
The Post-Transcriptional Regulatory Protein CsrA Amplifies Its Targetome through Direct Interactions with Stress-Response Regulatory Hubs: The EvgA and AcnA Cases


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plasmids and Strains
2.2. In Vitro Electrophoretic Mobility Shift Assays
2.3. In Vivo Fluorescence Reporter Assays
2.4. In Vitro Coupled Transcription-Translation Assays
2.5. RNA Isolation and Transcriptomics Analysis
2.6. Network Reconstruction and Analysis
3. Results
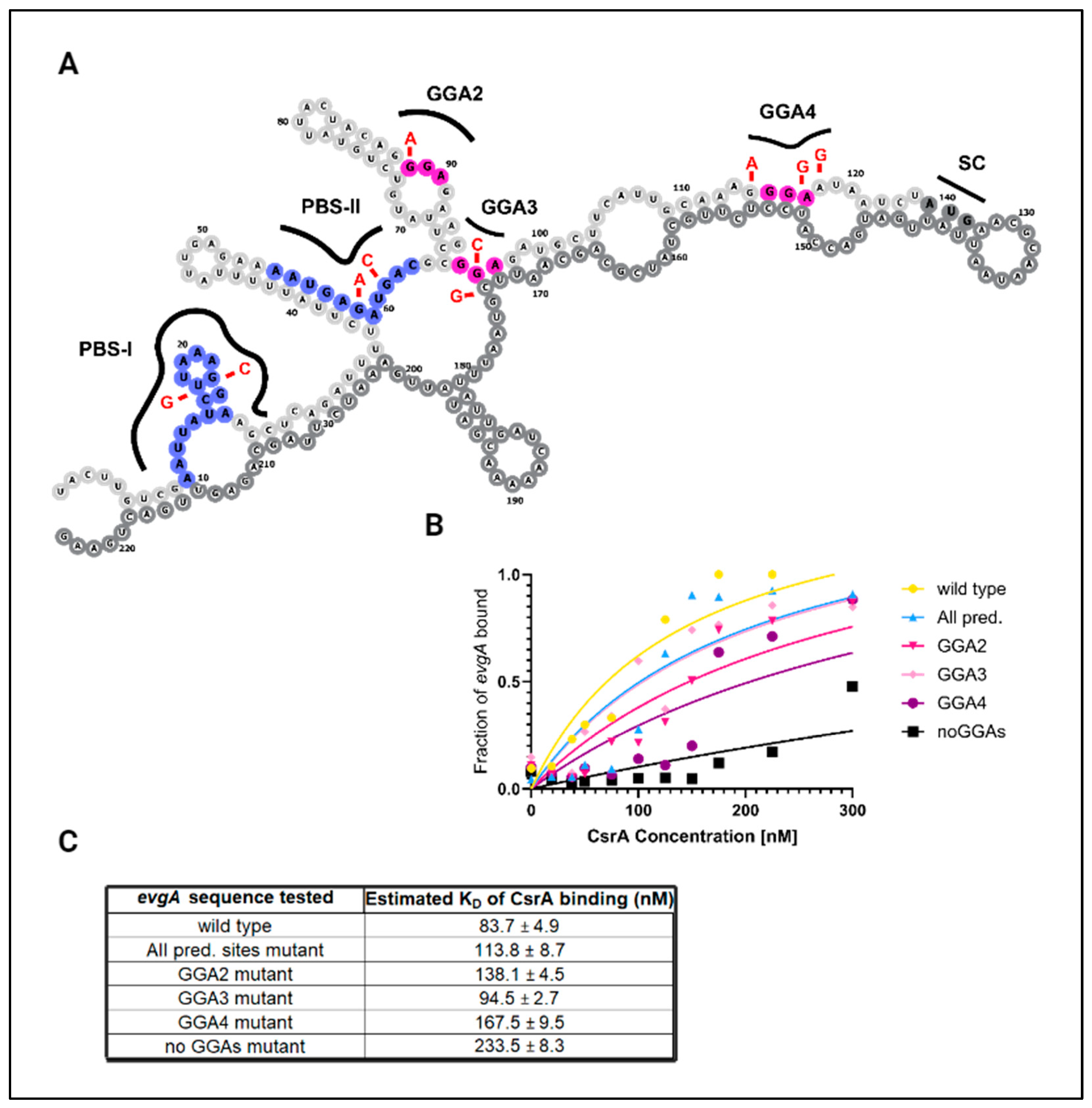
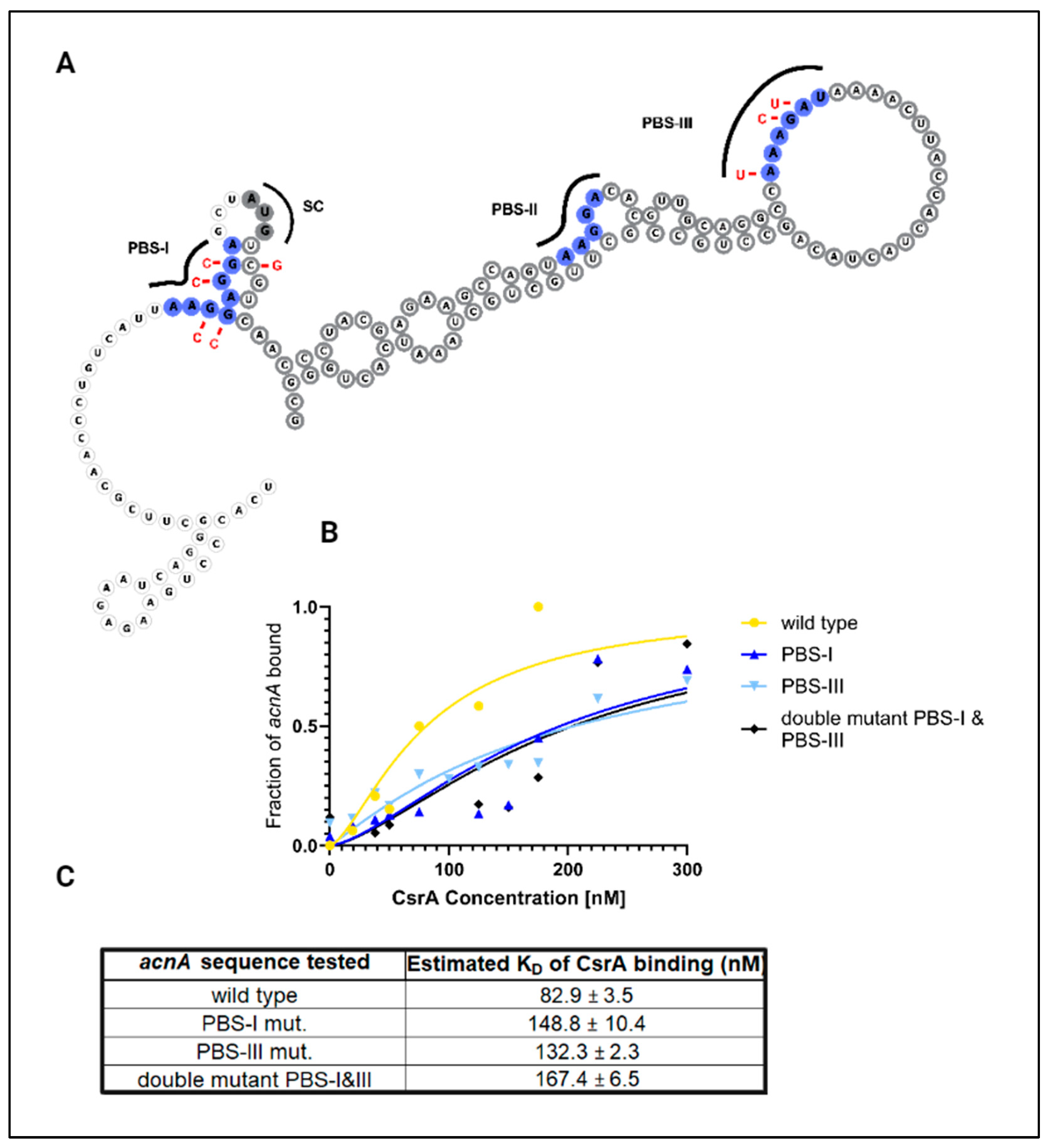
3.1. CsrA Binds Directly to the Leader Sequences of the evgA and acnA Transcripts Encoding Regulatory Proteins
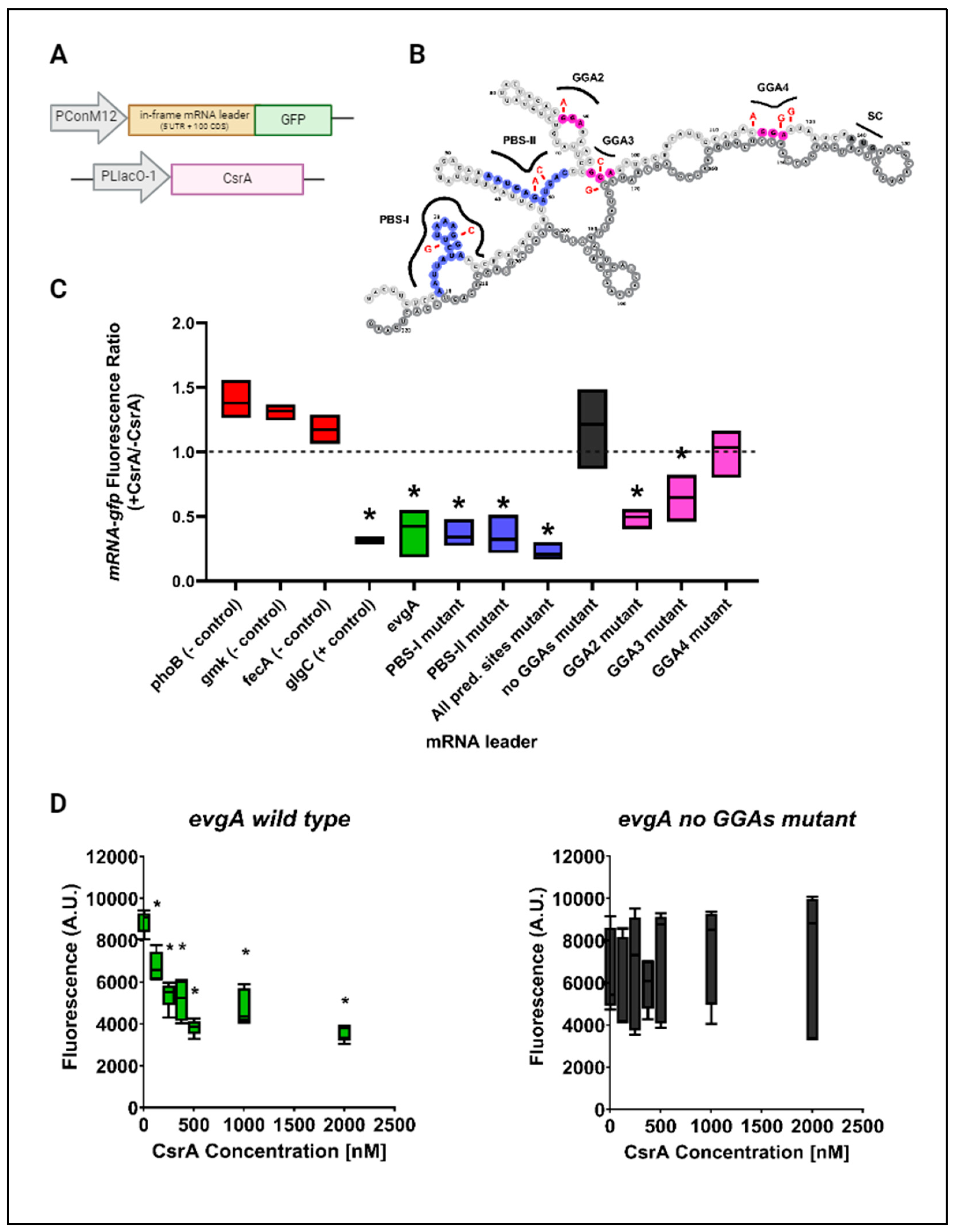
3.2. CsrA Represses Both evgA and acnA In Vivo and In Vitro
3.3. CsrA Influences Distinct Stress Phenotypic Clusters through Its Interactions with evgA and acnA
3.4. The CsrA-evgA Interaction Is Important for Acidic Stress Tolerance in E. coli
3.5. CsrA Influences Metal Ion Tolerance in E. coli through Its Interaction with acnA
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Romeo, T.; Gong, M.; Liu, M.Y.; Brun-Zinkernagel, A.M. Identification and molecular characterization of csrA, a pleiotropic gene from Escherichia coli that affects glycogen biosynthesis, gluconeogenesis, cell size, and surface properties. J. Bacteriol. 1993, 175, 4744–4755. [Google Scholar] [CrossRef] [PubMed]
- Potts, A.H.; Vakulskas, C.A.; Pannuri, A.; Yakhnin, H.; Babitzke, P.; Romeo, T. Global role of the bacterial post-transcriptional regulator CsrA revealed by integrated transcriptomics. Nat. Commun. 2017, 8, 1596. [Google Scholar] [CrossRef] [PubMed]
- Pourciau, C.; Pannuri, A.; Potts, A.; Yakhnin, H.; Babitzke, P.; Romeo, T. Regulation of Iron Storage by CsrA Supports Exponential Growth of Escherichia coli. mBio 2019, 10, e01034-19. [Google Scholar] [CrossRef] [PubMed]
- Rojano-Nisimura, A.M.; Simmons, T.R.; Leistra, A.N.; Mihailovic, M.K.; Buchser, R.; Ekdahl, A.M.; Joseph, I.; Curtis, N.C.; Contreras, L.M. CsrA selectively modules sRNA-mRNA regulatory outcomes. Front. Mol. Biosci. 2023, 2023, 1249528. [Google Scholar] [CrossRef]
- Dubey, A.K.; Baker, C.S.; Romeo, T.; Babitzke, P. RNA sequence and secondary structure participate in high-affinity CsrA-RNA interaction. RNA 2005, 11, 1579–1587. [Google Scholar] [CrossRef]
- Baker, C.S.; Morozov, I.; Suzuki, K.; Romeo, T.; Babitzke, P. CsrA regulates glycogen biosynthesis by preventing translation of glgC in Escherichia coli. Mol. Microbiol. 2002, 44, 1599–1610. [Google Scholar] [CrossRef]
- Dubey, A.K.; Baker, C.S.; Suzuki, K.; Jones, A.D.; Pandit, P.; Romeo, T.; Babitzke, P. CsrA regulates translation of the Escherichia coli carbon starvation gene, cstA, by blocking ribosome access to the cstA transcript. J. Bacteriol. 2003, 185, 4450–4460. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Dubey, A.K.; Suzuki, K.; Baker, C.S.; Babitzke, P.; Romeo, T. CsrA post-transcriptionally represses pgaABCD, responsible for synthesis of a biofilm polysaccharide adhesin of Escherichia coli. Mol. Microbiol. 2005, 56, 1648–1663. [Google Scholar] [CrossRef]
- Baker, C.S.; Eöry, L.A.; Yakhnin, H.; Mercante, J.; Romeo, T.; Babitzke, P. CsrA inhibits translation initiation of Escherichia coli hfq by binding to a single site overlapping the Shine-Dalgarno sequence. J. Bacteriol. 2007, 189, 5472–5481. [Google Scholar] [CrossRef]
- Jonas, K.; Edwards, A.N.; Simm, R.; Romeo, T.; Römling, U.; Melefors, O. The RNA binding protein CsrA controls cyclic di-GMP metabolism by directly regulating the expression of GGDEF proteins. Mol. Microbiol. 2008, 70, 236–257. [Google Scholar] [CrossRef]
- Edwards, A.N.; Patterson-Fortin, L.M.; Vakulskas, C.A.; Mercante, J.W.; Potrykus, K.; Vinella, D.; Camacho, M.I.; Fields, J.A.; Thompson, S.A.; Georgellis, D.; et al. Circuitry linking the Csr and stringent response global regulatory systems. Mol. Microbiol. 2011, 80, 1561–1580. [Google Scholar] [CrossRef]
- Yakhnin, H.; Baker, C.S.; Berezin, I.; Evangelista, M.A.; Rassin, A.; Romeo, T.; Babitzke, P. CsrA represses translation of sdiA, which encodes the N-acylhomoserine-L-lactone receptor of Escherichia coli, by binding exclusively within the coding region of sdiA mRNA. J. Bacteriol. 2011, 193, 6162–6170. [Google Scholar] [CrossRef]
- Yakhnin, H.; Yakhnin, A.V.; Baker, C.S.; Sineva, E.; Berezin, I.; Romeo, T.; Babitzke, P. Complex regulation of the global regulatory gene csrA: CsrA-mediated translational repression, transcription from five promoters by Eσ⁷⁰ and Eσ(S), and indirect transcriptional activation by CsrA. Mol. Microbiol. 2011, 81, 689–704. [Google Scholar] [CrossRef]
- Pannuri, A.; Yakhnin, H.; Vakulskas, C.A.; Edwards, A.N.; Babitzke, P.; Romeo, T. Translational repression of NhaR, a novel pathway for multi-tier regulation of biofilm circuitry by CsrA. J. Bacteriol. 2012, 194, 79–89. [Google Scholar] [CrossRef]
- Jørgensen, M.G.; Thomason, M.K.; Havelund, J.; Valentin-Hansen, P.; Storz, G. Dual function of the McaS small RNA in controlling biofilm formation. Genes Dev. 2013, 27, 1132–1145. [Google Scholar] [CrossRef] [PubMed]
- Yakhnin, A.V.; Baker, C.S.; Vakulskas, C.A.; Yakhnin, H.; Berezin, I.; Romeo, T.; Babitzke, P. CsrA activates flhDC expression by protecting flhDC mRNA from RNase E-mediated cleavage. Mol. Microbiol. 2013, 87, 851–866. [Google Scholar] [CrossRef]
- Park, H.; McGibbon, L.C.; Potts, A.H.; Yakhnin, H.; Romeo, T.; Babitzke, P. Translational Repression of the RpoS Antiadapter IraD by CsrA Is Mediated via Translational Coupling to a Short Upstream Open Reading Frame. mBio 2017, 8, e01355-17. [Google Scholar] [CrossRef] [PubMed]
- Yakhnin, H.; Aichele, R.; Ades, S.E.; Romeo, T.; Babitzke, P. Circuitry Linking the Global Csr- and σE-Dependent Cell Envelope Stress Response Systems. J. Bacteriol. 2017, 199, e00484-17. [Google Scholar] [CrossRef] [PubMed]
- Renda, A.; Poly, S.; Lai, Y.-J.; Pannuri, A.; Yakhnin, H.; Potts, A.H.; Bevilacqua, P.C.; Romeo, T.; Babitzke, P. CsrA-Mediated Translational Activation of ymdA Expression in Escherichia coli. mBio 2020, 11, e00849-20. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.J.; Yakhnin, H.; Pannuri, A.; Pourciau, C.; Babitzke, P.; Romeo, T. CsrA regulation via binding to the base-pairing small RNA Spot 42. Mol. Microbiol. 2022, 117, 32–53. [Google Scholar] [CrossRef] [PubMed]
- Pourciau, C.; Yakhnin, H.; Pannuri, A.; Gorelik, M.G.; Lai, Y.-J.; Romeo, T.; Babitzke, P. CsrA coordinates the expression of ribosome hibernation and anti-σ factor proteins. mBio 2023, 14, e0258523. [Google Scholar] [CrossRef] [PubMed]
- Sowa, S.W.; Gelderman, G.; Leistra, A.N.; Buvanendiran, A.; Lipp, S.; Pitaktong, A.; Vakulskas, C.A.; Romeo, T.; Baldea, M.; Contreras, L.M. Integrative FourD omics approach profiles the target network of the carbon storage regulatory system. Nucleic Acids Res. 2017, 45, 1673–1686. [Google Scholar] [CrossRef] [PubMed]
- Leistra, A.N.; Gelderman, G.; Sowa, S.W.; Moon-Walker, A.; Salis, H.M.; Contreras, L.M. A Canonical Biophysical Model of the CsrA Global Regulator Suggests Flexible Regulator-Target Interactions. Sci. Rep. 2018, 8, 9892. [Google Scholar] [CrossRef]
- McKee, A.E.; Rutherford, B.J.; Chivian, D.C.; Baidoo, E.K.; Juminaga, D.; Kuo, D.; Benke, P.I.; Dietrich, J.A.; Ma, S.M.; Arkin, A.P.; et al. Manipulation of the carbon storage regulator system for metabolite remodeling and biofuel production in Escherichia coli. Microb. Cell Fact. 2012, 11, 79. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, P.R.; Jia, T.; Kuehne, S.A.; Kerkering, T.M.; Morris, E.R.; Searle, M.S.; Heeb, S.; Rao, J.; Kulkarni, R.V. A sequence-based approach for prediction of CsrA/RsmA targets in bacteria with experimental validation in Pseudomonas aeruginosa. Nucleic Acids Res. 2014, 42, 6811–6825. [Google Scholar] [CrossRef]
- Esquerré, T.; Bouvier, M.; Turlan, C.; Carpousis, A.J.; Girbal, L.; Cocaign-Bousquet, M. The Csr system regulates genome-wide mRNA stability and transcription and thus gene expression in Escherichia coli. Sci. Rep. 2016, 6, 25057. [Google Scholar] [CrossRef]
- Romeo, T.; Babitzke, P. Global Regulation by CsrA and Its RNA Antagonists. Microbiol. Spectr. 2018, 6, 341–354. [Google Scholar] [CrossRef]
- Gerovac, M.; El Mouali, Y.; Kuper, J.; Kisker, C.; Barquist, L.; Vogel, J. Global discovery of bacterial RNA-binding proteins by RNase-sensitive gradient profiles reports a new FinO domain protein. RNA 2020, 26, 1448–1463. [Google Scholar] [CrossRef]
- Katsuya-Gaviria, K.; Paris, G.; Dendooven, T.; Bandyra, K.J. Bacterial RNA chaperones and chaperone-like riboregulators: Behind the scenes of RNA-mediated regulation of cellular metabolism. RNA Biol. 2022, 19, 419–436. [Google Scholar] [CrossRef]
- Eguchi, Y.; Utsumi, R. Alkali metals in addition to acidic pH activate the EvgS histidine kinase sensor in Escherichia coli. J. Bacteriol. 2014, 196, 3140–3149. [Google Scholar] [CrossRef]
- Nishino, K.; Inazumi, Y.; Yamaguchi, A. Global analysis of genes regulated by EvgA of the two-component regulatory system in Escherichia coli. J. Bacteriol. 2003, 185, 2667–2672. [Google Scholar] [CrossRef]
- Cunningham, L.; Gruer, M.J.; Guest, J.R. Transcriptional regulation of the aconitase genes (acnA and acnB) of Escherichia coli. Microbiology 1997, 143 Pt 12, 3795–3805. [Google Scholar] [CrossRef]
- Tang, Y.; Guest, J.R. Direct evidence for mRNA binding and post-transcriptional regulation by Escherichia coli aconitases. Microbiology 1999, 145 Pt 11, 3069–3079. [Google Scholar] [CrossRef]
- Tang, Y.; Quail, M.A.; Artymiuk, P.J.; Guest, J.R.; Green, J. Escherichia coli aconitases and oxidative stress: Post-transcriptional regulation of sodA expression. Microbiology 2002, 148 Pt 4, 1027–1037. [Google Scholar] [CrossRef]
- Mehrer, C.R.; Incha, M.R.; Politz, M.C.; Pfleger, B.F. Anaerobic production of medium-chain fatty alcohols via a β-reduction pathway. Metab. Eng. 2018, 48, 63–71. [Google Scholar] [CrossRef]
- Yakhnin, A.V.; Yakhnin, H.; Babitzke, P. Gel mobility shift assays to detect protein-RNA interactions. Methods Mol. Biol. 2012, 905, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Adamson, D.N.; Lim, H.N. Rapid and robust signaling in the CsrA cascade via RNA-protein interactions and feedback regulation. Proc. Natl. Acad. Sci. USA 2013, 110, 13120–13125. [Google Scholar] [CrossRef] [PubMed]
- Leistra, A.N.; Amador, P.; Buvanendiran, A.; Moon-Walker, A.; Contreras, L.M. Rational Modular RNA Engineering Based on In Vivo Profiling of Structural Accessibility. ACS Synth. Biol. 2017, 6, 2228–2240. [Google Scholar] [CrossRef] [PubMed]
- Lukasiewicz, A.J.; Contreras, L.M. Antisense probing of dynamic RNA structures. Methods 2020, 183, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Sweet, P.; Blacutt, J.; Gordon, V.; Contreras, L. Exposure of Shewanella oneidensis MR-1 to Sublethal Doses of Ionizing Radiation Triggers Short-Term SOS Activation and Longer-Term Prophage Activation. Appl. Environ. Microbiol. 2023, 89, e0171622. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- De Biase, D.; Lund, P.A. The Escherichia coli Acid Stress Response and Its Significance for Pathogenesis. Adv. Appl. Microbiol. 2015, 92, 49–88. [Google Scholar] [CrossRef]
- Nishino, K.; Yamaguchi, A. EvgA of the two-component signal transduction system modulates production of the yhiUV multidrug transporter in Escherichia coli. J. Bacteriol. 2002, 184, 2319–2323. [Google Scholar] [CrossRef]
- Gelderman, G.; Sivakumar, A.; Lipp, S.; Contreras, L. Adaptation of Tri-molecular fluorescence complementation allows assaying of regulatory Csr RNA-protein interactions in bacteria. Biotechnol. Bioeng. 2015, 112, 365–375. [Google Scholar] [CrossRef]
- Leistra, A.N.; Mihailovic, M.K.; Contreras, L.M. Fluorescence-Based Methods for Characterizing RNA Interactions In Vivo. Methods Mol. Biol. 2018, 1737, 129–164. [Google Scholar] [CrossRef]
- Gorelik, M.G.; Yakhnin, H.; Pannuri, A.; Walker, A.C.; Pourciau, C.; Czyz, D.; Romeo, T.; Babitzke, P. Multitier regulation of the E. coli extreme acid stress response by CsrA. J. Bacteriol. 2024, e0035423. [Google Scholar] [CrossRef] [PubMed]
- Mercante, J.; Edwards, A.N.; Dubey, A.K.; Babitzke, P.; Romeo, T. Molecular geometry of CsrA (RsmA) binding to RNA and its implications for regulated expression. J. Mol. Biol. 2009, 392, 511–528. [Google Scholar] [CrossRef] [PubMed]
- Kammel, M.; Pinske, C.; Sawers, R.G. FocA and its central role in fine-tuning pH homeostasis of enterobacterial formate metabolism. Microbiology 2022, 168, 001253. [Google Scholar] [CrossRef] [PubMed]
- Rhee, H.J.; Kim, E.J.; Lee, J.K. Physiological polyamines: Simple primordial stress molecules. J. Cell. Mol. Med. 2007, 11, 685–703. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, M.K.; Tabor, H. Polyamines are critical for the induction of the glutamate decarboxylase-dependent acid resistance system in Escherichia coli. J. Biol. Chem. 2013, 288, 33559–33570. [Google Scholar] [CrossRef]
- Masuda, N.; Church, G.M. Regulatory network of acid resistance genes in Escherichia coli. Mol. Microbiol. 2003, 48, 699–712. [Google Scholar] [CrossRef]
- Gunasekera, T.S.; Csonka, L.N.; Paliy, O. Genome-wide transcriptional responses of Escherichia coli K-12 to continuous osmotic and heat stresses. J. Bacteriol. 2008, 190, 3712–3720. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Condemine, G.; Berrier, C.; Plumbridge, J.; Ghazi, A. Function and expression of an N-acetylneuraminic acid-inducible outer membrane channel in Escherichia coli. J. Bacteriol. 2005, 187, 1959–1965. [Google Scholar] [CrossRef][Green Version]
- Li, K.; Xin, Y.; Xuan, G.; Zhao, R.; Liu, H.; Xia, Y.; Xun, L. Escherichia coli Uses Separate Enzymes to Produce H2S and Reactive Sulfane Sulfur From L-cysteine. Front. Microbiol. 2019, 10, 298. [Google Scholar] [CrossRef]
- D’Aquila, P.; De Rango, F.; Paparazzo, E.; Passarino, G.; Bellizzi, D. Epigenetic-Based Regulation of Transcriptome in Escherichia coli Adaptive Antibiotic Resistance. Microbiol. Spectr. 2023, 11, e0458322. [Google Scholar] [CrossRef]
- Weber, A.; Kögl, S.A.; Jung, K. Time-dependent proteome alterations under osmotic stress during aerobic and anaerobic growth in Escherichia coli. J. Bacteriol. 2006, 188, 7165–7175. [Google Scholar] [CrossRef]
- Schneider, B.L.; Hernandez, V.J.; Reitzer, L. Putrescine catabolism is a metabolic response to several stresses in Escherichia coli. Mol. Microbiol. 2013, 88, 537–550. [Google Scholar] [CrossRef]
- Adam, J.P.; Claudia, T.; James, C.P. Streptococcus pneumoniae uses glutathione to defend against oxidative stress and metal ion toxicity. J. Bacteriol. 2012, 194, 6248–6254. [Google Scholar] [CrossRef]
- Watling, H.R.; Collinson, D.M.; Corbett, M.K.; Shiers, D.W.; Kaksonen, A.H.; Watkin, E.L. Saline-water bioleaching of chalcopyrite with thermophilic, iron(II)- and sulfur-oxidizing microorganisms. Res. Microbiol. 2016, 167, 546–554. [Google Scholar] [CrossRef]
- Masuda, N.; Church, G.M. Escherichia coli gene expression responsive to levels of the response regulator EvgA. J. Bacteriol. 2002, 184, 6225–6234. [Google Scholar] [CrossRef]
- Nepal, S.; Kumar, P. Growth, Cell Division, and Gene Expression of Escherichia coli at Elevated Concentrations of Magnesium Sulfate: Implications for Habitability of Europa and Mars. Microorganisms 2020, 8, 637. [Google Scholar] [CrossRef]
- Hobman, J.L.; Yamamoto, K.; Oshima, T. Transcriptomic responses of bacterial cells to sublethal metal ion stress. In Molecular Microbiology of Heavy Metals; Nies, D.H., Silver, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 73–115. [Google Scholar]
- Gelmi, M.; Apostoli, P.; Cabibbo, E.; Porru, S.; Alassio, L.; Turano, A. Resistance to cadmium salts and metal absorption by different microbial species. Curr. Microbiol. 1994, 29, 335–341. [Google Scholar] [CrossRef]
- Bouvier, J.; Gordia, S.; Kampmann, G.; Lange, R.; Hengge-Aronis, R.; Gutierrez, C. Interplay between global regulators of Escherichia coli: Effect of RpoS, Lrp and H-NS on transcription of the gene osmC. Mol. Microbiol. 1998, 28, 971–980. [Google Scholar] [CrossRef]
- Eguchi, Y.; Okada, T.; Minagawa, S.; Oshima, T.; Mori, H.; Yamamoto, K.; Ishihama, A.; Utsumi, R. Signal transduction cascade between EvgA/EvgS and PhoP/PhoQ two-component systems of Escherichia coli. J. Bacteriol. 2004, 186, 3006–3014. [Google Scholar] [CrossRef][Green Version]
- Isarankura-Na-Ayudhya, P.; Isarankura-Na-Ayudhya, C.; Treeratanapaiboon, L.; Kasikun, K.; Thipkeaw, K.; Prachayasittikul, V. Proteomic profiling of Escherichia coli in response to heavy metals stress. Eur. J. Sci. Res. 2009, 25, 679–688. [Google Scholar]
- Berry, B.J.; Jenkins, D.G.; Schuerger, A.C. Effects of simulated Mars conditions on the survival and growth of Escherichia coli and Serratia liquefaciens. Appl. Environ. Microbiol. 2010, 76, 2377–2386. [Google Scholar] [CrossRef]
- Cabral, D.J.; Penumutchu, S.; Norris, C.; Morones-Ramirez, J.R.; Belenky, P. Microbial competition between Escherichia coli and Candida albicans reveals a soluble fungicidal factor. Microb. Cell 2018, 5, 249–255. [Google Scholar] [CrossRef]
- Moreira Martins, P.M.; Gong, T.; de Souza, A.A.; Wood, T.K. Copper Kills Escherichia coli Persister Cells. Antibiotics 2020, 9, 506. [Google Scholar] [CrossRef] [PubMed]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rojano-Nisimura, A.M.; Grismore, K.B.; Ruzek, J.S.; Avila, J.L.; Contreras, L.M. The Post-Transcriptional Regulatory Protein CsrA Amplifies Its Targetome through Direct Interactions with Stress-Response Regulatory Hubs: The EvgA and AcnA Cases. Microorganisms 2024, 12, 636. https://doi.org/10.3390/microorganisms12040636
Rojano-Nisimura AM, Grismore KB, Ruzek JS, Avila JL, Contreras LM. The Post-Transcriptional Regulatory Protein CsrA Amplifies Its Targetome through Direct Interactions with Stress-Response Regulatory Hubs: The EvgA and AcnA Cases. Microorganisms. 2024; 12(4):636. https://doi.org/10.3390/microorganisms12040636
Chicago/Turabian StyleRojano-Nisimura, Alejandra Matsuri, Kobe B. Grismore, Josie S. Ruzek, Jacqueline L. Avila, and Lydia M. Contreras. 2024. "The Post-Transcriptional Regulatory Protein CsrA Amplifies Its Targetome through Direct Interactions with Stress-Response Regulatory Hubs: The EvgA and AcnA Cases" Microorganisms 12, no. 4: 636. https://doi.org/10.3390/microorganisms12040636
APA StyleRojano-Nisimura, A. M., Grismore, K. B., Ruzek, J. S., Avila, J. L., & Contreras, L. M. (2024). The Post-Transcriptional Regulatory Protein CsrA Amplifies Its Targetome through Direct Interactions with Stress-Response Regulatory Hubs: The EvgA and AcnA Cases. Microorganisms, 12(4), 636. https://doi.org/10.3390/microorganisms12040636