.29 | 4.99 | 0.11 |
| ArD140166 | Wesselsbron | 26.07 | 0.24 | 24.81 | 0.03 | 3.30 | 0.11 |
| ArD142730 | Wesselsbron | 29.11 | 0.18 | 27.52 | 0.30 | 4.43 | 0.40 |
| ArD142098 | Wesselsbron | 22.93 | 0.63 | 23.12 | 0.27 | 4.83 | 0.04 |
| ARA23495 | Wesselsbron | 28.49 | 0.48 | 19.36 | 0.15 | 5.33 | 0.02 |
| ArD90431 | Wesselsbron | 24.33 | 0.24 | 15.18 | 0.01 | 3.56 | 0.47 |
| ArD90535 | Wesselsbron | 25.06 | 0.23 | 15.10 | 0.07 | 3.21 | 0.42 |
| ArD142143 | Wesselsbron | 31.41 | 0.26 | 26.36 | 0.38 | 3.65 | 0.02 |
| ArD65233 | Wesselsbron | 23.69 | 0.06 | 14.93 | 0.04 | 3.54 | 0.53 |
| ArD142775 | Wesselsbron | 24.67 | 0.22 | 22.77 | 0.44 | 3.21 | 0.06 |
| ArD140179 | Wesselsbron | 28.79 | 0.06 | 15.04 | 0.01 | 3.37 | 0.00 |
| ArD92269 | Wesselsbron | 25.54 | 0.07 | 15.89 | 0.13 | 4.94 | 0.23 |
| ArD142716 | Wesselsbron | 24.41 | 0.01 | 24.77 | 0.04 | 4.83 | 0.32 |
| ArD140184 | Wesselsbron | 21.49 | 0.03 | 14.71 | 0.01 | 4.15 | 0.76 |
| ArA20858 | Wesselsbron | 29.98 | 0.30 | 24.57 | 0.01 | 5.49 | 0.38 |
| SH88963 | Wesselsbron | 24.31 | 0.15 | 19.75 | 0.13 | 4.84 | 0.09 |
| ArA22079B | Wesselsbron | 22.69 | 0.04 | 24.67 | 0.20 | 5.49 | 0.64 |
| ArD141023 | Wesselsbron | 22.34 | 0.18 | 25.79 | 0.43 | 5.42 | 0.02 |
| ArD285495 | Wesselsbron | 14.78 | 0.14 | 27.59 | 0.13 | 5.78 | 0.06 |
| ArD90416 | Wesselsbron | 24.89 | 0.23 | 15.26 | 0.01 | 5.65 | 0.47 |
| ArD140187 | Wesselsbron | 27.77 | 0.05 | 15.45 | 0.01 | 5.39 | 0.02 |
| ArD90541 | Wesselsbron | 24.13 | 0.20 | 15.37 | 0.02 | 5.50 | 0.57 |
| ArD92276 | Wesselsbron | 25.58 | 0.04 | 15.72 | 0.01 | 4.94 | 0.06 |
| ArD140162 | Wesselsbron | 23.43 | 0.23 | 15.52 | 0.07 | 4.21 | 0.56 |
| ArD140194 | Wesselsbron | 21.68 | 0.04 | 14.71 | 0.04 | 4.83 | 0.08 |
| ArD85094 | Wesselsbron | 21.96 | 0.11 | 14.92 | 0.01 | 4.85 | 0.19 |
| ArA16523 | Wesselsbron | 22.29 | 0.05 | 14.80 | 0.02 | 3.44 | 0.08 |
| ArA 23139 | Bagaza | 26.11 | 0.02 | − | – | – | – |
| ArD76986 | WNV Lineage 1 | 23.09 | 0.01 | – | – | – | – |
| B956 | WNV Lineage 2 | 16.59 | 0.01 | – | – | – | – |
| ArD96655 | WNV Koutango | 23.25 | 0.01 | – | – | – | – |
| ArD94343 | WNV new Lineage | 19.39 | 0.05 | – | – | – | – |
| New Guinea C | DENV2 | 22.98 | 0.00 | – | – | – | – |
| H-241 | DENV4 | 25.65 | 0.01 | – | – | – | – |
| ArAAMT/7 | Yellow fever | 24.80 | 0.03 | – | – | – | – |
| MR766 | Zika | 27.88 | 0.01 | – | – | – | – |
| SAAR1776 | Usutu | 28.59 | 0.04 | – | – | – | – |
| ArB1803 | Usutu subtype | 19.83 | 0.02 | – | – | – | – |
| Strains | GenBank Accession Number | Species | Sample Type | WSLV RT-qPCR | WSLV RT-RPA | |||
|---|---|---|---|---|---|---|---|---|
| Mean Ct Value a | Mean Tt Value b | SD | Mean Tt Value b | SD | ||||
| WSLV-IP262451/SEN/2014 | KY056257 | human | Serum | 26.18 | 43.78 | 0.02 | 7.30 | 0.70 |
| WSLV-IP248525/SEN/2013 | KY056256 | human | Serum | 23.79 | 45.88 | 015 | 7.60 | 0.18 |
| WSLV-IP259570/SEN/2013 | KY056258 | rodent | Brain tissues | 24.81 | 43.87 | 1.37 | 7.50 | 0.15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faye, M.; Seye, T.; Patel, P.; Diagne, C.T.; Diagne, M.M.; Dia, M.; Thiaw, F.D.; Sall, A.A.; Faye, O. Development of Real-Time Molecular Assays for the Detection of Wesselsbron Virus in Africa. Microorganisms 2022, 10, 550. https://doi.org/10.3390/microorganisms10030550
Faye M, Seye T, Patel P, Diagne CT, Diagne MM, Dia M, Thiaw FD, Sall AA, Faye O. Development of Real-Time Molecular Assays for the Detection of Wesselsbron Virus in Africa. Microorganisms. 2022; 10(3):550. https://doi.org/10.3390/microorganisms10030550
Chicago/Turabian StyleFaye, Martin, Thiané Seye, Pranav Patel, Cheikh Tidiane Diagne, Moussa Moise Diagne, Moussa Dia, Fatou Diène Thiaw, Amadou Alpha Sall, and Ousmane Faye. 2022. "Development of Real-Time Molecular Assays for the Detection of Wesselsbron Virus in Africa" Microorganisms 10, no. 3: 550. https://doi.org/10.3390/microorganisms10030550
APA StyleFaye, M., Seye, T., Patel, P., Diagne, C. T., Diagne, M. M., Dia, M., Thiaw, F. D., Sall, A. A., & Faye, O. (2022). Development of Real-Time Molecular Assays for the Detection of Wesselsbron Virus in Africa. Microorganisms, 10(3), 550. https://doi.org/10.3390/microorganisms10030550