



Probiotics and Postbiotics as the Functional Food Components Affecting the Immune Response


Abstract
:1. Introduction
2. Probiotics and Host’s Immune System
2.1. Concept of Probiotics
2.2. Mucosal Immune System
2.3. The Immune Regulation by Probiotics
2.4. Effect of Immunomodulation by Selected Probiotics
3. Post-Immunobiotics
3.1. Immunomodulatory Effects of Postbiotics—General Remarks and Health-Promoting Benefits
3.2. The Postbiotics’ Impact on Immune Homeostasis
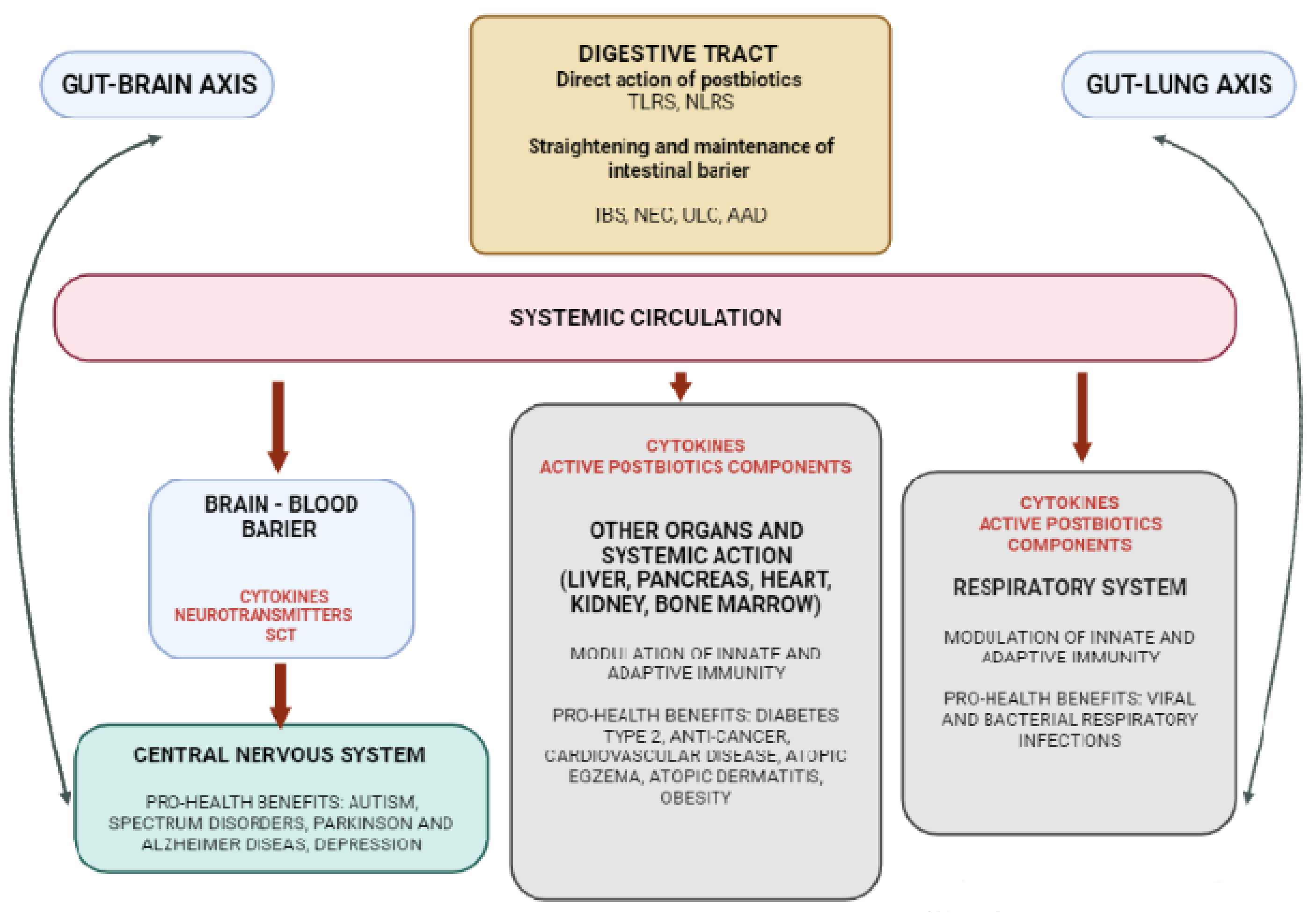
3.3. Postbiotics—The Role of Gut–Organ Axis on the Immune System Modulation
3.4. Safety of Postbiotics
4. Immunobiotics and Post-Immunobiotics as the Components of Functional Food
4.1. Immunobiotics in the Aspect of Functional Food
4.2. Technological Factors in Probiotic Food Development
4.3. New Sources of Probiotic Microorganisms
4.4. Functional Food Products with Post-Immunobiotics
Aspect of Sensory Quality
5. Future Trends and Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Definition of Functional Food. Available online: https://www.functionalfoodscenter.net/ (accessed on 13 September 2022).
- IOM/NAS. Opportunities In The Nutrition and Food Sciences; Thomas, P.R., Earl, R., Eds.; Institute of Medicine/National Academy of Sciences: Washington, DC, USA; National Academy Press: Washington, DC, USA, 1994; p. 109. [Google Scholar]
- IFIS. Functional foods. In Food Insight Media Guide; International Food Information Council Foundation: Washington, DC, USA, 1998. [Google Scholar]
- Scientific Concepts of Functional Foods in Europe Consensus Document. Br. J. Nutr. 1999, 81, S1–S27. [CrossRef] [Green Version]
- GAO. Food safety. In Improvements Needed in Overseeing the Safety of Dietary Supplements and “Functional Foods”; Report to Congressional Committees (GAO/RCED-00-156); Congressional Committees: Washington, DC, USA, 2000. [Google Scholar]
- IFIC. Functional Foods for Health Consumer Trending Survey. 2011. Available online: https://www.foodinsight.org/2011 (accessed on 12 August 2022).
- Martirosyan, D.M.; Singh, J. A new definition of functional food by FFC: What makes a new definition unique? Funct. Foods Health Dis. 2015, 5, 183. [Google Scholar] [CrossRef]
- Ashaolu, T.J. A review on selection of fermentative microorganisms for functional foods and beverages: The production and future perspectives. Int. J. Food Sci. Technol. 2019, 54, 2511–2519. [Google Scholar] [CrossRef]
- Dalonso, N.; Goldman, G.H.; Gern, R.M.M. β-(1→3),(1→6)-Glucans: Medicinal activities, characterization, biosynthesis and new horizons. Appl. Microbiol. Biotechnol. 2015, 99, 7893–7906. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, D.H.; Jo, S.; Cho, M.J.; Cho, Y.R.; Lee, Y.J.; Byun, S. Immunomodulatory functional foods and their molecular mechanisms. Exp. Mol. Med. 2022, 54, 11. [Google Scholar] [CrossRef]
- Vitetta, L.; Vitetta, G.; Hall, S. Immunological Tolerance and Function: Associations Between Intestinal Bacteria, Probiotics, Prebiotics, and Phages. Front. Immunol. 2018, 9, 2240. [Google Scholar] [CrossRef] [Green Version]
- Duda-Chodak, A.; Tarko, T.; Satora, P.; Sroka, P. Interaction of dietary compounds, especially polyphenols, with the intestinal microbiota: A review. Eur. J. Nutr. 2015, 54, 325–341. [Google Scholar] [CrossRef] [Green Version]
- Kocot, A.M.; Jarocka-Cyrta, E.; Drabińska, N. Overview of the Importance of Biotics in Gut Barrier Integrity. Int. J. Mol. Sci. 2022, 23, 2896. [Google Scholar] [CrossRef]
- Childs, C.E.; Calder, P.C.; Miles, E.A. Diet and Immune Function. Nutrients 2019, 11, 1933. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The international scientific association for probiotics and prebiotics consensus statement on the scope and ap-propriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Villena, J.; Aso, H.; Rutten, V.P.M.G.; Takahashi, H.; van Eden, W.; Kitazawa, H. Immunobiotics for the Bovine Host: Their Interaction with Intestinal Epithelial Cells and Their Effect on Antiviral Immunity. Front. Immunol. 2018, 9, 326. [Google Scholar] [CrossRef] [Green Version]
- Cristofori, F.; Dargenio, V.N.; Dargenio, C.; Miniello, V.L.; Barone, M.; Francavilla, R. Anti-Inflammatory and Immunomodulatory Effects of Probiotics in Gut Inflammation: A Door to the Body. Front. Immunol. 2021, 12, 578386. [Google Scholar] [CrossRef]
- Vlasova, A.N.; Kandasamy, S.; Chattha, K.S.; Rajashekara, G.; Saif, L.J. Comparison of probiotic lactobacilli and bifidobacteria effects, immune responses and rotavirus vaccines and infection in different host species. Veter-Immunol. Immunopathol. 2016, 172, 72–84. [Google Scholar] [CrossRef] [Green Version]
- Kerry, R.G.; Patra, J.K.; Gouda, S.; Park, Y.; Shin, H.-S.; Das, G. Benefaction of probiotics for human health: A review. J. Food Drug Anal. 2018, 26, 927–939. [Google Scholar] [CrossRef] [Green Version]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Sharma, M.; Shukla, G. Metabiotics: One Step ahead of Probiotics; an Insight into Mechanisms Involved in Anticancerous Effect in Colorectal Cancer. Front. Microbiol. 2016, 7, 1940. [Google Scholar] [CrossRef] [Green Version]
- Shenderov, B.A. Metabiotics: Novel idea or natural development of probiotic conception. Microb. Ecol. Heal. Dis. 2013, 24, 20399. [Google Scholar] [CrossRef]
- Vinderola, G.; Sanders, M.E.; Salminen, S. The Concept of Postbiotics. Foods 2022, 11, 1077. [Google Scholar] [CrossRef]
- Aguilar-Toalá, J.E.; Garcia-Varela, R.; Garcia, H.S.; Mata-Haro, V.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol. 2018, 75, 105–114. [Google Scholar] [CrossRef]
- Collado, M.; Vinderola, G.; Salminen, S. Postbiotics: Facts and open questions. A position paper on the need for a consensus definition. Benef. Microbes 2019, 10, 711–719. [Google Scholar] [CrossRef]
- Da Silva, T.F.; Casarotti, S.; De Oliveira, G.L.V.; Penna, A.L.B. The impact of probiotics, prebiotics, and synbiotics on the biochemical, clinical, and immunological markers, as well as on the gut microbiota of obese hosts. Crit. Rev. Food Sci. Nutr. 2020, 61, 337–355. [Google Scholar] [CrossRef] [PubMed]
- Rajpoot, M.; Sharma, A.K.; Sharma, A.; Gupta, G.K. Understanding the microbiome: Emerging biomarkers for exploiting the microbiota for personalized medicine against cancer. Semin. Cancer Biol. 2018, 52, 8. [Google Scholar] [CrossRef] [PubMed]
- Yeşilyurt, N.; Yılmaz, B.; Ağagündüz, D.; Capasso, R. Microbiome-based personalized nutrition as a result of the 4.0 technological revolution: A mini literature review. Process. Biochem. 2022, 121, 257–262. [Google Scholar] [CrossRef]
- Hughes, R.L.; Marco, M.L.; Hughes, J.P.; Keim, N.L.; Kable, M.E. The Role of the Gut Microbiome in Predicting Response to Diet and the Development of Precision Nutrition Models—Part I: Overview of Current Methods. Adv. Nutr. Int. Rev. J. 2019, 10, 953–978. [Google Scholar] [CrossRef]
- Miranda, J.M.; Anton, X.; Redondo-Valbuena, C.; Roca-Saavedra, P.; Rodriguez, J.A.; Lamas, A.; Franco, C.M.; Cepeda, A. Egg and Egg-Derived Foods: Effects on Human Health and Use as Functional Foods. Nutrients 2015, 7, 706–729. [Google Scholar] [CrossRef]
- Doan, T.; Melvold, R.; Viselli, S.; Waltenbaugh, C. Lippincott’s Illustrated Reviews: Immunology. In The Need to Know Your Self; Harvey, R.A., Ed.; Nobel Tıp Kitabevleri: İstanbul, Turkey, 2017; pp. 3–10. [Google Scholar]
- Pradeu, T.; Cooper, E.L. The danger theory: 20 years later. Front. Immunol. 2012, 3, 287. [Google Scholar] [CrossRef] [Green Version]
- Seong, S.-Y.; Matzinger, P. Hydrophobicity: An ancient damage-associated molecular pattern that initiates innate immune responses. Nat. Rev. Immunol. 2004, 4, 469–478. [Google Scholar] [CrossRef]
- Yu, Y.; Dunaway, S.; Champer, J.; Kim, J.; Alikhan, A. Changing our microbiome: Probiotics in dermatology. Br. J. Dermatol. 2019, 182, 39–46. [Google Scholar] [CrossRef]
- Ray, A.; Dittel, B.N. Interrelatedness between dysbiosis in the gut microbiota due to immunodeficiency and disease penetrance of colitis. Immunology 2015, 146, 359–368. [Google Scholar] [CrossRef] [Green Version]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino—Michaels, K.; Earley, Z.M.; Benyamin, F.W.; Lei, Y.M.; Jabri, B.; Alegre, M.L.; et al. Commensal Bifidobacterium promotes antitumor immunity and faciliyayes anti-PD-L1 efficacy. Science 2015, 350, 1084–1089. [Google Scholar] [CrossRef]
- Vieira, A.T.; Teixeira, M.M.; Martins, F.D.S. The Role of Probiotics and Prebiotics in Inducing Gut Immunity. Front. Immunol. 2013, 4, 445. [Google Scholar] [CrossRef]
- Hardy, H.; Harris, J.; Lyon, E.; Beal, J.; Foey, A.D. Probiotics, Prebiotics and Immunomodulation of Gut Mucosal Defences: Homeostasis and Immunopathology. Nutrients 2013, 5, 1869–1912. [Google Scholar] [CrossRef]
- Brandtzaeg, P. Chapter 1.2-Immunity in the Gut: Mechanisms and Functions; Lennart, S., Dessel-berger, U., Harry, B., Greenberg, M.K.E., Viral, G., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 23–46. [Google Scholar] [CrossRef]
- Galdeano, C.M.; Perdigon, G. Role of viability of probiotic strains in their persistence in the gut and in mucosal immune stimulation. J. Appl. Microbiol. 2004, 97, 673–681. [Google Scholar] [CrossRef]
- Dhanapal, A.P. Genomics of crop plant genetic resources. Adv. Biosci. Biotechnol. 2012, 03, 378–385. [Google Scholar] [CrossRef] [Green Version]
- Foligné, B.; Dewulf, J.; Breton, J.; Claisse, O.; Lonvaud-Funel, A.; Pot, B. Probiotic properties of non-conventional lactic acid bacteria: Immunomodulation by Oenococcus oeni. Int. J. Food Microbiol. 2010, 140, 136–145. [Google Scholar] [CrossRef]
- Chiba, Y.; Shida, K.; Nagata, S.; Wada, M.; Bian, L.; Wang, C.; Shimizu, T.; Yamashiro, Y.; Kiyoshima-Shibata, J.; Nanno, M.; et al. Well-controlled proinflammatory cytokine responses of Peyer’s patch cells to probiotic Lactobacillus casei. Immunology 2010, 130, 352–362. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Saruta, J.; Takahashi, T.; To, M.; Shimizu, T.; Hayashi, T.; Morozumi, T.; Kubota, N.; Kamata, Y.; Makino, S.; et al. Effect of ingesting yogurt fermented with Lactobacillus delbrueckii ssp. bulgaricus OLL1073R-1 on influenza virus-bound salivary IgA in elderly residents of nursing homes: A randomized controlled trial. Acta Odontol. Scand. 2019, 77, 517–524. [Google Scholar] [CrossRef]
- Pu, F.; Guo, Y.; Li, M.; Zhu, H.; Wang, S.; Shen, X.; He, M.; Huang, C.; He, F. Yogurt supplemented with probiotics can protect the healthy elderly from respiratory infections: A randomized controlled open-label trial. Clin. Interv. Aging 2017, 12, 1223–1231. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, S.; Hattori, M.; Sugiyama, H.; Hanaoka, A.; Okada, S.; Yoshida, T. Lactobacillus plantarum NRIC1832 Enhances IL-10 Production from CD4+T Cells in Vitro. Biosci. Biotechnol. Biochem. 2012, 76, 1925–1931. [Google Scholar] [CrossRef]
- Yoshida, T.; Enomoto, M.; Nakayama, S.; Adachi, Y.; Fujiwara, W.; Sugiyama, H.; Shimojoh, M.; Okada, S.; Hattori, M. Induction of ALDH Activity in Intestinal Dendritic Cells by Lactobacillus plantarum NRIC0380. Biosci. Biotechnol. Biochem. 2013, 77, 1826–1831. [Google Scholar] [CrossRef]
- Albarracin, L.; Kobayashi, H.; Iida, H.; Sato, N.; Nochi, T.; Aso, H.; Salva, S.; Alvarez, S.; Kitazawa, H.; Villena, J. Transcriptomic Analysis of the Innate Antiviral Immune Response in Porcine Intestinal Epithelial Cells: Influence of Immunobiotic Lactobacilli. Front. Immunol. 2017, 8, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki-Yoshida, A.; Yamada, K.; Hachimura, S.; Sashihara, T.; Ikegami, S.; Shimizu, M.; Totsuka, M. Enhancement of Oral Tolerance Induction in DO11.10 Mice by Lactobacillus gasseri OLL2809 via Increase of Effector Regulatory T Cells. PLoS ONE 2016, 11, e0158643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishizuka, T.; Kanmani, P.; Kobayashi, H.; Miyazaki, A.; Soma, J.; Suda, Y.; Aso, H.; Nochi, T.; Iwabuchi, N.; Xiao, J.-Z.; et al. Immunobiotic Bifidobacteria Strains Modulate Rotavirus Immune Response in Porcine Intestinal Epitheliocytes via Pattern Recognition Receptor Signaling. PLoS ONE 2016, 11, e0152416. [Google Scholar] [CrossRef] [PubMed]
- Badr, H.; El-Baz, A.; Mohamed, I.; Shetaia, Y.; El-Sayed, A.S.A.; Sorour, N. Bioprocess optimization of glutathione production by Saccharomyces boulardii: Biochemical characterization of glutathione peroxidase. Arch. Microbiol. 2021, 203, 6183–6196. [Google Scholar] [CrossRef] [PubMed]
- Fric, P. Probiotics and prebiotics—Renaissance of a therapeutic principle. Open Med. 2007, 2, 237–270. [Google Scholar] [CrossRef]
- Valdovinos-García, L.; Abreu, A.; Valdovinos-Díaz, M. Uso de probióticos en la práctica clínica: Resultados de una encuesta nacional a gastroenterólogos y nutriólogos. Rev. Gastroenterol. México 2019, 84, 303–309. [Google Scholar] [CrossRef]
- Abraham, D.; Feher, J.; Scuderi, G.L.; Szabo, D.; Dobolyi, A.; Cservenak, M.; Juhasz, J.; Ligeti, B.; Pongor, S.; Gomez-Cabrera, M.C.; et al. Exercise and probiotics attenuate the development of Alzheimer’s disease in transgenic mice: Role of microbiome. Exp. Gerontol. 2019, 115, 122–131. [Google Scholar] [CrossRef]
- Mohanty, D.; Panda, S.; Kumar, S.; Ray, P. In vitro evaluation of adherence and anti-infective property of probiotic Lactobacillus plantarum DM 69 against Salmonella enterica. Microb. Pathog. 2019, 126, 212–217. [Google Scholar] [CrossRef]
- Sakandar, H.A.; Kubow, S.; Sadiq, F.A. Isolation and in-vitro probiotic characterization of fructophilic lactic acid bacteria from Chinese fruits and flowers. Lwt 2019, 104, 70–75. [Google Scholar] [CrossRef]
- Chahibakhsh, N.; Hosseini, E.; Lslam, S.; Rahbar, A.R. Bitter almond gum reduces body mass index, serum triglyceride, hyperinsulinemia and insulin resistance in overweight subjects with hyperlipidemia. J. Funct. Foods 2019, 55, 343–351. [Google Scholar] [CrossRef]
- Kolaček, S.; Hojsak, I.; Berni Canani, R.; Guarino, A.; Indrio, F.; Orel, R.; Pot, B.; Shamir, R.; Szajewska, H.; Vandenplas, Y.; et al. Commercial Probiotic Products: A Call for Improved Quality Control. A Position Paper by the ESPGHAN Working Group for Probiotics and Prebiotics. J. Pediatr. Gastroenterol. Nutr. 2017, 65, 117–124. [Google Scholar] [CrossRef]
- Song, M.; Yun, B.; Moon, J.-H.; Park, D.-J.; Lim, K.; Oh, S. Characterization of Selected Lactobacillus Strains for Use as Probiotics. Korean J. Food Sci. Anim. Resour. 2015, 35, 551–556. [Google Scholar] [CrossRef] [Green Version]
- Wan, M.L.Y.; Ling, K.H.; El-Nezami, H.; Wang, M.F. Influence of functional food components on gut health. Crit. Rev. Food Sci. Nutr. 2018, 59, 1927–1936. [Google Scholar] [CrossRef]
- Hepworth, M.R.; Monticelli, L.A.; Fung, T.C.; Ziegler, C.G.K.; Grunberg, S.; Sinha, R.; Mantegazza, A.R.; Ma, H.-L.; Crawford, A.; Angelosanto, J.M.; et al. Innate lymphoid cells regulate CD4+ T-cell responses to intestinal commensal bacteria. Nature 2013, 498, 113–117. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Santamarina, A.; Gonzalez, E.; Lamas, A.; Mondragon, A.; Regal, P.; Miranda, J. Probiotics as a Possible Strategy for the Prevention and Treatment of Allergies. A Narrative Review. Foods 2021, 10, 701. [Google Scholar] [CrossRef]
- Dennis-Wall, J.C.; Culpepper, T.; Nieves, C., Jr.; Rowe, C.C.; Burns, A.M.; Rusch, C.T.; Federico, A.; Ukhanova, M.; Waugh, S.; Mai, V.; et al. Probiotics (Lactobacillus gasseri KS-13, Bifidobacterium bifidum G9-1, and Bifidobacterium longum MM-2) improve rhinoconjunctivitis-specific quality of life in individuals with seasonal allergies: A double-blind, placebo-controlled, randomized trial. Am. J. Clin. Nutr. 2017, 105, 758–767. [Google Scholar] [CrossRef] [Green Version]
- Feyisetan, O.; Tracey, C.; Hellawell, G.O. Probiotics, dendritic cells and bladder cancer. BJU Int. 2011, 109, 1594–1597. [Google Scholar] [CrossRef]
- Xie, Z.; Cai, J.; Sun, W.; Hua, S.; Wang, X.; Li, A.; Jiang, J. Development and Validation of Prognostic Model in Transitional Bladder Cancer Based on Inflammatory Response-Associated Genes. Front. Oncol. 2021, 11, 740985. [Google Scholar] [CrossRef]
- Popova, M.; Molimard, P.; Courau, S.; Crociani, J.; Dufour, C.; Le Vacon, F.; Carton, T. Beneficial effects of probiotics in upper respiratory tract infections and their mechanical actions to antagonize pathogens. J. Appl. Microbiol. 2012, 113, 1305–1318. [Google Scholar] [CrossRef]
- Picó-Monllor, J.A.; Ruzafa-Costas, B.; Núñez-Delegido, E.; Sánchez-Pellicer, P.; Peris-Berraco, J.; Navarro-Lopez, V. Selection of Probiotics in the Prevention of Respiratory Tract Infections and Their Impact on Occupational Health: Scoping Review. Nutrients 2021, 13, 4419. [Google Scholar] [CrossRef]
- WHO COVID-19 Weekly Epidemiological Update World Health Organization. 2022. Available online: https://www.who.int/publications/m/item/weekly-epidemiological-update-on-covid-19---18-january-2022 (accessed on 20 September 2022).
- Nguyen, Q.V.; Chong, L.C.; Hor, Y.-Y.; Lew, L.-C.; Rather, I.A.; Choi, S.-B. Role of Probiotics in the Management of COVID-19: A Computational Perspective. Nutrients 2022, 14, 274. [Google Scholar] [CrossRef] [PubMed]
- Doron, S.; Snydman, D.R. Risk and Safety of Probiotics. Clin. Infect. Dis. 2015, 60, S129–S134. [Google Scholar] [CrossRef] [Green Version]
- Rannikko, J.; Holmberg, V.; Karppelin, M.; Arvola, P.; Huttunen, R.; Mattila, E.; Kerttula, N.; Puhto, T.; Tamm, Ü.; Koivula, I. Fungemia and Other Fungal Infections Associated with Use of Saccharomyces boulardii Probiotic Supplements. Emerg. Infect. Dis. 2021, 27, 2090–2096. [Google Scholar] [CrossRef] [PubMed]
- Kareem, K.Y.; Ling, F.H.; Chwen, L.T.; Foong, O.M.; Asmara, S.A. Inhibitory activity of postbiotic produced by strains of Lactobacillus plantarum using reconstituted media supplemented with inulin. Gut Pathog. 2014, 6, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haileselassie, Y.; Navis, M.; Vu, N.; Qazi, K.R.; Rethi, B.; Sverremark-Ekström, E. Postbiotic Modulation of Retinoic Acid Imprinted Mucosal-like Dendritic Cells by Probiotic Lactobacillus reuteri 17938 In Vitro. Front. Immunol. 2016, 7, 96. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, F.; Ishida, Y.; Sawada, D.; Ashida, N.; Sugawara, T.; Sakai, M.; Goto, T.; Kawada, T.; Fujiwara, S. Fragmented Lactic Acid Bacterial Cells Activate Peroxisome Proliferator-Activated Receptors and Ameliorate Dyslipidemia in Obese Mice. J. Agric. Food Chem. 2016, 64, 2549–2559. [Google Scholar] [CrossRef]
- Tiptiri-Kourpeti, A.; Spyridopoulou, K.; Santarmaki, V.; Aindelis, G.; Tompoulidou, E.; Lamprianidou, E.E.; Saxami, G.; Ypsilantis, P.; Lampri, E.S.; Simopoulos, C.; et al. Lactobacillus casei Exerts Anti-Proliferative Effects Accompanied by Apoptotic Cell Death and Up-Regulation of TRAIL in Colon Carcinoma Cells. PLoS ONE 2016, 11, e0147960. [Google Scholar] [CrossRef]
- da Costa Waite, C.C.; da Silva, G.O.A.; Bitencourt, J.A.P.; Sabadini-Santos, E.; Crapez, M.A.C. Copper and lead removal from aqueous solutions by bacterial consortia acting as biosorbents. Marine Pollut. Bull. 2016, 109, 386–392. [Google Scholar] [CrossRef]
- Konstantinov, S.R.; Kuipers, E.J.; Peppelenbosch, M. Functional genomic analyses of the gut microbiota for CRC screening. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 741–745. [Google Scholar] [CrossRef]
- Tsilingiri, K.; Rescigno, M. Postbiotics: What else? Benef. Microbes 2013, 4, 101–107. [Google Scholar] [CrossRef]
- Peluzio, M.D.C.G.; Martinez, J.A.; Milagro, F.I. Postbiotics: Metabolites and mechanisms involved in microbiota-host interactions. Trends Food Sci. Technol. 2020, 108, 11–26. [Google Scholar] [CrossRef]
- Piqué, N.; Berlanga, M.; Miñana-Galbis, D. Health Benefits of Heat-Killed (Tyndallized) Probiotics: An Overview. Int. J. Mol. Sci. 2019, 20, 2534. [Google Scholar] [CrossRef] [Green Version]
- Żółkiewicz, J.; Marzec, A.; Ruszczyński, M.; Feleszko, W. Postbiotics—A Step Beyond Pre- and Probiotics. Nutrients 2020, 12, 2189. [Google Scholar] [CrossRef]
- Dang, A.T.; Marsland, B.J. Microbes, metabolites, and the gut–lung axis. Mucosal Immunol. 2019, 12, 843–850. [Google Scholar] [CrossRef] [Green Version]
- Hou, H.; Chen, D.; Zhang, K.; Zhang, W.; Liu, T.; Wang, S.; Dai, X.; Wang, B.; Zhong, W.; Cao, H. Gut microbiota-derived short-chain fatty acids and colorectal cancer: Ready for clinical translation? Cancer Lett. 2021, 526, 225–235. [Google Scholar] [CrossRef]
- Ichinohe, T.; Pang, I.K.; Kumamoto, Y.; Peaper, D.R.; Ho, J.H.; Murray, T.S.; Iwasaki, A. Microbiota regulates immune defense against respiratory tract influenza A virus infection. Proc. Natl. Acad. Sci. USA 2011, 108, 5354–5359. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Du, W.; Ni, Y.; Lan, G.; Shi, G. The effect of short-chain fatty acids on M2 macrophages polarization in vitro and in vivo. Clin. Exp. Immunol. 2021, 207, 53–64. [Google Scholar] [CrossRef]
- Chudzik, A.; Orzyłowska, A.; Rola, R.; Stanisz, G. Probiotics, Prebiotics and Postbiotics on Mitigation of Depression Symptoms: Modulation of the Brain–Gut–Microbiome Axis. Biomolecules 2021, 11, 1000. [Google Scholar] [CrossRef]
- Bravo, J.A.; Julio-Pieper, M.; Forsythe, P.; Kunze, W.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Communication between gastrointestinal bacteria and the nervous system. Curr. Opin. Pharmacol. 2012, 12, 667–672. [Google Scholar] [CrossRef]
- Haghikia, A.; Jörg, S.; Duscha, A.; Berg, J.; Manzel, A.; Waschbisch, A.; Hammer, A.; Lee, D.-H.; May, C.; Wilck, N.; et al. Dietary Fatty Acids Directly Impact Central Nervous System Autoimmunity via the Small Intestine. Immunity 2015, 43, 817–829. [Google Scholar] [CrossRef] [Green Version]
- Belkaid, Y.; Harrison, O.J. Homeostatic Immunity and the Microbiota. Immunity 2017, 46, 562–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serhan, C.N.; Savill, J. Resolution of inflammation: The beginning programs the end. Nat. Immunol. 2005, 6, 1191–1197. [Google Scholar] [CrossRef] [PubMed]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of Antioxidants and Natural Products in Inflammation. Oxid. Med. Cell. Longev. 2016, 2016, 5276130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bermudez-Brito, M.; Plaza-Díaz, J.; Muñoz-Quezada, S.; Gómez-Llorente, C.; Gil, A. Probiotic Mechanisms of Action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Hoang, T.K.; He, B.; Wang, T.; Tran, D.Q.; Rhoads, J.M.; Liu, Y. Protective effect of Lactobacillus reuteri DSM 17938 against experimental necrotizing enterocolitis is mediated by Toll-like receptor 2. Am. J. Physiol. Liver Physiol. 2018, 315, G231–G240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menard, S.; Candalh, C.; Bambou, J.C.; Terpend, K.; Cerf-Bensussan, N.; Heyman, M. Lactic acid bacteria secrete metabolites retaining anti-inflammatory properties after intestinal transport. Gut 2004, 53, 821–828. [Google Scholar] [CrossRef]
- Yu, S.; Gao, N. Compartmentalizing intestinal epithelial cell toll-like receptors for immune surveillance. Cell. Mol. Life Sci. 2015, 72, 3343–3353. [Google Scholar] [CrossRef] [Green Version]
- Zaki, M.H.; Boyd, K.L.; Vogel, P.; Kastan, M.B.; Lamkanfi, M.; Kanneganti, T.-D. The NLRP3 Inflammasome Protects against Loss of Epithelial Integrity and Mortality during Experimental Colitis. Immunity 2010, 32, 379–391. [Google Scholar] [CrossRef] [Green Version]
- De Marco, S.; Sichetti, M.; Muradyan, D.; Piccioni, M.; Traina, G.; Pagiotti, R.; Pietrella, D. Probiotic Cell-Free Supernatants Exhibited Anti-Inflammatory and Antioxidant Activity on Human Gut Epithelial Cells and Macrophages Stimulated with LPS. Evidence-Based Complement. Altern. Med. 2018, 2018, 1756308. [Google Scholar] [CrossRef]
- Gill, P.A.; Van Zelm, M.C.; Muir, J.G.; Gibson, P.R. Review article: Short chain fatty acids as potential therapeutic agents in human gastrointestinal and inflammatory disorders. Aliment. Pharmacol. Ther. 2018, 48, 15–34. [Google Scholar] [CrossRef] [Green Version]
- Nagai, Y.; Garrett, K.P.; Ohta, S.; Bahrun, U.; Kouro, T.; Akira, S.; Takatsu, K.; Kincade, P.W. Toll-like receptors on hematopoietic progenitor cells stimulate innate immune system replenishment. Immunity 2006, 24, 801–812. [Google Scholar] [CrossRef] [Green Version]
- Thorakkattu, P.; Khanashyam, A.C.; Shah, K.; Babu, K.S.; Mundanat, A.S.; Deliephan, A.; Deokar, G.S.; Santivarangkna, C.; Nirmal, N.P. Postbiotics: Current Trends in Food and Pharmaceutical Industry. Foods 2022, 11, 3094. [Google Scholar] [CrossRef]
- Jurášková, D.; Ribeiro, S.C.; Silva, C.C.G. Exopolysaccharides Produced by Lactic Acid Bacteria: From Biosynthesis to Health-Promoting Properties. Foods 2022, 11, 156. [Google Scholar] [CrossRef]
- Gezginç, Y.; Karabekmez-Erdem, T.; Tatar, H.D.; Ayman, S.; Ganiyusufoğlu, E.; Dayisoylu, K.S. Health promoting benefits of postbiotics produced by lactic acid bacteria: Exopolysaccharide. Biotech Stud. 2022, 31, 62–63. [Google Scholar] [CrossRef]
- Saeui, C.T.; Urias, E.; Liu, L.; Mathew, M.P.; Yarema, K.J. Metabolic glycoengineering bacteria for therapeutic, recombinant protein, and metabolite production applications. Glycoconj. J. 2015, 32, 425–441. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; MacLeod, D.T.; Di Nardo, A. Commensal Bacteria Lipoteichoic Acid Increases Skin Mast Cell Antimicrobial Activity against Vaccinia Viruses. J. Immunol. 2012, 189, 1551–1558. [Google Scholar] [CrossRef] [Green Version]
- Zadeh, M.; Khan, M.W.; Goh, Y.J.; Selle, K.; Owen, J.L.; Klaenhammer, T.; Mohamadzadeh, M. Induction of intestinal pro-inflammatory immune responses by lipoteichoic acid. J. Inflamm. 2012, 9, 7. [Google Scholar] [CrossRef] [Green Version]
- Chuah, L.-O.; Foo, H.L.; Loh, T.C.; Alitheen, N.B.M.; Yeap, S.K.; Mutalib, N.E.A.; Rahim, R.A.; Yusoff, K. Postbiotic metabolites produced by Lactobacillus plantarum strains exert selective cytotoxicity effects on cancer cells. BMC Complement. Altern. Med. 2019, 19, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Mack, D.R.; Michail, S.; Wei, S.; McDougall, L.; Hollingsworth, M.A. Probiotics inhibit enteropathogenic E. coli adherence in vitro by inducing intestinal mucin gene expression. Am. J. Physiol. Liver Physiol. 1999, 276, G941–G950. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, Z.; Qiu, L.; Zhang, F.; Xu, X.; Wei, H.; Tao, X. Characterization and bioactivities of the exopolysaccharide from a probiotic strain of Lactobacillus plantarum WLPL04. J. Dairy Sci. 2017, 100, 6895–6905. [Google Scholar] [CrossRef]
- Sionek, B.; Kołożyn-Krajewska, D. Bezpieczeństwo stosowania probiotyków przez ludzi. Zywnosc Nauk. Technol. Jakosc/Food Sci. Technol. Qual. 2019, 120, 5–21. [Google Scholar] [CrossRef]
- Tsilingiri, K.; Barbosa, T.; Penna, G.; Caprioli, F.; Sonzogni, A.M.; Viale, G.; Rescigno, M. Probiotic and postbiotic activity in health and disease: Comparison on a novel polarised ex-vivo organ culture model. Gut 2012, 61, 1007–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, G.; Korenaga, H.; Nagamine, R.; Takayama, H.; Kawahara, S.; Takeda, S.; Kikuchi, Y.; Dashnyam, B.; Kono, T.; Sakai, M. Cytokine responses in the Japanese pufferfish (Takifugu rubripes) head kidney cells induced with heat-killed probiotics isolated from the Mongolian dairy products. Fish Shellfish. Immunol. 2013, 34, 1170–1177. [Google Scholar] [CrossRef] [PubMed]
- Koscik, R.J.E.; Reid, G.; Kim, S.O.; Li, W.; Challis, J.R.G.; Bocking, A.D. Effect of Lactobacillus rhamnosus GR-I Supernatant on Cytokine and Chemokine Output From Human Amnion Cells Treated With Lipoteichoic Acid and Lipopolysaccharide. Reprod. Sci. 2018, 25, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Kolling, Y.; Salva, S.; Villena, J.; Alvarez, S. Are the immunomodulatory properties of Lactobacillus rhamnosus CRL1505 peptidoglycan common for all Lactobacilli during respiratory infection in malnourished mice? PLoS ONE 2018, 13, e0194034. [Google Scholar] [CrossRef] [Green Version]
- Sang, L.-X.; Chang, B.; Dai, C.; Gao, N.; Liu, W.-X.; Jiang, M. Heat-killed VSL#3 Ameliorates Dextran Sulfate Sodium (DSS)-Induced Acute Experimental Colitis in Rats. Int. J. Mol. Sci. 2013, 15, 15–28. [Google Scholar] [CrossRef]
- Ueno, N.; Fujiya, M.; Segawa, S.; Nata, T.; Moriichi, K.; Tanabe, H.; Mizukami, Y.; Kobayashi, N.; Ito, K.; Kohgo, Y. Heat-killed body of lactobacillus brevis SBC8803 ameliorates intestinal injury in a murine model of colitis by enhancing the intestinal barrier function. Inflamm. Bowel Dis. 2011, 17, 2235–2250. [Google Scholar] [CrossRef]
- Merghni, A.; Dallel, I.; Noumi, E.; Kadmi, Y.; Hentati, H.; Tobji, S.; Ben Amor, A.; Mastouri, M. Antioxidant and antiproliferative potential of biosurfactants isolated from Lactobacillus casei and their anti-biofilm effect in oral Staphylococcus aureus strains. Microb. Pathog. 2017, 104, 84–89. [Google Scholar] [CrossRef]
- Gur, J.; Mawuntu, M.; Martirosyan, D. FFC’s Advancement of Functional Food Definition. Funct. Foods Heal. Dis. 2018, 8, 385. [Google Scholar] [CrossRef]
- Kaur, S.; Das, M. Functional foods: An overview. Food Sci. Biotechnol. 2011, 20, 861–875. [Google Scholar] [CrossRef]
- Markowiak, P.; Śliżewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef]
- Karbowiak, M.; Zielińska, D. Postbiotyki–właściwości, zastosowanie i wpływ na zdrowie człowieka. Zywnosc Nauk. Technol. Jakosc/Food Sci. Technol. Qual. 2020, 123, 22–37. [Google Scholar] [CrossRef]
- Williams, N.T. Probiotics. Am. J. Health Syst. Pharm. 2010, 67, 449–458. [Google Scholar] [CrossRef]
- Fenster, K.; Freeburg, B.; Hollard, C.; Wong, C.; Laursen, R.R.; Ouwehand, A.C. The Production and Delivery of Probiotics: A Review of a Practical Approach. Microorganisms 2019, 7, 83. [Google Scholar] [CrossRef] [Green Version]
- Imperial, I.C.V.J.; Ibana, J.A. Addressing the Antibiotic Resistance Problem with Probiotics: Reducing the Risk of Its Double-Edged Sword Effect. Front. Microbiol. 2016, 07, 1983. [Google Scholar] [CrossRef] [Green Version]
- Johnson, C.N.; Kogut, M.H.; Genovese, K.; He, H.; Kazemi, S.; Arsenault, R.J. Administration of a Postbiotic Causes Immunomodulatory Responses in Broiler Gut and Reduces Disease Pathogenesis Following Challenge. Microorganisms 2019, 7, 268. [Google Scholar] [CrossRef] [Green Version]
- Flach, J.; van der Waal, M.; Nieuwboer, M.V.D.; Claassen, H.; Larsen, O.F.A. The underexposed role of food matrices in probiotic products: Reviewing the relationship between carrier matrices and product parameters. Crit. Rev. Food Sci. Nutr. 2017, 58, 2570–2584. [Google Scholar] [CrossRef] [Green Version]
- Wan, M.L.Y.; Forsythe, S.J.; El-Nezami, H. Probiotics interaction with foodborne pathogens: A potential alternative to antibiotics and future challenges. Crit. Rev. Food Sci. Nutr. 2018, 59, 3320–3333. [Google Scholar] [CrossRef]
- Sarao, L.K.; Arora, M. Probiotics, prebiotics, and microencapsulation: A review. Crit. Rev. Food Sci. Nutr. 2015, 57, 344–371. [Google Scholar] [CrossRef]
- Tripathy, S.; Verma, D.K.; Thakur, M.; Patel, A.R.; Srivastav, P.P.; Singh, S.; Chávez-González, M.L.; Aguilar, C.N. Encapsulated Food Products as a Strategy to Strengthen Immunity Against COVID-19. Front. Nutr. 2021, 8, 673174. [Google Scholar] [CrossRef]
- Atia, A.; Gomma, A.I.; Fliss, I.; Beyssac, E.; Garrait, G.; Subirade, M. Molecular and biopharmaceutical investigation of alginate–inulin synbiotic coencapsulation of probiotic to target the colon. J. Microencapsul. 2017, 34, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, T.; Song, Y.; Shu, G.; Chen, H. Effect of xanthan-chitosan-xanthan double layer encapsulation on survival of Bifidobacterium BB01 in simulated gastrointestinal conditions, bile salt solution and yogurt. Lwt 2017, 81, 274–280. [Google Scholar] [CrossRef]
- Vishwakarma, S.; Panigrahi, C.; Barua, S.; Sahoo, M.; Mandliya, S. Food nutrients as inherent sources of immunomodulation during COVID-19 pandemic. Lwt 2022, 158, 113154. [Google Scholar] [CrossRef] [PubMed]
- Rodrã guez-Gã³Mez, F.; Romero-Gil, V.; Garcã a-Garcã a, P.; Fernández, A.G.; Arroyo-Lã³Pez, F.N. Fortification of table olive packing with the potential probiotic bacteria Lactobacillus pentosus TOMC-LAB2. Front. Microbiol. 2014, 5, 467. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, T.C.; da Costa, W.K.A.; Barão, C.E.; Rosset, M.; Magnani, M. Vegan probiotic products: A modern tendency or the newest challenge in functional foods. Food Res. Int. 2020, 140, 110033. [Google Scholar] [CrossRef] [PubMed]
- Zoumpopoulou, G.; Tzouvanou, A.; Mavrogonatou, E.; Alexandraki, V.; Georgalaki, M.; Anastasiou, R.; Papadelli, M.; Manolopoulou, E.; Kazou, M.; Kletsas, D.; et al. Probiotic Features of Lactic Acid Bacteria Isolated from a Diverse Pool of Traditional Greek Dairy Products Regarding Specific Strain-Host Interactions. Probiotics Antimicrob. Proteins 2017, 10, 313–322. [Google Scholar] [CrossRef]
- Bengoa, A.A.; Zavala, L.; Carasi, P.; Trejo, S.; Bronsoms, S.; Serradell, M.D.L.A.; Garrote, G.L.; Abraham, A.G. Simulated gastrointestinal conditions increase adhesion ability of Lactobacillus paracasei strains isolated from kefir to Caco-2 cells and mucin. Food Res. Int. 2018, 103, 462–467. [Google Scholar] [CrossRef]
- Caggia, C.; De Angelis, M.; Pitino, I.; Pino, A.; Randazzo, C. Probiotic features of Lactobacillus strains isolated from Ragusano and Pecorino Siciliano cheeses. Food Microbiol. 2015, 50, 109–117. [Google Scholar] [CrossRef]
- Xing, Z.; Tang, W.; Geng, W.; Zheng, Y.; Wang, Y. In vitro and in vivo evaluation of the probiotic attributes of Lactobacillus kefiranofaciens XL10 isolated from Tibetan kefir grain. Appl. Microbiol. Biotechnol. 2016, 101, 2467–2477. [Google Scholar] [CrossRef]
- Zielińska, D.; Kolożyn-Krajewska, D. Food-Origin Lactic Acid Bacteria May Exhibit Probiotic Properties: Review. BioMed Res. Int. 2018, 2018, 5063185. [Google Scholar] [CrossRef] [Green Version]
- Salvador, P.B.U.; Dalmacio, L.M.M.; Kim, S.H.; Kang, D.-K.; Balolong, M.P. Immunomodulatory potential of four candidate probiotic Lactobacillus strains from plant and animal origin using comparative genomic analysis. Access Microbiol. 2021, 3, 000299. [Google Scholar] [CrossRef]
- Chaluvadi, A.T.; Hotchkiss, K.L. Yam. In Gut Microbiota: Impact Of Probiotics, Prebiotics, Synbiotics, Pharmabiotics, A Postbi-Otics On Human; Health, R.R., Watson, V.R.P., Eds.; Probiotics, prebiotics and synbiotics. Bioactive foods in health pro-motion; Elsevier: London, UK, 2016; pp. 515–523. [Google Scholar]
- Hosseini, S.A.; Abbasi, A.; Sabahi, S.; Khani, N. Application of Postbiotics Produced By Lactic Acid Bacteria in the Development of Active Food Packaging. Biointerface Res. Appl. Chem. 2021, 12, 6164–6183. [Google Scholar] [CrossRef]
- Morniroli, D.; Vizzari, G.; Consales, A.; Mosca, F.; Giannì, M. Postbiotic Supplementation for Children and Newborn’s Health. Nutrients 2021, 13, 781. [Google Scholar] [CrossRef]
- Garnier, L.; Mounier, J.; Lê, S.; Pawtowski, A.; Pinon, N.; Camier, B.; Chatel, M.; Garric, G.; Thierry, A.; Coton, E.; et al. Development of antifungal ingredients for dairy products: From in vitro screening to pilot scale application. Food Microbiol. 2018, 81, 97–107. [Google Scholar] [CrossRef]
- Darwish, M.S.; Qiu, L.; Taher, M.A.; Zaki, A.A.; Abou-Zeid, N.A.; Dawood, D.H.; Shalabi, O.M.A.K.; Khojah, E.; Elawady, A.A. Health Benefits of Postbiotics Produced by E. coli Nissle 1917 in Functional Yogurt Enriched with Cape Gooseberry (Physalis peruviana L.). Fermentation 2022, 8, 128. [Google Scholar] [CrossRef]
- Ramos, I.M.; Rodríguez-Sánchez, S.; Seseña, S.; Palop, M.L.; Poveda, J.M. Assessment of safety characteristics, postbiotic potential, and technological stress response of Leuconostoc strains from different origins for their use in the production of functional dairy foods. LWT 2022, 165, 113722. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Category | Food Components | Research Effect | Reference |
|---|---|---|---|
| Immunobiotics | |||
| Lactobacillus | L. delbrueckii ssp. bulgaricus OLL1073R-1 (1073R-1-yogurt) | Consumption of the product affected influenza A virus subtype H3N2-bound Immunoglobulin A (IgA) levels in saliva. | Yamato et al., 2019 [44] |
| L. paracasei N1115 | The intake of yogurt containing L. paracasei could protect against the risk of acute upper respiratory tract infection in the middle-aged and elderly; it might be that L. paracasei stimulated T-cell immunity | Pu et al., 2017 [45] | |
| L. plantarum NRIC1832 L. plantarum NRIC0380 | Inhibition of allergy, induction of regulatory T cells by enhancement of IL-10 production, RALDH activity | Noguchi et al., 2012 [46] Yoshida et al., 2013 [47] | |
| L. rhamnosus CRL1505 L. plantarum CRL1506 | Antiviral factors and cytokines/chemokines were increased in lactobacilli-treated PIE cells. The expression of the IL-15 and RAE1 genes that mediate poly (I:C) inflammatory damage was also reduced | Albarracin et al., 2017 [48] | |
| L. gasseri OLL2809 | Induction of regulatory T cells | Aoki-Yoshida et. al., 2016 [49] | |
| Bifidobacterium | B. longum MCC1, B. infantis MCC12, B. breve MCC16, B. pseudolongum MCC92, L. paracasei MCC1375, L. gasseri MCC587, and L. sub ssp. lactis MCC866 | B. infantis MCC12 and B. breve MCC1274 increased the production of INF-β in PIE cells, in response to VR infection. They also increased the expression of CXCL10 and IL-6 genes, especially the B. infantis | Ishizuka et al., 2016 [50] |
| Saccharomyces | S. cerevisiae var. boulardii | Formation of glutathione, which is responsible for the stimulation of the activity of immune cells. | Badr et al., 2021 [51] |
| Category Probiotic Microorganism | Postbiotic Components | Research Effect | Reference |
|---|---|---|---|
| L. paracasei B21060 | Cell-free supernatants | Anti-inflammatory effect | Tsilingiri et al., 2012 [110] |
| L. paracasei spp. paracasei 06TCa22 and L. plantarum 06CC2 | Heat-killed cells | Immunomodulation effect | Biswas et al., 2013 [111] |
| L. rhamnosus GR-1 | Cell-free supernatants | Immunomodulatory activity | Kościk et al., 2018 [112] |
| L. rhamnosus CRL1505 | Peptidoglycan | Improved of Th2 response | Kolling et al., 2018 [113] |
| VSL#3 (L. plantarum, L. bulgaricus, L. casei and L. acidophilus; S. salivarius subsp. thermophilus) | Heat-killed cells | Anti-inflammatory | Sang et al., 2014 [114] |
| L. brevis SBC8803, L. brevis 8013 B. longum, and Streptococcus faecalis | Heat-killed cells | Anti-inflammatory and Enhancement of epithelial barrier permeability | Ueno et al., 2011 [115] |
| L. casei B1 | Biosurfactants | Anti-proliferative, anti-oxidative, and anti-adhesion activity against S. aureus | Merghini et al., 2017 [116] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szydłowska, A.; Sionek, B. Probiotics and Postbiotics as the Functional Food Components Affecting the Immune Response. Microorganisms 2023, 11, 104. https://doi.org/10.3390/microorganisms11010104
Szydłowska A, Sionek B. Probiotics and Postbiotics as the Functional Food Components Affecting the Immune Response. Microorganisms. 2023; 11(1):104. https://doi.org/10.3390/microorganisms11010104
Chicago/Turabian StyleSzydłowska, Aleksandra, and Barbara Sionek. 2023. "Probiotics and Postbiotics as the Functional Food Components Affecting the Immune Response" Microorganisms 11, no. 1: 104. https://doi.org/10.3390/microorganisms11010104