
Quorum Sensing and Quorum Quenching with a Focus on Cariogenic and Periodontopathic Oral Biofilms


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Bacterial QS Model Systems
2.1. Intraspecies QS in Gram-Negative Bacteria
2.2. Intraspecies QS in Gram-Positive Bacteria
2.3. Interspecies AI-2 Signalling Model
3. The Effect of Fluid Flow in QS
4. QS in Cariology
4.1. QS and QQ in S. mutans
4.1.1. CSP/ComABCDE QS Pathway in S. mutans
4.1.2. XIP-ComRS QS Pathway in S. mutans
4.1.3. PdrA-WGK QS Pathway in S. mutans

4.1.4. QQ in S. mutans by Plant Products
4.1.5. QQ in S. mutans by Microbial Products
4.1.6. QQ in S. mutans by Synthetic Molecules
4.2. QS and QQ in Candida albicans and Dual Species S. mutans-C. albicans Biofilms
5. QS and QQ in Periodontitis
5.1. QS Interactions among Periodontal Pathogens
5.2. QQ Compounds in Periodontitis
5.2.1. Furanone Compounds and Combinations with D-Ribose
5.2.2. D-Galactose
5.2.3. Natural Products
5.2.4. Synthetic Analogues of AHL
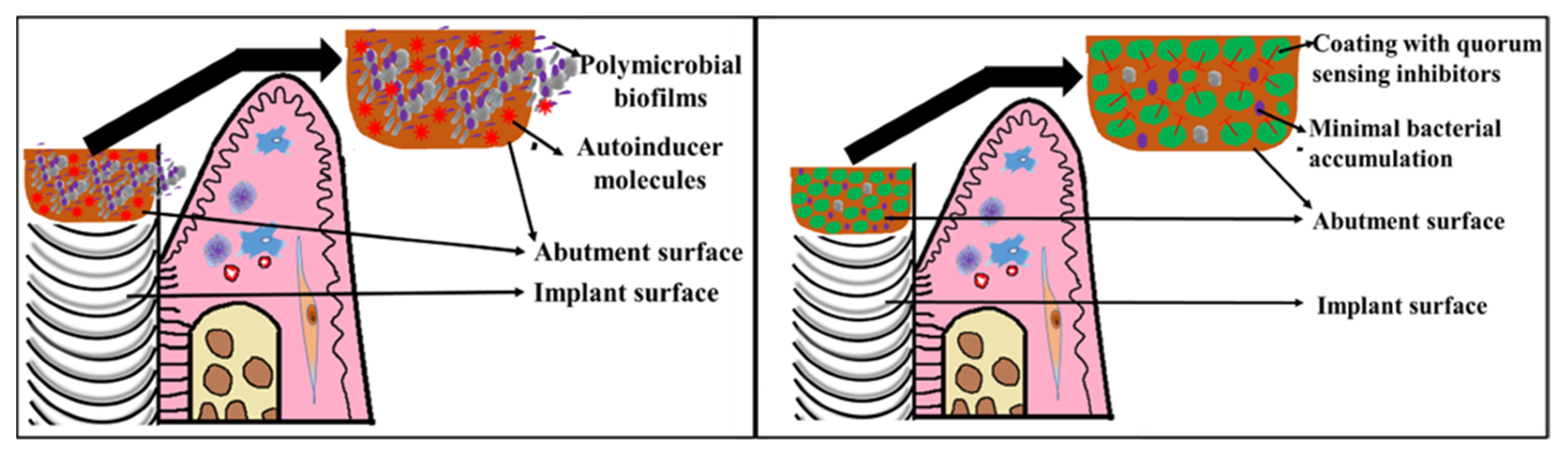
6. Coating Surfaces of Dental Implants with QS Inhibitors
7. Conclusions, Limitations, and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Grandclément, C.; Tannières, M.; Moréra, S.; Dessaux, Y.; Faure, D. Quorum quenching: Role in nature and applied developments. FEMS Microbiol. Rev. 2016, 40, 86–116. [Google Scholar] [CrossRef] [PubMed]
- Bzdrenga, J.; Daudé, D.; Rémy, B.; Jacquet, P.; Plener, L.; Elias, M.; Chabrière, E. Biotechnological applications of quorum quenching enzymes. Chem. Biol. Interact. 2017, 267, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, S.T.; Bassler, B.L. Bacterial quorum sensing: Its role in virulence and possibilities for its control. Cold Spring Harb. Perspect. Med. 2012, 2, a012427. [Google Scholar] [CrossRef] [PubMed]
- Whiteley, M.; Diggle, S.P.; Greenberg, E.P. Bacterial quorum sensing: The progress and promise of an emerging research area. Nature 2017, 551, 313. [Google Scholar] [CrossRef] [PubMed]
- Nealson, K.H.; Platt, T.; Hastings, J.W. Cellular control of the synthesis and activity of the bacterial luminescent system. J. Bacteriol. 1970, 104, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef]
- Jayathilake, P.G.; Jana, S.; Rushton, S.; Swailes, D.; Bridgens, B.; Curtis, T.; Chen, J. Extracellular polymeric substance production and aggregated bacteria colonization influence the competition of microbes in biofilms. Front. Microbiol. 2017, 8, 1865. [Google Scholar] [CrossRef]
- Frederick, M.R.; Kuttler, C.; Hense, B.A.; Eberl, H.J. A mathematical model of quorum sensing regulated EPS production in biofilm communities. Theor. Biol. Med. Model. 2011, 8, 8. [Google Scholar] [CrossRef]
- Tan, C.H.; Koh, K.S.; Xie, C.; Tay, M.; Zhou, Y.; Williams, R.; Ng, W.J.; Rice, S.A.; Kjelleberg, S. The role of quorum sensing signalling in EPS production and the assembly of a sludge community into aerobic granules. ISME J. 2014, 8, 1186–1197. [Google Scholar] [CrossRef]
- Smith, E.G.; Spatafora, G.A. Gene regulation in S. mutans: Complex control in a complex environment. J. Dent. Res. 2012, 91, 133–141. [Google Scholar] [CrossRef]
- Balcázar, J.L.; Subirats, J.; Borrego, C.M. The role of biofilms as environmental reservoirs of antibiotic resistance. Front. Microbiol. 2015, 6, 1216. [Google Scholar] [CrossRef] [PubMed]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Shelenkov, A. Whole-genome sequencing of pathogenic bacteria-new insights into antibiotic resistance spreading. Microorganisms 2021, 9, 2624. [Google Scholar] [CrossRef] [PubMed]
- Aslam, B.; Khurshid, M.; Arshad, M.I.; Muzammil, S.; Rasool, M.; Yasmeen, N.; Shah, T.; Chaudhry, T.H.; Rasool, M.H.; Shahid, A.; et al. Antibiotic resistance: One health one world outlook. Front. Cell Infect. Microbiol. 2021, 11, 771510. [Google Scholar] [CrossRef]
- Collaborators. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Piddock, L.J. The crisis of no new antibiotics—What is the way forward? Lancet Infect. Dis. 2012, 12, 249–253. [Google Scholar] [CrossRef]
- Albernaz-Gonçalves, R.; Olmos, G.; Hötzel, M.J. Exploring farmers’ reasons for antibiotic use and misuse in pig farms in brazil. Antibiotics 2021, 10, 331. [Google Scholar] [CrossRef]
- Gozdzielewska, L.; King, C.; Flowers, P.; Mellor, D.; Dunlop, P.; Price, L. Scoping review of approaches for improving antimicrobial stewardship in livestock farmers and veterinarians. Prev. Vet. Med. 2020, 180, 105025. [Google Scholar] [CrossRef]
- Roy, V.; Adams, B.L.; Bentley, W.E. Developing next generation antimicrobials by intercepting AI-2 mediated quorum sensing. Enzym. Microb. Technol. 2011, 49, 113–123. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Lamont, R.J. Polymicrobial communities in periodontal disease: Their quasi-organismal nature and dialogue with the host. Periodontology 2000 2021, 86, 210–230. [Google Scholar] [CrossRef]
- Iebba, V.; Totino, V.; Gagliardi, A.; Santangelo, F.; Cacciotti, F.; Trancassini, M.; Mancini, C.; Cicerone, C.; Corazziari, E.; Pantanella, F.; et al. Eubiosis and dysbiosis: The two sides of the microbiota. New Microbiol. 2016, 39, 1–12. [Google Scholar] [PubMed]
- Magda Feres, H.D. Systemic Antibiotics in Periodontal Therapy. In Lindhe’s Clinical Periodontology and Implant Dentistry; Niklaus, W.V.G., Lang, P., Mariano Sanz, T.B., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2021. [Google Scholar]
- Ali, I.A.A.; Neelakantan, P. Antibiofilm activity of phytochemicals against Enterococcus faecalis: A literature review. Phytother. Res. 2022, 36, 2824–2838. [Google Scholar] [CrossRef] [PubMed]
- Hasan, S.; Danishuddin, M.; Adil, M.; Singh, K.; Verma, P.K.; Khan, A.U. Efficacy of E. officinalis on the cariogenic properties of Streptococcus mutans: A novel and alternative approach to suppress quorum-sensing mechanism. PLoS ONE 2012, 7, e40319. [Google Scholar]
- Li, X.; Yin, L.; Ramage, G.; Li, B.; Tao, Y.; Zhi, Q.; Lin, H.; Zhou, Y. Assessing the impact of curcumin on dual-species biofilms formed by Streptococcus mutans and Candida albicans. Microbiologyopen 2019, 8, e937. [Google Scholar] [CrossRef]
- Vijayakumar, A.; Sarveswari, H.B.; Vasudevan, S.; Shanmugam, K.; Solomon, A.P.; Neelakantan, P. Baicalein inhibits Streptococcus mutans biofilms and dental caries-related virulence phenotypes. Antibiotics 2021, 10, 215. [Google Scholar] [CrossRef]
- Listl, S.; Galloway, J.; Mossey, P.A.; Marcenes, W. Global economic impact of dental diseases. J. Dent. Res. 2015, 94, 1355–1361. [Google Scholar] [CrossRef]
- Kassebaum, N.; Bernabé, E.; Dahiya, M.; Bhandari, B.; Murray, C.; Marcenes, W. Global burden of untreated caries: A systematic review and metaregression. J. Dent. Res. 2015, 94, 650–658. [Google Scholar] [CrossRef]
- Das, M.; Upadhyaya, V.; Ramachandra, S.S.; Jithendra, K.D. Periodontal treatment needs in diabetic and non-diabetic individuals: A case-control study. Indian J. Dent. Res. 2011, 22, 291–294. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, S.Q.; Zhao, L.; Ren, Z.H.; Hu, C.Y. Global, regional and national burden of periodontitis from 1990 to 2019: Results from the Global Burden of Disease study 2019. J. Periodontol. 2022. Online ahead of print. Available online: https://aap.onlinelibrary.wiley.com/doi/10.1002/JPER.21-0469 (accessed on 31 August 2022).
- Mendhi, J.; Ramachandra, S.S.; Prasadam, I.; Ivanovski, S.; Yang, Y.; Xiao, Y. Endogenous nitric oxide-generating surfaces via polydopamine-copper coatings for preventing biofilm dispersal and promoting microbial killing. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 128, 112297. [Google Scholar] [CrossRef]
- Xiang, E.; Gómez-Cerezo, M.N.; Ali, Y.; Ramachandra, S.S.; Yang, N.; Dargusch, M.; Moran, C.S.; Ivanovski, S.; Abdal-Hay, A. Surface Modification of pure zinc by acid etching: Accelerating the corrosion rate and enhancing biocompatibility and antibacterial characteristics. ACS Appl. Mater. Interfaces 2022, 14, 22554–22569. [Google Scholar] [CrossRef]
- Kim, W.J.; Soh, Y.; Heo, S.-M. Recent advances of therapeutic targets for the treatment of periodontal disease. Biomol. Ther. 2021, 29, 263. [Google Scholar] [CrossRef] [PubMed]
- Werlang, C.A.; Chen, W.G.; Aoki, K.; Wheeler, K.M.; Tymm, C.; Mileti, C.J.; Burgos, A.C.; Kim, K.; Tiemeyer, M.; Ribbeck, K. Mucin O-glycans suppress quorum-sensing pathways and genetic transformation in Streptococcus mutans. Nat. Microbiol. 2021, 6, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Duddy, O.P.; Bassler, B.L. Quorum sensing across bacterial and viral domains. PLoS Pathog. 2021, 17, e1009074. [Google Scholar] [CrossRef] [PubMed]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P. Quorum sensing in bacteria: The LuxR-LuxI family of cell density-responsive transcriptional regulators. J. Bacteriol. 1994, 176, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, A.; Burlingame, A.L.; Eberhard, C.; Kenyon, G.L.; Nealson, K.H.; Oppenheimer, N. Structural identification of autoinducer of Photobacterium fischeri luciferase. Biochemistry 1981, 20, 2444–2449. [Google Scholar] [CrossRef]
- Engebrecht, J.; Nealson, K.; Silverman, M. Bacterial bioluminescence: Isolation and genetic analysis of functions from Vibrio fischeri. Cell 1983, 32, 773–781. [Google Scholar] [CrossRef]
- Engebrecht, J.; Silverman, M. Identification of genes and gene products necessary for bacterial bioluminescence. Proc. Natl. Acad. Sci. USA 1984, 81, 4154–4158. [Google Scholar] [CrossRef]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal-response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef]
- Case, R.J.; Labbate, M.; Kjelleberg, S. AHL-driven quorum-sensing circuits: Their frequency and function among the Proteobacteria. ISME J. 2008, 2, 345–349. [Google Scholar] [CrossRef]
- Bassler, B.L.; Greenberg, E.P.; Stevens, A.M. Cross-species induction of luminescence in the quorum-sensing bacterium Vibrio harveyi. J. Bacteriol. 1997, 179, 4043–4045. [Google Scholar] [CrossRef]
- Schauder, S.; Shokat, K.; Surette, M.G.; Bassler, B.L. The LuxS family of bacterial autoinducers: Biosynthesis of a novel quorum-sensing signal molecule. Mol. Microbiol. 2001, 41, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Ben Amara, H.; Song, H.Y.; Ryu, E.; Park, J.S.; Schwarz, F.; Kim, B.M.; Choi, B.K.; Koo, K.T. Effects of quorum-sensing inhibition on experimental periodontitis induced by mixed infection in mice. Eur. J. Oral Sci. 2018, 126, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Schauder, S.; Potier, N.; Van Dorsselaer, A.; Pelczer, I.; Bassler, B.L.; Hughson, F.M. Structural identification of a bacterial quorum-sensing signal containing boron. Nature 2002, 415, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.T.; Xavier, K.B.; Campagna, S.R.; Taga, M.E.; Semmelhack, M.F.; Bassler, B.L.; Hughson, F.M. Salmonella typhimurium recognizes a chemically distinct form of the bacterial quorum-sensing signal AI-2. Mol. Cell 2004, 15, 677–687. [Google Scholar] [CrossRef]
- Zhang, L.; Li, S.; Liu, X.; Wang, Z.; Jiang, M.; Wang, R.; Xie, L.; Liu, Q.; Xie, X.; Shang, D.; et al. Sensing of autoinducer-2 by functionally distinct receptors in prokaryotes. Nat. Commun. 2020, 11, 5371. [Google Scholar] [CrossRef]
- Dalwadi, M.P. Pearce, P. Emergent robustness of bacterial quorum sensing in fluid flow. Proc. Natl. Acad. Sci. USA 2021, 118, e2022312118. [Google Scholar] [CrossRef]
- Azeredo, J.; Azevedo, N.F.; Briandet, R.; Cerca, N.; Coenye, T.; Costa, A.R.; Desvaux, M.; Di Bonaventura, G.; Hebraud, M.; Jaglic, Z.; et al. Critical review on biofilm methods. Crit. Rev. Microbiol. 2017, 43, 313–351. [Google Scholar] [CrossRef]
- Kirisits, M.J.; Margolis, J.J.; Purevdorj-Gage, B.L.; Vaughan, B.; Chopp, D.L.; Stoodley, P.; Parsek, M.R. Influence of the hydrodynamic environment on quorum sensing in Pseudomonas aeruginosa biofilms. J. Bacteriol. 2007, 189, 8357–8360. [Google Scholar] [CrossRef]
- Kim, M.K.; Ingremeau, F.; Zhao, A.; Bassler, B.L.; Stone, H.A. Local and global consequences of flow on bacterial quorum sensing. Nat. Microbiol. 2016, 1, 15005. [Google Scholar] [CrossRef]
- Nicolae, V.; Neamtu, B.; Picu, O.; Stefanache, M.A.M.; Cioranu, V.S.I. The comparative evaluation of salivary biomarkers (Calcium, Phosphate, Salivary pH) in cario-resistance versus cario-activity. Rev. Chim. 2016, 67, 821–824. [Google Scholar]
- Gross, E.L.; Beall, C.J.; Kutsch, S.R.; Firestone, N.D.; Leys, E.J.; Griffen, A.L. Beyond Streptococcus mutans: Dental caries onset linked to multiple species by 16S rRNA community analysis. PLoS ONE 2012, 7, e47722. [Google Scholar] [CrossRef] [PubMed]
- Johansson, I.; Witkowska, E.; Kaveh, B.; Lif Holgerson, P.; Tanner, A.C. The microbiome in populations with a low and high prevalence of caries. J. Dent. Res. 2016, 95, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Huang, X.; Alkhers, N.; Alzamil, H.; Alzoubi, S.; Wu, T.T.; Castillo, D.A.; Campbell, F.; Davis, J.; Herzog, K.; et al. Candida albicans and early childhood caries: A systematic review and meta-analysis. Caries Res. 2018, 52, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Bikash, C.R. Tal-Gan, Y. Structure activity relationship study of the XIP quorum sensing pheromone in Streptococcus mutans reveal inhibitors of the competence regulon. ACS Chem. Biol. 2020, 15, 2833–2841. [Google Scholar] [CrossRef]
- OmerOglou, E.; Karaca, B.; Kibar, H.; Haliscelik, O.; Kiran, F. The role of microbiota-derived postbiotic mediators on biofilm formation and quorum sensing-mediated virulence of Streptococcus mutans: A perspective on preventing dental caries. Microb. Pathog. 2022, 164, 105390. [Google Scholar] [CrossRef]
- Kaspar, J.R.; Walker, A.R. Expanding the vocabulary of peptide signals in Streptococcus mutans. Front. Cell Infect. Microbiol. 2019, 9, 194. [Google Scholar] [CrossRef]
- Suzuki, Y.; Nagasawa, R.; Senpuku, H. Inhibiting effects of fructanase on competence-stimulating peptide-dependent quorum sensing system in Streptococcus mutans. J. Infect. Chemother. 2017, 23, 634–641. [Google Scholar] [CrossRef]
- Okinaga, T.; Niu, G.; Xie, Z.; Qi, F.; Merritt, J. The hdrRM operon of Streptococcus mutans encodes a novel regulatory system for coordinated competence development and bacteriocin production. J. Bacteriol. 2010, 192, 1844–1852. [Google Scholar] [CrossRef]
- Underhill, S.A.M.; Shields, R.C.; Kaspar, J.R.; Haider, M.; Burne, R.A.; Hagen, S.J. Intracellular signaling by the comRS system in Streptococcus mutans genetic competence. Msphere 2018, 3, e00444-18. [Google Scholar] [CrossRef]
- Xie, Z.; Okinaga, T.; Niu, G.; Qi, F.; Merritt, J. Identification of a novel bacteriocin regulatory system in Streptococcus mutans. Mol. Microbiol. 2010, 78, 1431–1447. [Google Scholar] [CrossRef]
- Shanker, E.; Federle, M.J. Quorum sensing regulation of competence and bacteriocins in Streptococcus pneumoniae and mutans. Genes 2017, 8, 15. [Google Scholar] [CrossRef] [PubMed]
- Rued, B.E.; Covington, B.C.; Bushin, L.B.; Szewczyk, G.; Laczkovich, I.; Seyedsayamdost, M.R.; Federle, M.J. Quorum sensing in streptococcus mutans regulates production of tryglysin, a novel ras-ripp antimicrobial compound. MBio 2021, 12, e02688-20. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, X.; Ling, J. Streptococcus gordonii LuxS/autoinducer-2 quorum-sensing system modulates the dual-species biofilm formation with Streptococcus mutans. J. Basic Microbiol. 2017, 57, 605–616. [Google Scholar] [CrossRef]
- Yuan, K.; Hou, L.; Jin, Q.; Niu, C.; Mao, M.; Wang, R.; Huang, Z. Comparative transcriptomics analysis of Streptococcus mutans with disruption of LuxS/AI-2 quorum sensing and recovery of methyl cycle. Arch. Oral Biol. 2021, 127, 105137. [Google Scholar] [CrossRef] [PubMed]
- Kaspar, J.R.; Lee, K.; Richard, B.; Walker, A.R.; Burne, R.A. Direct interactions with commensal streptococci modify intercellular communication behaviors of Streptococcus mutans. ISME J. 2021, 15, 473–488. [Google Scholar] [CrossRef]
- Muadcheingka, T.; Tantivitayakul, P. Distribution of Candida albicans and non-albicans Candida species in oral candidiasis patients: Correlation between cell surface hydrophobicity and biofilm forming activities. Arch. Oral Biol. 2015, 60, 894–901. [Google Scholar] [CrossRef]
- Pereira, D.F.A.; Seneviratne, C.J.; Koga-Ito, C.Y.; Samaranayake, L.P. Is the oral fungal pathogen Candida albicans a cariogen? Oral Dis. 2018, 24, 518–526. [Google Scholar] [CrossRef]
- Lozano Moraga, C.P.; Rodríguez Martínez, G.A.; Lefimil Puente, C.A.; Morales Bozo, I.C.; Urzúa Orellana, B.R. Prevalence of Candida albicans and carriage of Candida non-albicans in the saliva of preschool children, according to their caries status. Acta Odontol. Scand. 2017, 75, 30–35. [Google Scholar] [CrossRef]
- Hornby, J.M.; Jensen, E.C.; Lisec, A.D.; Tasto, J.J.; Jahnke, B.; Shoemaker, R.; Dussault, P.; Nickerson, K.W. Quorum sensing in the dimorphic fungus Candida albicans is mediated by farnesol. Appl. Environ. Microbiol. 2001, 67, 2982–2992. [Google Scholar] [CrossRef]
- Petersen, F.C.; Scheie, A.A. Genetic transformation in Streptococcus mutans requires a peptide secretion-like apparatus. Oral Microbiol. Immunol. 2000, 15, 329–334. [Google Scholar] [CrossRef]
- Li, Y.H.; Lau, P.C.; Lee, J.H.; Ellen, R.P.; Cvitkovitch, D.G. Natural genetic transformation of Streptococcus mutans growing in biofilms. J. Bacteriol. 2001, 183, 897–908. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Cao, L.; Kim, A.; Biswas, I. SepM, a streptococcal protease involved in quorum sensing, displays strict substrate specificity. J. Bacteriol. 2016, 198, 436–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.H.; Tang, N.; Aspiras, M.B.; Lau, P.C.; Lee, J.H.; Ellen, R.P.; Cvitkovitch, D.G. A quorum-sensing signaling system essential for genetic competence in Streptococcus mutans is involved in biofilm formation. J. Bacteriol. 2002, 184, 2699–2708. [Google Scholar] [CrossRef] [PubMed]
- Ishii, S.; Fukui, K.; Yokoshima, S.; Kumagai, K.; Beniyama, Y.; Kodama, T.; Fukuyama, T.; Okabe, T.; Nagano, T.; Kojima, H.; et al. High.-throughput screening of small molecule inhibitors of the Streptococcus quorum-sensing signal pathway. Sci. Rep. 2017, 7, 4029. [Google Scholar] [CrossRef]
- Kaspar, J.; Underhill, S.A.; Shields, R.C.; Reyes, A.; Rosenzweig, S.; Hagen, S.J.; Burne, R.A. Intercellular communication via the comX-inducing peptide (XIP) of Streptococcus mutans. J. Bacteriol. 2017, 199, e00404-17. [Google Scholar] [CrossRef]
- Mashburn-Warren, L.; Morrison, D.A.; Federle, M.J. A novel double-tryptophan peptide pheromone controls competence in Streptococcus spp. via an Rgg regulator. Mol. Microbiol. 2010, 78, 589–606. [Google Scholar] [CrossRef]
- Talagas, A.; Fontaine, L.; Ledesma-García, L.; Mignolet, J.; Li de la Sierra-Gallay, I.; Lazar, N.; Aumont-Nicaise, M.; Federle, M.J.; Prehna, G.; Hols, P.; et al. Structural insights into streptococcal competence regulation by the cell-to-cell communication system ComRS. PLoS Pathog. 2016, 12, e1005980. [Google Scholar] [CrossRef]
- Ledesma-García, L.; Ensinck, I.; Dereinne, D.; Viela, F.; Mignolet, J.; Dufrêne, Y.F.; Soumillion, P.; Nessler, S.; Hols, P. Coevolution of the bacterial pheromone ComS and sensor ComR fine-tunes natural transformation in streptococci. J. Biol. Chem. 2021, 297, 101346. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, Y.; Lu, M.; Lyu, X.; Gong, T.; Tang, B.; Wang, L.; Zeng, J.; Li, Y. Rhodiola rosea extract inhibits the biofilm formation and the expression of virulence genes of cariogenic oral pathogen Streptococcus mutans. Arch. Oral Biol. 2020, 116, 104762. [Google Scholar] [CrossRef]
- Balasubramanian, A.; Vasudevan, S.; Shanmugam, K.; Lévesque, C.; Solomon, A.; Neelakantan, P. Combinatorial effects of trans-cinnamaldehyde with fluoride and chlorhexidine on Streptococcus mutans. J. Appl. Microbiol. 2021, 130, 382–393. [Google Scholar] [CrossRef]
- Okada, A.; Igarashi, M.; Okajima, T.; Kinoshita, N.; Umekita, M.; Sawa, R.; Inoue, K.; Watanabe, T.; Doi, A.; Martin, A.; et al. Walkmycin B targets WalK (YycG), a histidine kinase essential for bacterial cell growth. J. Antibiot. 2010, 63, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, Y.; Kubo, N.; Matsunaga, H.; Igarashi, M.; Utsumi, R. Development of an antivirulence drug against Streptococcus mutans: Repression of biofilm formation, acid tolerance, and competence by a histidine kinase inhibitor, walkmycin C. Antimicrob. Agents Chemother. 2011, 55, 1475–1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, A.; Furukawa, S.; Fujita, S.; Mitobe, J.; Kawarai, T.; Narisawa, N.; Sekizuka, T.; Kuroda, M.; Ochiai, K.; Ogihara, H.; et al. Inhibition of Streptococcus mutans biofilm formation by Streptococcus salivarius FruA. Appl. Env. Microbiol. 2011, 77, 1572–1580. [Google Scholar] [CrossRef] [PubMed]
- Wasfi, R.; Abd El-Rahman, O.A.; Zafer, M.M.; Ashour, H.M. Probiotic Lactobacillus sp. inhibit growth, biofilm formation and gene expression of caries-inducing Streptococcus mutans. J. Cell Mol. Med. 2018, 22, 1972–1983. [Google Scholar] [CrossRef] [PubMed]
- Pourhajibagher, M.; Alaeddini, M.; Etemad-Moghadam, S.; Rahimi Esboei, B.; Bahrami, R.; Mousavi, M.; Bahador, A. Quorum quenching of Streptococcus mutans via the nano-quercetin-based antimicrobial photodynamic therapy as a potential target for cariogenic biofilm. BMC Microbiol. 2022, 22, 1–18. [Google Scholar] [CrossRef]
- Davis-Hanna, A.; Piispanen, A.E.; Stateva, L.I.; Hogan, D.A. Farnesol and dodecanol effects on the Candida albicans Ras1-cAMP signalling pathway and the regulation of morphogenesis. Mol. Microbiol. 2008, 67, 47–62. [Google Scholar] [CrossRef]
- Sebaa, S.; Boucherit-Otmani, Z.; Courtois, P. Effects of tyrosol and farnesol on Candida albicans biofilm. Mol. Med. Rep. 2019, 19, 3201–3209. [Google Scholar] [CrossRef]
- Kim, D.; Sengupta, A.; Niepa, T.H.; Lee, B.H.; Weljie, A.; Freitas-Blanco, V.S.; Murata, R.M.; Stebe, K.J.; Lee, D.; Koo, H. Candida albicans stimulates Streptococcus mutans microcolony development via cross-kingdom biofilm-derived metabolites. Sci. Rep. 2017, 7, 41332. [Google Scholar] [CrossRef]
- Bowen, W.H.; Koo, H. Biology of Streptococcus mutans-derived glucosyltransferases: Role in extracellular matrix formation of cariogenic biofilms. Caries Res. 2011, 45, 69–86. [Google Scholar] [CrossRef]
- Sztajer, H.; Szafranski, S.P.; Tomasch, J.; Reck, M.; Nimtz, M.; Rohde, M.; Wagner-Döbler, I. Cross-feeding and interkingdom communication in dual-species biofilms of Streptococcus mutans and Candida albicans. ISME J. 2014, 8, 2256–2271. [Google Scholar] [CrossRef]
- He, J.; Kim, D.; Zhou, X.; Ahn, S.J.; Burne, R.A.; Richards, V.P.; Koo, H. RNA-Seq reveals enhanced sugar metabolism in Streptococcus mutans co-cultured with Candida albicans within mixed-species biofilms. Front. Microbiol. 2017, 8, 1036. [Google Scholar] [CrossRef] [PubMed]
- Lobo, C.I.V.; Lopes, A.C.U.d.A.; Klein, M.I. Compounds with distinct targets present diverse antimicrobial and antibiofilm efficacy against Candida albicans and Streptococcus mutans, and combinations of compounds potentiate their effect. J. Fungi. 2021, 7, 340. [Google Scholar] [CrossRef] [PubMed]
- Priya, A.; Selvaraj, A.; Divya, D.; Karthik Raja, R.; Pandian, S.K. In vitro and in vivo anti-infective potential of thymol against early childhood caries causing dual species Candida albicans and Streptococcus mutans. Front. Pharmacol. 2021, 12, 760768. [Google Scholar] [CrossRef]
- Alemán-González-Duhart, D.; Álvarez-Almazán, S.; Valdes, M.; Tamay-Cach, F.; Mendieta-Wejebe, J.E. In vivo and ex vivo evaluation of 1,3-thiazolidine-2,4-dione derivatives as euglycemic agents. PPAR Res. 2021, 2021, 5100531. [Google Scholar] [CrossRef]
- Feldman, M.; Shenderovich, J.; Lavy, E.; Friedman, M.; Steinberg, D. A sustained-release membrane of thiazolidinedione-8: Effect on formation of a candida/bacteria mixed biofilm on hydroxyapatite in a continuous flow model. Biomed. Res. Int. 2017, 2017, 3510124. [Google Scholar] [CrossRef]
- Yost, S.; Duran-Pinedo, A.E.; Teles, R.; Krishnan, K.; Frias-Lopez, J. Functional signatures of oral dysbiosis during periodontitis progression revealed by microbial metatranscriptome analysis. Genome Med. 2015, 7, 27. [Google Scholar] [CrossRef] [PubMed]
- Frias, J.; Olle, E.; Alsina, M. Periodontal pathogens produce quorum sensing signal molecules. Infect. Immun. 2001, 69, 3431–3434. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G.; Darveau, R.P.; Curtis, M.A. The keystone-pathogen hypothesis. Nat. Rev. Microbiol. 2012, 10, 717–725. [Google Scholar] [CrossRef]
- Lamont, R.J.; Koo, H.; Hajishengallis, G. The oral microbiota: Dynamic communities and host interactions. Nat. Rev. Microbiol. 2018, 16, 745–759. [Google Scholar] [CrossRef]
- Socransky, S.S.; Haffajee, A.D.; Cugini, M.A.; Smith, C.; Kent, R.L., Jr. Microbial complexes in subgingival plaque. J. Clin. Periodontol. 1998, 25, 134–144. [Google Scholar] [CrossRef]
- Thurnheer, T.; Karygianni, L.; Flury, M.; Belibasakis, G.N. Fusobacterium species and subspecies differentially affect the composition and architecture of supra-and subgingival biofilms models. Front. Microbiol. 2019, 10, 1716. [Google Scholar] [CrossRef] [PubMed]
- Muras, A.; Mayer, C.; Otero-Casal, P.; Exterkate, R.A.M.; Brandt, B.W.; Crielaard, W.; Otero, A.; Krom, B.P. Short-chain N-acylhomoserine lactone quorum-sensing molecules promote periodontal pathogens in in vitro oral biofilms. Appl. Environ. Microbiol 2020, 86, e01941-19. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.J.; Choi, Y.J.; Lee, S.H.; Jun, H.K.; Choi, B.K. Autoinducer 2 of Fusobacterium nucleatum as a target molecule to inhibit biofilm formation of periodontopathogens. Arch. Oral Biol. 2013, 58, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.J.; Sim, J.; Jun, H.K.; Choi, B.K. Differential effect of autoinducer 2 of Fusobacterium nucleatum on oral streptococci. Arch. Oral Biol. 2013, 58, 1594–1602. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, W.; Song, Y.; Wang, J. Metatranscriptomic analysis of an in vitro biofilm model reveals strain-specific interactions among multiple bacterial species. J. Oral Microbiol. 2019, 11, 1599670. [Google Scholar] [CrossRef]
- Rupani, D.; Izano, E.A.; Schreiner, H.C.; Fine, D.H.; Kaplan, J.B. Aggregatibacter actinomycetemcomitans serotype f O-polysaccharide mediates coaggregation with Fusobacterium nucleatum. Oral Microbiol. Immunol. 2008, 23, 127–130. [Google Scholar] [CrossRef]
- Azakami, H.; Teramura, I.; Matsunaga, T.; Akimichi, H.; Noiri, Y.; Ebisu, S.; Kato, A. Characterization of autoinducer 2 signal in Eikenella corrodens and its role in biofilm formation. J. Biosci. Bioeng. 2006, 102, 110–117. [Google Scholar] [CrossRef]
- Szafrański, S.P.; Deng, Z.L.; Tomasch, J.; Jarek, M.; Bhuju, S.; Rohde, M.; Sztajer, H.; Wagner-Döbler, I. Quorum sensing of Streptococcus mutans is activated by Aggregatibacter actinomycetemcomitans and by the periodontal microbiome. BMC Genom. 2017, 18, 238. [Google Scholar] [CrossRef]
- Novak, E.A.; Shao, H.; Daep, C.A.; Demuth, D.R. Autoinducer-2 and QseC control biofilm formation and in vivo virulence of Aggregatibacter actinomycetemcomitans. Infect. Immun. 2010, 78, 2919–2926. [Google Scholar] [CrossRef]
- Ponnusamy, K.; Paul, D.; Sam Kim, Y.; Kweon, J.H. 2(5H)-Furanone: A prospective strategy for biofouling-control in membrane biofilm bacteria by quorum sensing inhibition. Braz. J. Microbiol. 2010, 41, 227–234. [Google Scholar] [CrossRef]
- Rasmussen, T.B.; Manefield, M.; Andersen, J.B.; Eberl, L.; Anthoni, U.; Christophersen, C.; Steinberg, P.; Kjelleberg, S.; Givskov, M. How Delisea pulchra furanones affect quorum sensing and swarming motility in Serratia liquefaciens MG1. Microbiology 2000, 146 Pt 12, 3237–3244. [Google Scholar] [CrossRef]
- Park, J.S.; Ryu, E.J.; Li, L.; Choi, B.K.; Kim, B.M. New bicyclic brominated furanones as potent autoinducer-2 quorum-sensing inhibitors against bacterial biofilm formation. Eur. J. Med. Chem. 2017, 137, 76–87. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, S.; Zhou, X.; Xui, Y. Quorum sensing inhibitor brominated furanone affects Porphyromonas gingivalis biofilm formation. West. China J. Stomatol. 2011, 29, 469–472. [Google Scholar]
- Manny, A.J.; Kjelleberg, S.; Kumar, N.; de Nys, R.; Read, R.W.; Steinberg, P. Reinvestigation of the sulfuric acid-catalysed cyclisation of brominated 2-alkyllevulinic acids to 3-alkyl-5-methylene-2 (5H)-furanones. Tetrahedron 1997, 53, 15813–15826. [Google Scholar] [CrossRef]
- Cho, Y.J.; Song, H.Y.; Ben Amara, H.; Choi, B.K.; Eunju, R.; Cho, Y.A.; Seol, Y.; Lee, Y.; Ku, Y.; Rhyu, I.C.; et al. In vivo inhibition of Porphyromonas gingivalis growth and prevention of periodontitis with quorum-sensing inhibitors. J. Periodontol. 2016, 87, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Ryu, E.J.; Sim, J.; Sim, J.; Lee, J.; Choi, B.K. D-Galactose as an autoinducer 2 inhibitor to control the biofilm formation of periodontopathogens. J. Microbiol. 2016, 54, 632–637. [Google Scholar] [CrossRef]
- Fournier-Larente, J.; Morin, M.P.; Grenier, D. Green tea catechins potentiate the effect of antibiotics and modulate adherence and gene expression in Porphyromonas gingivalis. Arch. Oral Biol. 2016, 65, 35–43. [Google Scholar] [CrossRef]
- Asahi, Y.; Noiri, Y.; Igarashi, J.; Asai, H.; Suga, H.; Ebisu, S. Effects of N-acyl homoserine lactone analogues on Porphyromonas gingivalis biofilm formation. J. Periodontal. Res. 2010, 45, 255–261. [Google Scholar] [CrossRef]
- Elani, H.W.; Starr, J.R.; Da Silva, J.D.; Gallucci, G.O. Trends in dental implant use in the U.S., 1999–2016, and projections to 2026. J. Dent. Res. 2018, 97, 1424–1430. [Google Scholar] [CrossRef]
- Ramachandra, S.S.; Patil, M.; Mehta, D.S. Implants placed into extraction sockets: A literature review. Dent. Implant. Update 2009, 20, 1–8. [Google Scholar]
- Ramachandra, S.S.; Rana, R.; Reetika, S.; Jithendra, K.D. Options to avoid the second surgical site: A review of literature. Cell Tissue Bank 2014, 15, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Heitz-Mayfield, L.J.A.; Salvi, G.E. Peri-implant mucositis. J. Clin. Periodontol. 2018, 45 (Suppl. 20), S237–S245. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, F.; Derks, J.; Monje, A.; Wang, H.L. Peri-implantitis. J. Clin. Periodontol. 2018, 45 (Suppl. 20), S246–S266. [Google Scholar] [CrossRef] [PubMed]
- Renvert, S.; Persson, G.R.; Pirih, F.Q.; Camargo, P.M. Peri-implant health, peri-implant mucositis, and peri-implantitis: Case definitions and diagnostic considerations. J. Clin. Periodontol. 2018, 45 (Suppl. 20), S278–S285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolae, V.D.; Chiscop, I.; Cioranu, V.S.I.; Mârțu, M.-A.; Luchian, A.I.; Mârțu, S.; Solomon, S.M. The use of photoactivated blue-O toluidine for periimplantitis treatment in patients with periodontal disease. Rev. Chim. (Buchar.) 2016, 66, 2121–2123. [Google Scholar]
- Kang, M.; Kim, S.; Kim, H.; Song, Y.; Jung, D.; Kang, S.; Seo, J.H.; Nam, S.; Lee, Y. Calcium-binding polymer-coated poly(lactide- co-glycolide) microparticles for sustained release of quorum sensing inhibitors to prevent biofilm formation on hydroxyapatite surfaces. ACS Appl. Mater. Interfaces 2019, 11, 7686–7694. [Google Scholar] [CrossRef]
- An, S.J.; Namkung, J.U.; Ha, K.W.; Jun, H.K.; Kim, H.Y.; Choi, B.K. Inhibitory effect of D-arabinose on oral bacteria biofilm formation on titanium discs. Anaerobe 2022, 102533. [Google Scholar] [CrossRef]
- Rajkumar, D.; Rubini, D.; Sudharsan, M.; Suresh, D.; Nithyanand, P. Novel thiazolinyl-picolinamide based palladium(II) complex-impregnated urinary catheters quench the virulence and disintegrate the biofilms of uropathogens. Biofouling 2020, 36, 351–367. [Google Scholar] [CrossRef]
- Jothipandiyan, S.; Suresh, D.; Sankaran, S.V.; Thamotharan, S.; Shanmugasundaram, K.; Vincent, P.; Sekaran, S.; Gowrishankar, S.; Pandian, S.K.; Paramasivam, N. Heteroleptic pincer palladium(II) complex coated orthopedic implants impede the AbaI/AbaR quorum sensing system and biofilm development by Acinetobacter baumannii. Biofouling 2022, 38, 55–70. [Google Scholar] [CrossRef]
- Ganesan, S.M.; Dabdoub, S.M.; Nagaraja, H.N.; Mariotti, A.J.; Ludden, C.W.; Kumar, P.S. Biome-microbiome interactions in peri-implantitis: A pilot investigation. J. Periodontol. 2022, 93, 814–823. [Google Scholar] [CrossRef]
- Kumar, P.S.; Mason, M.R.; Brooker, M.R.; O’Brien, K. Pyrosequencing reveals unique microbial signatures associated with healthy and failing dental implants. J. Clin. Periodontol. 2012, 39, 425–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wright, P.P.; Ramachandra, S.S. Quorum Sensing and Quorum Quenching with a Focus on Cariogenic and Periodontopathic Oral Biofilms. Microorganisms 2022, 10, 1783. https://doi.org/10.3390/microorganisms10091783
Wright PP, Ramachandra SS. Quorum Sensing and Quorum Quenching with a Focus on Cariogenic and Periodontopathic Oral Biofilms. Microorganisms. 2022; 10(9):1783. https://doi.org/10.3390/microorganisms10091783
Chicago/Turabian StyleWright, Patricia P., and Srinivas Sulugodu Ramachandra. 2022. "Quorum Sensing and Quorum Quenching with a Focus on Cariogenic and Periodontopathic Oral Biofilms" Microorganisms 10, no. 9: 1783. https://doi.org/10.3390/microorganisms10091783
APA StyleWright, P. P., & Ramachandra, S. S. (2022). Quorum Sensing and Quorum Quenching with a Focus on Cariogenic and Periodontopathic Oral Biofilms. Microorganisms, 10(9), 1783. https://doi.org/10.3390/microorganisms10091783