Abstract
Bacterial volatiles play important roles in mediating beneficial interactions between plants and their associated microbiota. Despite their relevance, bacterial volatiles are mostly studied under laboratory conditions, although these strongly differ from the natural environment bacteria encounter when colonizing plant roots or shoots. In this work, we ask the question whether plant-associated bacteria also emit bioactive volatiles when growing on plant leaves rather than on artificial media. Using four potato-associated Pseudomonas, we demonstrate that potato leaves offer sufficient nutrients for the four strains to grow and emit volatiles, among which 1-undecene and Sulfur compounds have previously demonstrated the ability to inhibit the development of the oomycete Phytophthora infestans, the causative agent of potato late blight. Our results bring the proof of concept that bacterial volatiles with known plant health-promoting properties can be emitted on the surface of leaves and warrant further studies to test the bacterial emission of bioactive volatiles in greenhouse and field-grown plants.
1. Introduction
Plants are densely colonized from the roots to the shoots by a plethora of microbes and increasing evidence indicates that these microbiota play essential roles for the growth, development and defense of its plant host [1,2,3,4]. Bacteria belonging to the genus Pseudomonas are an important component of this diverse plant microbiota and have been shown to display many different plant-beneficial properties, and, in particular, to be very well equipped to help their host defend itself against disease-causing agents, e.g., through induction of systemic resistance, niche competition or the production of antimicrobial compoundsac [5,6]. Among the wide variety of chemical weapons encoded in the genomes of many plant-associated Pseudomonas strains, volatile organic compounds (VOCs) have recently attracted increased attention as short- and long-distance bioactive compounds with strong inhibiting effects against various disease-causing agents [7,8,9,10,11,12]. Although initially demonstrated on the model plant Arabidopsis [13,14], the bioactivity of these volatiles has meanwhile also been reported in many plants of agronomical relevance such as grapevines, peppers, tomatoes or potatoes [15,16,17,18]. This latter crop is mainly threatened by the oomycete Phytophthora infestans (Mont.) de Bary, the causative agent of late blight, which leads to high economic losses worldwide. Over 150 years after the Irish Famine partly caused by this pathogen, we are currently still struggling to control this disease and rely to this end on repeated applications of synthetic or copper-based fungicides with deleterious side-effects on environmental and human health [19,20,21].
In an attempt to find alternative solutions to control potato late blight, we have previously isolated and characterized many members of its microbiota [9]. From the assembled strain collection, Pseudomonas strains isolated from both the roots and the shoots were the emitters of the most bioactive volatiles, and we identified 1-Undecene as well as diverse Sulfur-containing volatiles (S-VOCs), which were particularly efficient in inhibiting different life stages of P. infestans [9,16,22]. Some of these S-VOCs were shown in other studies to contribute to plant Sulfur nutrition or to induced systemic resistance, highlighting their versatile nature, which could be of great interest for practical applications [23,24].
However, so far, all studies dealing with the identification and characterization of volatile emissions by plant-associated bacteria have been carried out in artificial laboratory media, which represent very different growth conditions from those encountered by bacteria on their plant host. This is especially true for the phyllosphere environment, which in contrast to the rhizosphere is very poor in the organic carbon sustaining the growth of heterotrophic bacteria such as Pseudomonas. Therefore, the aim of this study was to assess whether such bioactive volatiles would also be emitted by bacteria growing on leaves without any external supply of nutrients.
Using leaves of in vitro grown potato plants inoculated or not with different Pseudomonas strains and taking advantage of a specific volatile analysis method allowing sensitive detection of S-VOCs, we provide evidence that potato leaf-colonizing bacteria emit detectable quantities of volatiles with known anti-Phytophthora activity. Although the system used in the present study lies still far away from the reality of field-grown potato plants, our results bring the important proof of concept that the emission of volatile compounds by bacteria is not a laboratory artefact but occurs also in the nutrient-poor leaf surface environment.
2. Materials and Methods
2.1. Cultivation of Microbial Strains and of Plants
2.1.1. Bacterial Strains
The four Pseudomonas strains used for this study were isolated in 2015 from field-grown potatoes as described in [9,25], two from the roots (Pseudomonas donghuensis R32, P. chlororaphis R47) and two from the shoots (Pseudomonas sp. S04, Pseudomonas sp. S35). We selected these strains because they differed in their protective activity against P. infestans and in their ability to emit VOCs on laboratory media. The strains were kept in 25% (v/v) glycerol for long-term storage and passaged weekly on 1.5% agar (w/v; Erne, Switzerland) LB medium (20 g/L; Applichem, Schaffhausen, Switzerland) or ABG medium [26] for routine use.
2.1.2. Phytophthora Infestans
The isolate P. infestans was originally isolated by H. Krebs (Agroscope, Switzerland) in 2001. It was later characterized as an A2 mating type isolate belonging to the lineage Blue13 [27]. P. infestans was maintained on pea growth medium [28] or rye agar [9] on Petri dishes sealed with Parafilm (BEMIS Flexible Packaging, Neenah, WI, USA) and incubated at 19 °C in the dark.
2.1.3. Potato Plantsa
Sterile in vitro potato plantlets (var. Désirée) were obtained from E. Droz (Agroscope, Switzerland) and cultivated on CMS medium, a minimal medium containing per liter 20 g sucrose, 8 g Agar-Agar (Kobe), 230 mg NH4H2PO4, 370 mg MgSO4 • 7 H2O, 1.21 g KNO3, 708 mg Ca(NO3)2 • 4 H2O, 37 mg Na2EDTA, 9.3 mg FeSO4 • 7 H2O, 8.15 mg KI, and 1 mL CMS solution (containing per liter 11.1 g MnSO4 • 4 H2O, 8.41 g MnSO4 • H2O, 2.87 g ZnSO4 • 7 H2O, 0.25 g CuSO4 • 5 H2O, 0.25 g Na2MoO4 • 2 H2O and 25 mg CoCl2 • 6 H2O). For propagation, one internode with an intact leaf was taken and placed into a new culture tube containing 20 mL of CMS medium. Plants were grown for 30 d in a culture room (16H/8H D/N; 20 °C). After 30 d, the upper part of the plant with the four last expanded leaves was collected for the inoculation and volatile analysis.
2.2. Inoculation of Plants and Collection of Volatiles
Sterile 20 mL GC headspace screw vials were filled with 4 mL of water agar (WA). To prepare the bacterial culture, four colonies of an overnight culture (24 °C) of each strain were resuspended in 500 µL 0.9% NaCl (w/v) and 100 µL were plated on ABG medium. Plates were incubated overnight at 24 °C. The day after, the bacterial lawn was resuspended in 10 mL 0.9% NaCl (w/v), washed two times with 0.9% NaCl (w/v) and one additional time with sterile deionised water. After these three washing steps aiming to remove traces of the cultivation medium, bacteria were finally resuspended in sterile deionised water. Sporangia of a 3-week-old P. infestans culture were harvested with sterile deionised water.
Potato leaves were dipped in (i) sterile water (control), (ii) washed bacterial solution at OD600 = 1 of each strain separately, (iii) a sporangial solution of 125,000 sporangia/mL, or (iv) a mixed solution of bacteria (OD600 = 1) and sporangia (125,000/mL). After dipping, leaves were carefully deposited on the top of the WA in the screw vials, caps were applied and screwed by letting one quarter turn open to allow for air penetration. Vials were kept in the dark at 20 °C for two days, after which they were completely closed and left for two additional days at 20 °C in the dark until volatile collection (see below). The biomass of each plant sample was determined by weighing the vial before and after addition of the leaves.
2.3. Analysis of the Emitted Volatiles by HS-SPME GC-PFPD/MS
Samples were extracted using a 2 cm solid phase microextraction (SPME) carboxen/polydimethylsiloxane (CAR/PDMS) 85 μm StableFlex fiber (Supelco, Bellefonte, PA, USA). The fiber was conditioned according to the supplier’s recommendations (300 °C for 60 min under a nitrogen flow in a conditioner SPME station). The analyses were realized using an MPS2 autosampler equipped with Maestro1 software, V.1.4.8.14/3.5 (Gerstel, Sursee, Switzerland), a Trace GC Ultra GC coupled with a DSQ II mass selective detector (MSD) (Thermo Finnigan, Milan, Italy), and a pulsed flame photometric detector (PFPD) (OI Analytical, College Station, TX, USA) as a specific detector for sulfur compounds, as described in [29].
The headspace was extracted for 120 min at 60 °C with an agitation rate of 250 rpm without incubation. Bound volatiles were desorbed for 1 min at 250 °C in the injector, which was in the splitless mode for 30 s, and then the split valve was opened (split flow = 80 mL/min). VOCs were separated on a TRB-FFAP fused silica capillary column (100% polyethylene glycol PEG with nitroterephthalic acid, bonded and cross-linked, 30 m × 0.32 mm, 1.0 μm film, Teknokroma, Barcelona, Spain) with helium as the carrier gas at a constant flow of 2.1 mL/min (37 cm/s). The oven temperature was programmed as follows: 4 min at 40 °C, then heated to 220 °C at a rate of 5 °C/min, with a final hold time of 5 min. The settings of the PFPD were as follows: 250 °C, voltage at 540 V, ignitor current at 2.6 A, trigger level at 300, range at 10, and attenuation at 32 with flow rates of air1 at 17 mL/min, H2 at 14 mL/min, and air2 at 10 mL/min. The MS settings were as follows: transfer line at 230 °C, source temperature at 230 °C, and analytes monitored in SCAN mode between 30 and 150 amu without solvent delay.
The detector response signals were integrated using the Xcalibur 2.0.7 software (Thermo Fisher Scientific AG, Reinach, Switzerland). The NIST/EPA/NIH mass spectral library (NIST11) version 2.0 g (NIST, Gaithersburg, MD, USA) was used for peak identification. The PFPD detector was configured to detect and trace only the volatile organic sulfur compounds (S-VOCs) in the chromatogram. Quantitative ion peak area was integrated in order to evaluate the abundance of each VOC. This amount was then divided by the fresh weight of the leaves contained in each vial to estimate the volatile emission per unit of biological material and allow comparisons between the different samples.
2.4. Data and Statistical Analysis
The leaf inoculation and volatile collection experiment was performed in three independent biological replicates containing each two technical repetitions. In each independent experiment, the mean of the two technical repetitions was calculated for each treatment. ANOVA of the VOCs quantitative ion peak area/g in conjunction with Tukey’s test (unadjusted p < 0.05) was applied using the software package R (2.8.0). Data were centered and assigned to different qualitative groups (R32, R47, S04, S35, non-inoculated). PCA was performed using the ‘FactoMineR’ function of the R-package.
3. Results
3.1. Inoculation with Pseudomonas strains Leads to Major Changes in Potato Leaf Volatile Emissions
In order to characterize the volatile blends emitted by potato-associated Pseudomonas when growing on their host plant rather than on artificial laboratory media, four selected Pseudomonas strains (R32, R47, S04 and S35) were inoculated on sterile potato leaves. Since we were also interested to see whether concomitant inoculation and infection with Phytophthora infestans would alter the volatile profiles of the strains, we performed the analysis on leaves inoculated with the bacterial strains alone or together with P. infestans. The first question was whether the inoculated strains would be able to survive and grow on sterile potato leaves. Our measurement of bacterial density a few hours and five days after inoculation revealed that the four strains were able not only to survive, but to grow well on the leaves, reaching ca. 1011 cells per leaf (Table S1), which was a prerequisite for their emission of VOCs. To analyse the VOCs they emitted, we used a combination of two detection methods: PFPD to identify Sulfur-containing volatile organic compounds (S-VOCs) with increased selectivity, and mass spectrometry (MS) to identity all detected compounds. A total of 75 VOCs were detected in the samples including 13 S-VOCs detected specifically with the PFPD detector.
An ANOVA analysis on these 75 detected VOCs revealed 32 VOCs which were differentially emitted by the leaves depending on their inoculation status (differing either between non-inoculated and inoculated leaves or between leaves inoculated with different strains, see Table S2 for details). In contrast, only five VOCs were emitted differentially in leaves infected vs. non-infected with P. infestans (Table 1).

Table 1.
Differentially emitted VOCs in samples inoculated with bacterial strains P. donghuensis R32, P. chlororaphis R47, Pseudomonas sp. S04, Pseudomonas sp. S35 in presence and absence of P. infestans (n = 3). Only differentially expressed VOCs according to bacterial strain inoculation or infection status with P. infestans are presented.
A principal component analysis (PCA) was then performed on these 32 differentially emitted VOCs to identify the most discriminative factors between the different samples. As expected from the results shown in Table 1, PCA did not discriminate between leaves inoculated with P. infestans and uninoculated leaves (Supplementary Figure S1). For this reason, PCA is presented with only the factor strain (Figure 1). The main principal component (PC1) accounted for 38% of the variance, while PC2 accounted for 16% of the variation.
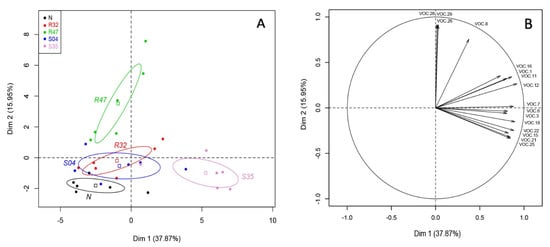
Figure 1.
Principal component analysis of the volatiles emitted by leaves inoculated with Pseudomonas strains. (A) Score Plot of PC1 and PC2 of potato leaves non-inoculated (N) or inoculated with different Pseudomonas strains (R32, R47, S04, S35) following PCA. Superimposed on the plot are mean scores on the components for qualitative variables that are included in the PCA command and the colored ellipses illustrating the 95% confidence interval with each score. (B) Results of principal components analysis (PCA): projection on two first factors (PC1 and PC2) of the VOCs with a cosinus square superior to 0.6. VOC.1: 2-methyl-3-methylthiofuran; VOC.3: dimethyl disulfide, VOC.6: dimethyl trisulfide, VOC.7: methanethiol, VOC.8: thiocyanic methylester, VOC.11: 3-pentanone, VOC.12: Unknown 5C, VOC.15: hexanal, VOC.16: 1-undecene, VOC.18: 2-heptanone, VOC.21: (Z)-2-heptenal, VOC.22: 6-octene-2-one, VOC.25: (E)-2-octenal, VOC.26: putative, longiverbenone or isomer, VOC.28: Cyclobutane, tetrakis (1-methylethylidene)-, VOC.29: (2,6,6-trimethylcyclohexen-1-yl) methylsulfonylbenzene.
A clear difference could be observed in the VOCs emitted by leaves inoculated with Pseudomonas sp. S35 on PC1 (Figure 1A), while PC2 discriminated non-inoculated leaves and those inoculated with S04, R32 and S35 from those inoculated with P. chlororaphis R47. Figure 1B shows the vectors of the 16 VOCs that were best represented on the projection of the two axis and therefore likely responsible for the differences in volatilome profiles.
3.2. Volatiles with Known Anti-Phytophthora Activity Are Emitted on Potato Leaves by Pseudomonas sp. S35
We then compared the emission of the 16 discriminative compounds mentioned above in the different samples (leaves non-inoculated or inoculated with the four Pseudomonas strains) using Tukey post hoc comparison (p-value < 0.05) (Figure 2).
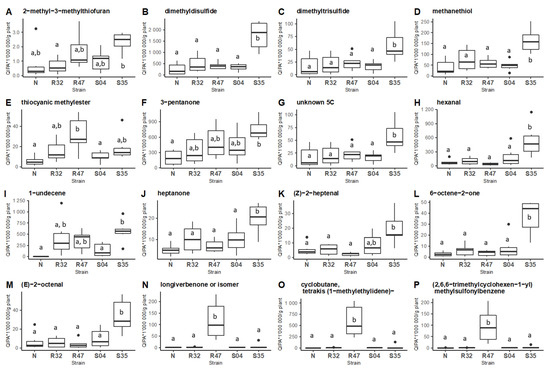
Figure 2.
Comparative emission of VOCs by different Pseudomonas inoculated on potato leaves. Boxplot of the 16 VOCs with a cosinus square superior to 0.6 according to the two first dimensions of the principal component analysis (A–P). Tukey HSD post-hoc tests with Holm correction were performed on each VOC between potato leaves non-inoculated (N) or inoculated with different Pseudomonas strains (R32, R47, S04, S35; N = 6; Quantitative Ion Peak Area/g plant (QIPA/g plant)). Different letters indicate statistically significant differences at a significance level of p < 0.05).
This analysis revealed 11 VOCs to be emitted in higher quantities in leaves inoculated with Pseudomonas sp. S35, among which most S-VOCs and 1-Undecene. It is worth noting that many of these compounds were previously shown to inhibit P. infestans in in vitro experiments. Inoculation of the leaves with P. chlororaphis R47 also led to higher emission of four VOCs, while the other Pseudomonas strains only marginally affected the volatile profiles of the leaves.
4. Discussion
The importance of volatile organic compounds (VOCs) for microbial communication and for the establishment of mutualistic interactions with other microbes, with plants and with animals is increasingly recognized [47,48]. Microbes, and bacteria in particular, are found in a wide range of environments differing in their physical and chemical properties, as well as in their nutrient availability. These factors are expected to exert a strong influence on the quality and quantity of the emitted VOCs. Accordingly, earlier studies have shown that similar strains can emit very different VOCs depending on their cultivation medium or growth phase [49,50,51,52]. The same laboratory studies have shown that even very nutrient-poor media are sufficient to support the emission of VOCs, triggering e.g., plant growth promotion [51] or inhibition of fungal phytopathogens [49]. Such inhibiting activity of bacterial volatiles on plant disease-causing agents is of great interest considering the need to find alternatives to the synthetic fungicides currently used to protect plants. However, the ability of plant-associated bacteria to emit such potent volatiles has been so-far only demonstrated in laboratory conditions, and the question remained whether they would also be emitted by bacteria growing on nutrient-poor leaf surfaces.
Using four Pseudomonas strains previously isolated either from the roots or from the shoots of potato plants, we have shown in this work that such volatile emission can indeed also occur on plant leaves. One particular volatile was detected only in leaves harboring Pseudomonas bacteria, namely 1-undecene, which is a well-known volatile emitted by members of this genus, and is even used as marker of infection by the widespread pathogen P. aeruginosa [53]. Plants, in contrast, are not known to produce this alkene, which indicates that the VOCs detected in our study are indeed produced by the bacteria and not by the plant as a reaction to bacterial inoculation. Likewise, the S-VOCs detected in our study such as dimethyl disulfide or dimethyl trisulfide are not known to be emitted by potato, although they are produced by other plants belonging to the Brassicaceae or the Alliaceae families [54]. In addition to being a marker of the genus Pseudomonas, 1-undecene was shown to strongly inhibit zoospore release in the late blight-causing oomycete Phytophthora infestans [9]. The fact that potato-associated Pseudomonas are able to emit this volatile at the site of infection is therefore of particular relevance for their putative use as biocontrol agents, although we cannot warrant at this stage that 1-undecene or the S-VOCs would be emitted in sufficient concentrations to support anti-Phytophthora activity. Indeed, this study only allowed comparative analysis of the emitted VOCs, but did not allow to determine the absolute concentrations found in the leaf headspaces, which is a goal of future research.
One surprising observation we made was the low number of VOCs differentially emitted by leaves infected vs. non-infected with P. infestans in our experimental conditions. This was probably due to a very mild infection that could have been caused by low fitness of the sporangia used to inoculate the leaves, or to the relatively short time span of the incubation, which did not allow the infection to fully develop. It might also be that infection with a hemibiotrophic pathogen does not induce the same extent of changes in VOCs emissions as those reported after attacks by herbivores or necrotrophic fungi [55].
Interestingly, the Pseudomonas strains used in this study differed in their VOCs profiles when growing on leaves, as illustrated in Figure 2. From the four strains studied, two stood out, with higher divergence from the non-inoculated control leaves: P. chlororaphis R47 and Pseudomonas sp. S35. These strains differ in many aspects, including the environment they were isolated from (potato rhizosphere for R47 and phyllosphere for S35) or their genome size and content (R47 harboring a wide variety of genes encoding antimicrobial compounds and S35 harboring several genes involved in plant colonization) [25]. While R47 showed very strong in vitro inhibition of P. infestans at various developmental stages, S35, which was largely inactive in vitro, provided the most consistent in planta protection against late blight in three different potato cultivars [25,56]. Although infection assays usually do not allow to separate VOCs-mediated effects from direct effects, our present study showing the emission of a range of anti-Phytophthora volatiles (e.g., 1-undecene, heptanone, dimethyl disulfide, and dimethyl trisulfide [22]) from the surface of S35-inoculated leaves highlights these VOCs as candidate compounds underlying at least partly the very good in planta protection conferred by this strain against P. infestans infection.
In conclusion, our study brings the proof of concept that bacterial VOCs can be emitted on leaf surfaces and not only on laboratory media. This further strengthens the translational potential of these bioactive metabolites and of their emitting strains, which could be used either as inducers of systemic resistance or as direct inhibitors of the pathogens’ growth and development. However, both the metabolites and the strains producing them would then need to withstand a yet harsher environment than that encountered on our in vitro potato leaves, facing e.g., UV light or drastic changes in temperature and humidity. Further studies shall therefore investigate the true potential of such in situ bacterial VOCs emission on plants growing under greenhouse and field conditions.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms10081510/s1, Figure S1: Principal component analysis of the volatiles emitted by leaves inoculated with Pseudomonas strains in presence and absence of P. infestans; Table S1: Abundance of bacteria on leaves 2 h and 120 h after inoculation. Table S2: List of volatile organic compounds identified in all leaf samples.
Author Contributions
Conceptualization, L.W., A.G., P.F. and M.D.V.; methodology, A.G., P.F. and M.D.V.; resources, L.W. and K.G.; data curation, P.F. and A.G.; writing—original draft preparation, L.W. and A.G.; writing—review and editing, P.F., A.G., K.G. and L.W.; supervision, L.W. and K.G.; project administration, L.W. and P.F.; funding acquisition, L.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Swiss National Science Foundation, grant number 179310 and the APC was also funded by the Swiss National Science Foundation.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors wish to thank Eric Droz and Heinz Krebs for providing in vitro potato plants and the P. infestans isolate, respectively. Financial support from the Swiss National Science Foundation (grant 179310 to L.W.) is gratefully acknowledged.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schlaeppi, K.; Bulgarelli, D. The Plant Microbiome at Work. Mol. Plant-Microbe Interact. 2015, 28, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–Microbiome Interactions: From Community Assembly to Plant Health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Müller, D.B.; Vogel, C.; Bai, Y.; Vorholt, J.A. The Plant Microbiota: Systems-Level Insights and Perspectives. Annu. Rev. Genet. 2016, 50, 211–234. [Google Scholar] [CrossRef] [PubMed]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef]
- Haas, D.; Défago, G. Biological Control of Soil-Borne Pathogens by Fluorescent Pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.M.; Bakker, P.A.H.M.H.M.; Pieterse, M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; et al. Induced Systemic Resistance by Beneficial Microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef]
- Weisskopf, L. The Potential of Bacterial Volatiles for Crop Protection against Phytophathogenic Fungi. In Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education; Formatex: Guadalajara, Mexico, 2013; pp. 1352–1363. [Google Scholar]
- Garbeva, P.; Weisskopf, L. Airborne Medicine—Bacterial Volatiles and Their Influence on Plant Health. New Phytol. 2020, 226, 32–43. [Google Scholar] [CrossRef]
- Hunziker, L.; Bönisch, D.; Groenhagen, U.; Bailly, A.; Schulz, S.; Weisskopf, L. Pseudomonas Strains Naturally Associated with Potato Plants Produce Volatiles with High Potential for Inhibition of Phytophthora Infestans. Appl. Environ. Microbiol. 2015, 81, 821–830. [Google Scholar] [CrossRef]
- Corre, M.; Mercier, A.; Bouteiller, M.; Khalil, A.; Ginevra, C.; Depayras, S.; Dupont, C. Bacterial Long-Range Warfare: Aerial Killing of Legionella Pneumophila by Pseudomonas Fl Uorescens. Microbiol. Spectr. 2021, 9, e00404-21. [Google Scholar] [CrossRef]
- Ossowicki, A.; Jafra, S.; Garbeva, P. The Antimicrobial Volatile Power of the Rhizospheric Isolate Pseudomonas Donghuensis P482. PLoS ONE 2017, 12, e0174362. [Google Scholar] [CrossRef]
- Fernando, W.G.D.; Ramarathnam, R.; Krishnamoorthy, A.S.; Savchuk, S.C. Identification and Use of Potential Bacterial Organic Antifungal Volatiles in Biocontrol. Soil Biol. Biochem. 2005, 37, 955–964. [Google Scholar] [CrossRef]
- Ryu, C.-M.; Farag, M.A.; Hu, C.-H.; Reddy, M.S.; Wei, H.-X.; Paré, P.W.; Kloepper, J.W. Bacterial Volatiles Promote Growth in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 4927–4932. [Google Scholar] [CrossRef]
- Ryu, C.-M.; Farag, M.A.; Hu, C.-H.; Reddy, M.S.; Kloepper, J.W.; Paré, P.W. Bacterial Volatiles Induce Systemic Resistance in Arabidopsis. Plant Physiol. 2004, 134, 1017–1026. [Google Scholar] [CrossRef]
- Baroja-Fernández, E.; Almagro, G.; Sánchez-López, Á.M.; Bahaji, A.; Gámez-Arcas, S.; De Diego, N.; Dolezal, K.; Muñoz, F.J.; Climent Sanz, E.; Pozueta-Romero, J. Enhanced Yield of Pepper Plants Promoted by Soil Application of Volatiles From Cell-Free Fungal Culture Filtrates Is Associated With Activation of the Beneficial Soil Microbiota. Front. Plant Sci. 2021, 12, 752653. [Google Scholar] [CrossRef]
- Chinchilla, D.; Bruisson, S.; Meyer, S.; Zühlke, D.; Hirschfeld, C.; Joller, C.; L’Haridon, F.; Mène-Saffrané, L.; Riedel, K.; Weisskopf, L. A Sulfur-Containing Volatile Emitted by Potato-Associated Bacteria Confers Protection against Late Blight through Direct Anti-Oomycete Activity. Sci. Rep. 2019, 9, 18778. [Google Scholar] [CrossRef]
- Li, Q.; Ning, P.; Zheng, L.; Huang, J.; Li, G.; Hsiang, T. Effects of Volatile Substances of Streptomyces Globisporus JK-1 on Control of Botrytis Cinerea on Tomato Fruit. Biol. Control 2012, 61, 113–120. [Google Scholar] [CrossRef]
- Lazazzara, V.; Vicelli, B.; Bueschl, C.; Parich, A.; Pertot, I.; Schuhmacher, R.; Perazzolli, M. Trichoderma Spp. Volatile Organic Compounds Protect Grapevine Plants by Activating Defense-Related Processes against Downy Mildew. Physiol. Plant. 2021, 172, 1950–1965. [Google Scholar] [CrossRef]
- Kamoun, S.; Furzer, O.; Jones, J.D.G.; Judelson, H.S.; Ali, G.S.; Dalio, R.J.D.; Roy, S.G.; Schena, L.; Zambounis, A.; Panabières, F.; et al. The Top 10 Oomycete Pathogens in Molecular Plant Pathology. Mol. Plant Pathol. 2015, 16, 413–434. [Google Scholar] [CrossRef]
- Fry, W.E.; Birch, P.R.J.; Judelson, H.S.; Grünwald, N.J.; Danies, G.; Everts, K.L.; Gevens, A.J.; Gugino, B.K.; Johnson, D.A.; Johnson, S.B.; et al. Five Reasons to Consider Phytophthora Infestans a Reemerging Pathogen. Phytopathology 2015, 105, 966–981. [Google Scholar] [CrossRef]
- Goodwin, S.B.; Smart, C.D.; Sandrock, R.W.; Deahl, K.L.; Punja, Z.K.; Fry, W.E. Genetic Change Within Populations of Phytophthora Infestans in the United States and Canada During 1994 to 1996: Role of Migration and Recombination. Phytopathology 1998, 88, 939–949. [Google Scholar] [CrossRef]
- De Vrieze, M.; Pandey, P.; Bucheli, T.D.; Varadarajan, A.R.; Ahrens, C.H.; Weisskopf, L.; Bailly, A. Volatile Organic Compounds from Native Potato-Associated Pseudomonas as Potential Anti-Oomycete Agents. Front. Microbiol. 2015, 6, 1295. [Google Scholar] [CrossRef]
- Meldau, D.G.; Meldau, S.; Hoang, L.H.; Underberg, S.; Wünsche, H.; Baldwin, I.T.; Wunsche, H.; Baldwin, I.T. Dimethyl Disulfide Produced by the Naturally Associated Bacterium Bacillus Sp B55 Promotes Nicotiana Attenuata Growth by Enhancing Sulfur Nutrition. Plant Cell 2013, 25, 2731–2747. [Google Scholar] [CrossRef]
- Huang, C.J.; Tsay, J.F.; Chang, S.Y.; Yang, H.P.; Wu, W.S.; Chen, C.Y. Dimethyl Disulfide Is an Induced Systemic Resistance Elicitor Produced by Bacillus Cereus C1L. Pest Manag. Sci. 2012, 68, 1306–1310. [Google Scholar] [CrossRef]
- De Vrieze, M.; Varadarajan, A.R.; Schneeberger, K.; Bailly, A.; Ahrens, C.H.; Weisskopf, L. Linking Comparative Genomics of Nine Potato-Associated Pseudomonas Isolates with Their Differing Biocontrol Potential against Late Blight. Front. Microbiol. 2020, 11, 857. [Google Scholar] [CrossRef]
- Clark, D.J.; Maaløe, O. DNA Replication and the Division Cycle in Escherichia Coli. J. Mol. Biol. 1967, 23, 99–112. [Google Scholar] [CrossRef]
- De Vrieze, M.; Gloor, R.; Massana Codina, J.; Torriani, S.; Gindro, K.; L’Haridon, F.; Bailly, A.; Weisskopf, L. Biocontrol Activity of Three Pseudomonas on a Newly Assembled Collection of Phytophthora Infestans Isolates. Phytopathology 2019, 109, 1555–1565. [Google Scholar] [CrossRef]
- Jaime-Garcia, R.; Trinidad-Correa, R.; Felix-Gastelum, R.; Orum, T.V.; Wasmann, C.C.; Nelson, M.R. Temporal and Spatial Patterns of Genetic Structure of Phytophthora Infestans from Tomato and Potato in the Del Fuerte Valley. Phytopathology 2000, 90, 1188–1195. [Google Scholar] [CrossRef][Green Version]
- Fuchsmann, P.; Stern, M.T.; Brügger, Y.A.; Breme, K. Olfactometry Profiles and Quantitation of Volatile Sulfur Compounds of Swiss Tilsit Cheeses. J. Agric. Food Chem. 2015, 63, 7511–7521. [Google Scholar] [CrossRef]
- Carrapiso, A.I.; Jurado, Á.; Timón, M.L.; García, C. Odor-Active Compounds of Iberian Hams with Different Aroma Characteristics. J. Agric. Food Chem. 2002, 50, 6453–6458. [Google Scholar] [CrossRef]
- Varming, C.; Petersen, M.A.; Poll, L. Comparison of Isolation Methods for the Determination of Important Aroma Compounds in Black Currant (Ribes nigrum L.) Juice, Using Nasal Impact Frequency Profiling. J. Agric. Food Chem. 2004, 52, 1647–1652. [Google Scholar] [CrossRef]
- Ott, A.; Fay, L.B.; Chaintreau, A. Determination and Origin of the Aroma Impact Compounds of Yogurt Flavor. J. Agric. Food Chem. 1997, 45, 850–858. [Google Scholar] [CrossRef]
- Tatsuka, K.; Suekane, S.; Sakai, Y.; Sumitani, H. Volatile Constituents of Kiwi Fruit Flowers: Simultaneous Distillation and Extraction versus Headspace Sampling. J. Agric. Food Chem. 1990, 38, 2176–2180. [Google Scholar] [CrossRef]
- Guillard, A.-S.; Le Quere, J.-L.; Vendeuvre, J.-L. Emerging Research Approaches Benefit to the Study of Cooked Cured Ham Flavour. Food Chem. 1997, 59, 567–572. [Google Scholar] [CrossRef]
- Piyachaiseth, T.; Jirapakkul, W.; Chaiseri, S. Aroma Compounds of Flash-Fried Rice. Kasetsart J.-Nat. Sci. 2011, 45, 717–729. [Google Scholar]
- Alasalvar, C.; Shahidi, F.; Cadwallader, K.R. Comparison of Natural and Roasted Turkish Tombul Hazelnut (Corylus avellana L.) Volatiles and Flavor by DHA/GC/MS and Descriptive Sensory Analysis. J. Agric. Food Chem. 2003, 51, 5067–5072. [Google Scholar] [CrossRef]
- Boti, J.B.; Koukoua, G.; N’Guessan, T.Y.; Muselli, A.; Bernardini, A.-F.; Casanova, J. Composition of the Leaf, Stem Bark and Root Bark Oils of Isolona Cooperi Investigated by GC (Retention Index), GC-MS and 13C-NMR Spectroscopy. Phytochem. Anal. 2005, 16, 357–363. [Google Scholar] [CrossRef]
- Cho, I.H.; Namgung, H.J.; Choi, H.K.; Kim, Y.S. Volatiles and Key Odorants in the Pileus and Stipe of Pine-Mushroom (Tricholoma Matsutake Sing.). Food Chem. 2008, 106, 71–76. [Google Scholar] [CrossRef]
- Hirth, M.; Leiter, A.; Beck, S.M.; Schuchmann, H.P. Effect of Extrusion Cooking Process Parameters on the Retention of Bilberry Anthocyanins in Starch Based Food. J. Food Eng. 2014, 125, 139–146. [Google Scholar] [CrossRef]
- Jarunrattanasri, A.; Theerakulkait, C.; Cadwallader, K.R. Aroma Components of Acid-Hydrolyzed Vegetable Protein Made by Partial Hydrolysis of Rice Bran Protein. J. Agric. Food Chem. 2007, 55, 3044–3050. [Google Scholar] [CrossRef]
- Mockute, D.; Bernotiene, G. The Main Citral−Geraniol and Carvacrol Chemotypes of the Essential Oil of Thymus pulegioides L. Growing Wild in Vilnius District (Lithuania). J. Agric. Food Chem. 1999, 47, 3787–3790. [Google Scholar] [CrossRef]
- Radulović, N.; Blagojević, P.; Palić, R. Comparative Study of the Leaf Volatiles of Arctostaphylos Uva-ursi (L.) Spreng. and Vaccinium Vitis-idaea L. (Ericaceae). Molecules 2010, 15, 6168–6185. [Google Scholar] [CrossRef] [PubMed]
- Deasy, W.; Shepherd, T.; Alexander, C.J.; Birch, A.N.E.; Evans, K.A. Development and Validation of a SPME-GC-MS Method for In Situ Passive Sampling of Root Volatiles from Glasshouse-Grown Broccoli Plants Undergoing Below-Ground Herbivory by Larvae of Cabbage Root Fly, Delia radicum L. Phytochem. Anal. 2016, 27, 375–393. [Google Scholar] [CrossRef] [PubMed]
- Rychlik, M.; Schieberle, P.; Grosch, W. Compilation of Odor Thresholds, Odor Qualities and Retention Indices of Key Food Odorants; Technical University of Munich Garching: München, Germany, 1998. [Google Scholar]
- Wei, S.; Marton, I.; Dekel, M.; Shalitin, D.; Lewinsohn, E.; Bravdo, B.-A.; Shoseyov, O. Manipulating Volatile Emission in Tobacco Leaves by Expressing Aspergillus Nigerβ-Glucosidase in Different Subcellular Compartments. Plant Biotechnol. J. 2004, 2, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Wakamura, S.; Hattori, M.; Igita, K.; Yasuda, K. Tridjaka Sex Pheromone of Etiella Behrii, a Pod Borer of Soybean in Indonesia: Identification and Field Attraction. Entomol. Exp. Appl. 1999, 91, 413–420. [Google Scholar] [CrossRef]
- Weisskopf, L.; Schulz, S.; Garbeva, P. Microbial Volatile Organic Compounds in Intra-Kingdom and Inter-Kingdom Interactions. Nat. Rev. Microbiol. 2021, 19, 391–404. [Google Scholar] [CrossRef]
- Netzker, T.; Shepherdson, E.M.F.; Zambri, M.P.; Elliot, M.A. Bacterial Volatile Compounds: Functions in Communication, Cooperation, and Competition. Annu. Rev. Microbiol. 2020, 74, 409–430. [Google Scholar] [CrossRef]
- Garbeva, P.; Hordijk, C.; Gerards, S.; de Boer, W. Volatiles Produced by the Mycophagous Soil Bacterium Collimonas. FEMS Microbiol. Ecol. 2014, 87, 639–649. [Google Scholar] [CrossRef]
- Kai, M.; Crespo, E.; Cristescu, S.M.; Harren, F.J.M.; Francke, W.; Piechulla, B. Serratia Odorifera: Analysis of Volatile Emission and Biological Impact of Volatile Compounds on Arabidopsis Thaliana. Appl. Microbiol. Biotechnol. 2010, 88, 965–976. [Google Scholar] [CrossRef]
- Blom, D.; Fabbri, C.; Connor, E.C.; Schiestl, F.P.; Klauser, D.R.; Boller, T.; Eberl, L.; Weisskopf, L. Production of Plant Growth Modulating Volatiles Is Widespread among Rhizosphere Bacteria and Strongly Depends on Culture Conditions. Environ. Microbiol. 2011, 13, 3047–3058. [Google Scholar] [CrossRef]
- Becher, P.G.; Verschut, V.; Bibb, M.J.; Bush, M.J.; Molnár, B.P.; Barane, E.; Al-Bassam, M.M.; Chandra, G.; Song, L.; Challis, G.L.; et al. Developmentally Regulated Volatiles Geosmin and 2-Methylisoborneol Attract a Soil Arthropod to Streptomyces Bacteria Promoting Spore Dispersal. Nat. Microbiol. 2020, 5, 821–829. [Google Scholar] [CrossRef]
- Rui, Z.; Li, X.; Zhu, X.; Liu, J.; Domigan, B.; Barr, I.; Cate, J.H.D.; Zhang, W. Microbial Biosynthesis of Medium-Chain 1-Alkenes by a Nonheme Iron Oxidase. Proc. Natl. Acad. Sci. USA 2014, 111, 10–15. [Google Scholar] [CrossRef]
- Kremr, D.; Bajerová, P.; Bajer, T.; Eisner, A.; Adam, M.; Ventura, K. Using Headspace Solid-Phase Microextraction for Comparison of Volatile Sulphur Compounds of Fresh Plants Belonging to Families Alliaceae and Brassicaceae. J. Food Sci. Technol. 2015, 52, 5727–5735. [Google Scholar] [CrossRef]
- Ameye, M.; Allmann, S.; Verwaeren, J.; Smagghe, G.; Haesaert, G.; Schuurink, R.C.; Audenaert, K. Green Leaf Volatile Production by Plants: A Meta-Analysis. New Phytol. 2018, 220, 666–683. [Google Scholar] [CrossRef]
- De Vrieze, M.; Germanier, F.; Vuille, N.; Weisskopf, L. Combining Different Potato-Associated Pseudomonas Strains for Improved Biocontrol of Phytophthora Infestans. Front. Microbiol. 2018, 9, 2573. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).