
Role of Staphylococcus aureus’s Buoyant Density in the Development of Biofilm Associated Antibiotic Susceptibility
 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
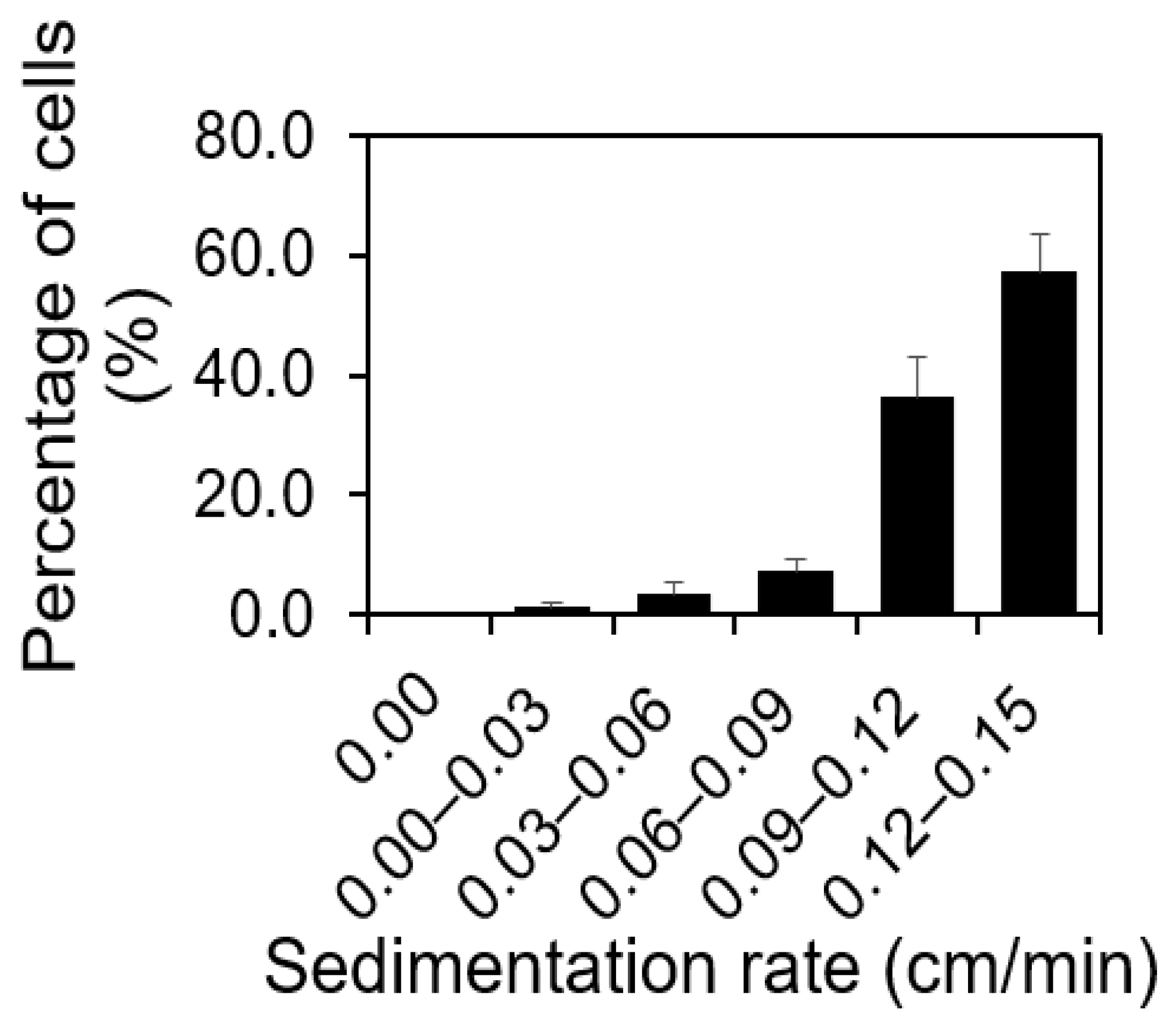
2.1. For Any Isogenic S. aureus Population, There Was a Distribution of Cell Buoyant Density, Regardless of the Population’s Age
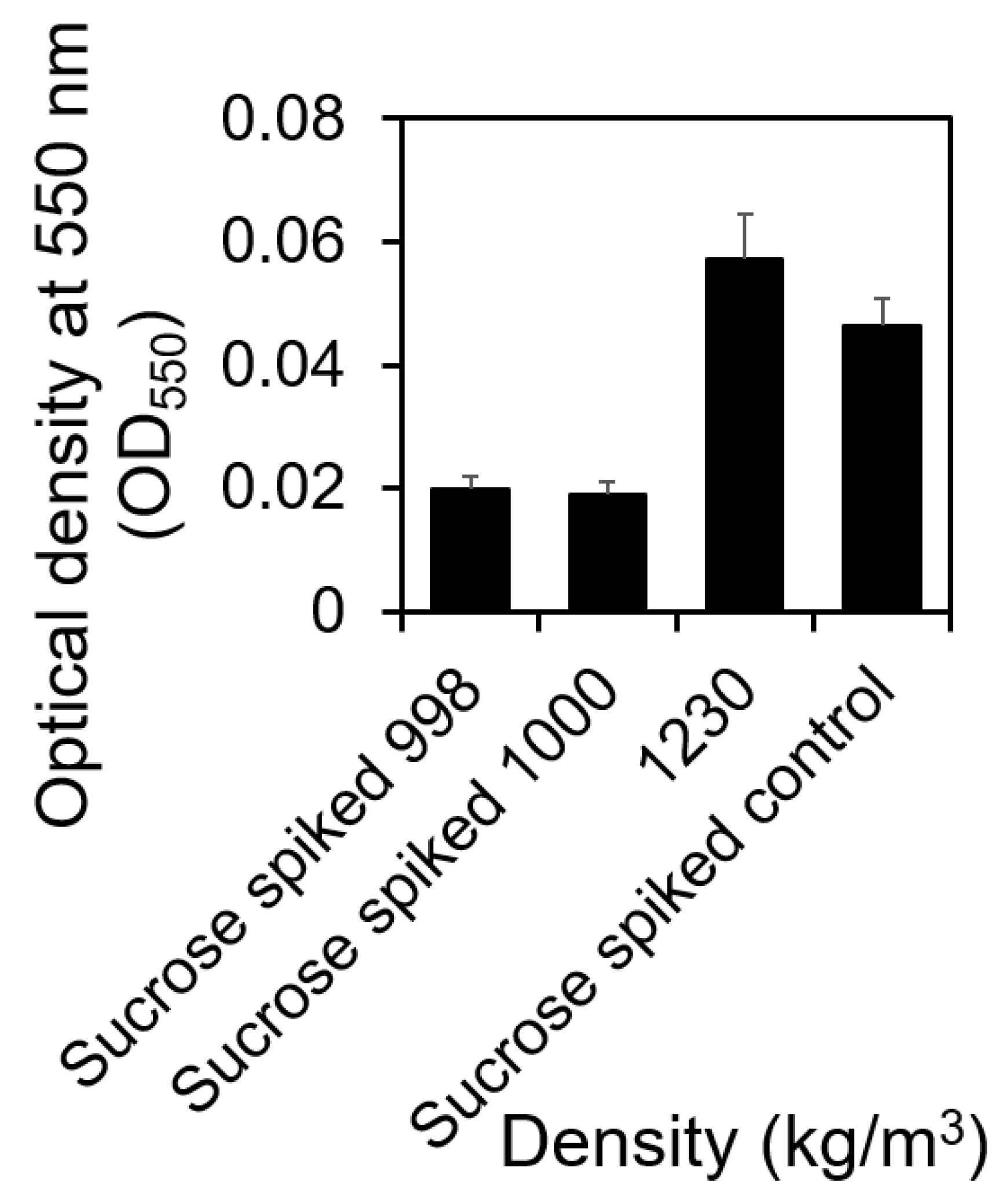
2.2. Correlating Sedimentation Rates with Cells’ Buoyant Density Using Sucrose Density Gradient Centrifugation
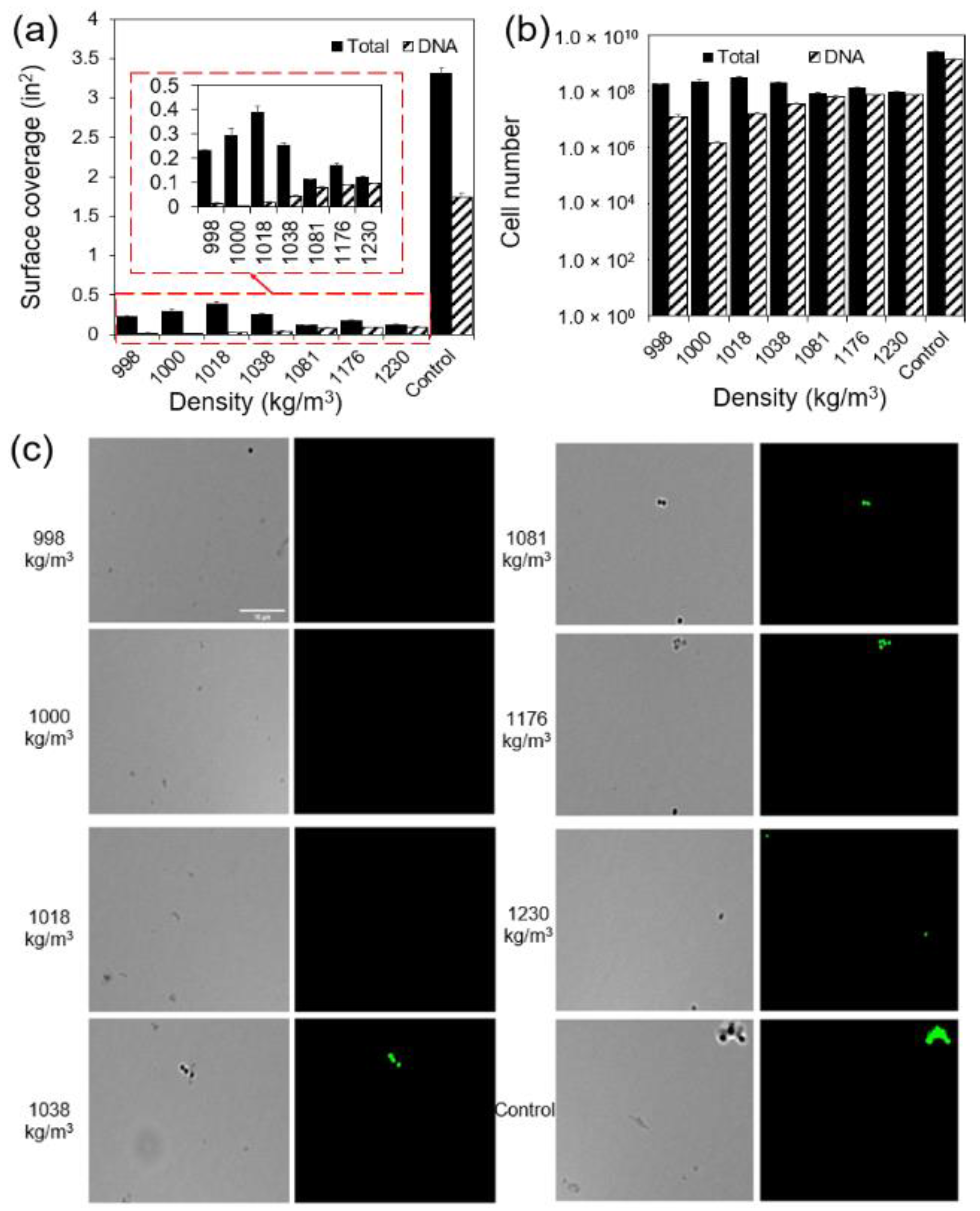
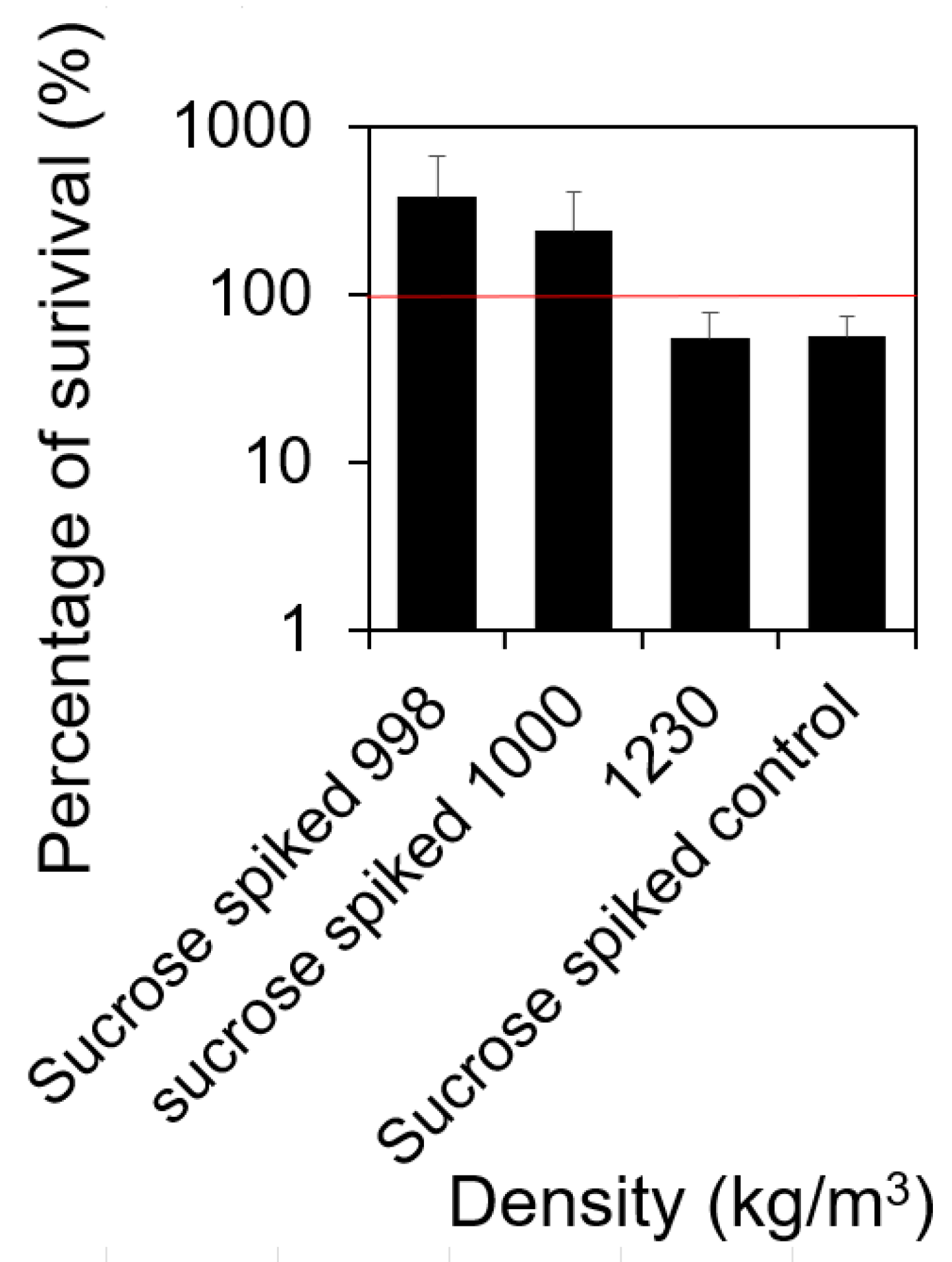
2.3. Cells with Higher Density Formed More Biofilms at the Liquid–Solid Interface Compared to Those with Lower Mass Density
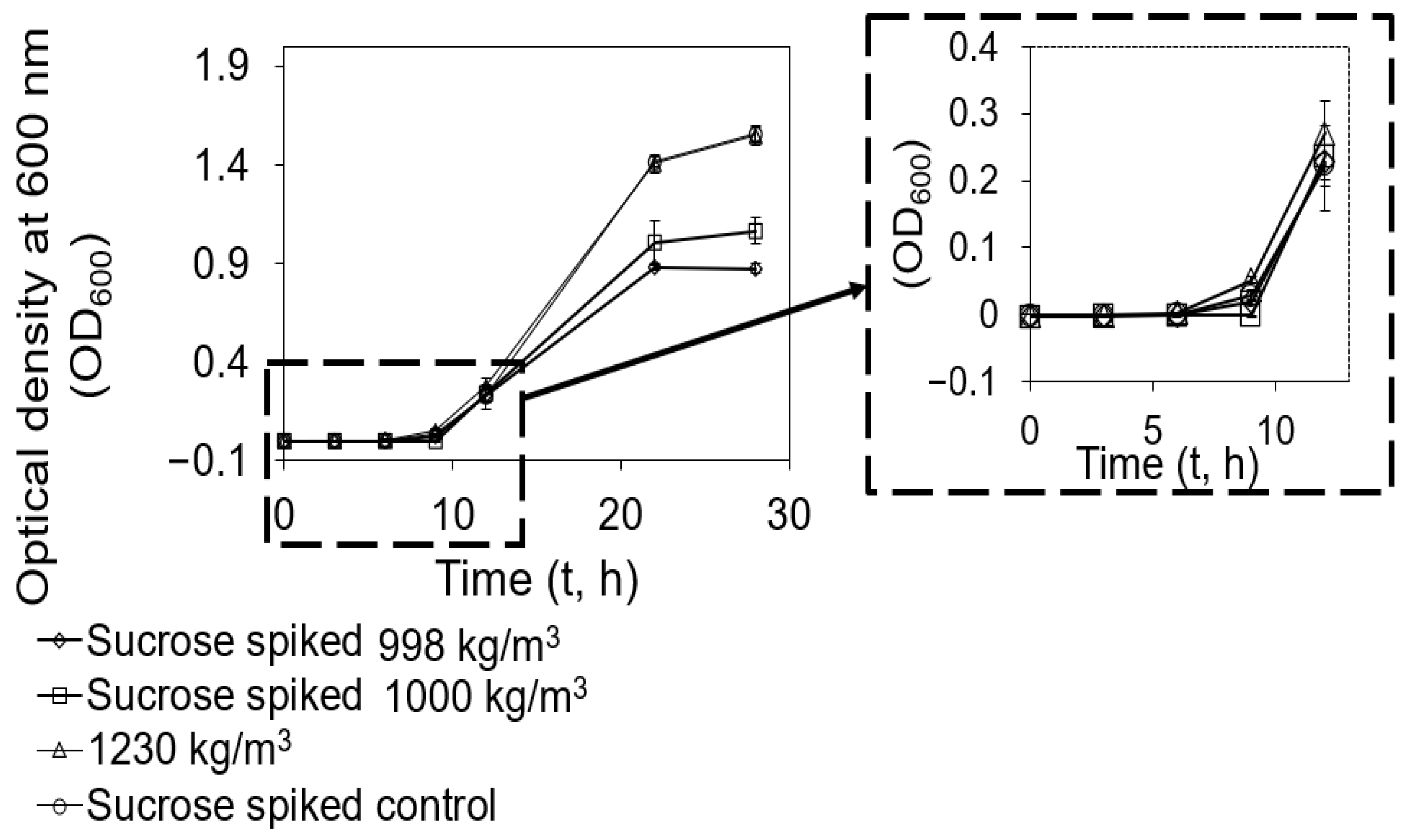
2.4. Cells with Higher Density Revived Faster from Nutrient Deprivation Than Those with Lower Density When Nutrients Were Supplemented
2.5. Higher Percentage of Cells with Higher Buoyant Density Had DNA
2.6. Cells with Higher Buoyant Density Were More Susceptible to Antibiotics
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Strains and Growth Media
5.2. Density Gradient Centrifugation Separation
5.3. Gravity-Driven Sedimentation
5.4. Biofilm Inoculation
5.5. Antibiotic Susceptibility and Most Probable Number (MPN) Assay
5.6. DNA Level Quantification Using Acridine Orange (AO) and Fluorescence Microscope
5.7. Statistics
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fulaz, S.; Vitale, S.; Quinn, L.; Casey, E. Nanoparticle–Biofilm Interactions: The Role of the EPS Matrix. Trends Microbiol. 2019, 27, 915–926. [Google Scholar] [CrossRef] [PubMed]
- Desai, N.; Ardekani, A.M. Biofilms at interfaces: Microbial distribution in floating films. Soft Matter. 2020, 16, 1731–1750. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.-C.; van Hullebusch, E.D.; Neu, T.R.; Nielsen, P.H.; Seviour, T.; Stoodley, P.; Wingender, J.; Wuertz, S. The biofilm matrix: Multitasking in a shared space. Nat. Rev. Microbiol. 2022, 21, 70–86. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K. Riddle of Biofilm Resistance. Antimicrob. Agents 2001, 45, 999–1007. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.W.; Mah, T.F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef] [PubMed]
- Manuse, S.; Shan, Y.; Canas-Duarte, S.J.; Bakshi, S.; Sun, W.S.; Mori, H.; Paulsson, J.; Lewis, K. Bacterial persisters are a stochastically formed subpopulation of low-energy cells. PLoS Biol. 2021, 19, e3001194. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Peng, B.; Qu, J.; Zheng, J. Are Bacterial Persisters Dormant Cells Only? Front. Microbiol. 2021, 12, 708580. [Google Scholar] [CrossRef]
- Romling, U.; Kjelleberg, S.; Normark, S.; Nyman, L.; Uhlin, B.E.; Akerlund, B. Microbial biofilm formation: A need to act. J. Intern. Med. 2014, 276, 98–110. [Google Scholar] [CrossRef]
- Wolcott, R.D.; Rhoads, D.D.; Bennett, M.E.; Wolcott, B.M.; Gogokhia, L.; Costerton, J.W.; Dowd, S.E. Chronic wounds and the medical biofilm paradigm. J. Wound Care 2010, 19, 45–53. [Google Scholar] [CrossRef]
- Sharma, S.; Mohler, J.; Mahajan, S.D.; Schwartz, S.A.; Bruggemann, L.; Aalinkeel, R. Microbial Biofilm: A Review on Formation, Infection, Antibiotic Resistance, Control Measures, and Innovative Treatment. Microorganisms 2023, 11, 1614. [Google Scholar] [CrossRef]
- Lee, S.W.; Phillips, K.S.; Gu, H.; Kazemzadeh-Narbat, M.; Ren, D. How microbes read the map: Effects of implant topography on bacterial adhesion and biofilm formation. Biomaterials 2021, 268, 120595. [Google Scholar] [CrossRef] [PubMed]
- Howden, B.P.; Giulieri, S.G.; Wong Fok Lung, T.; Baines, S.L.; Sharkey, L.K.; Lee, J.Y.H.; Hachani, A.; Monk, I.R.; Stinear, T.P. Staphylococcus aureus host interactions and adaptation. Nat. Rev. Microbiol. 2023, 21, 380–395. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Nishimura, M.; Nayak, B.B.; Kogure, K. Separation of marine bacteria according to buoyant density by use of the density-dependent cell sorting method. Appl. Env. Microbiol. 2007, 73, 1049–1053. [Google Scholar] [CrossRef] [PubMed]
- Kubitschek, H.E.; Baldwin, W.W.; Graetzer, R. Buoyant density constancy during the cell cycle of Escherichia coli. J. Bacteriol. 1983, 155, 1027–1032. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, W.W.; Kubitschek, H.E. Buoyant density variation during the cell cycle of Saccharomyces cerevisiae. J. Bacteriol. 1984, 158, 701–704. [Google Scholar] [CrossRef]
- Martínez-Salas, E.; Martín, J.A.; Vicente, M. Relationship of Escherichia coli density to growth rate and cell age. J. Bacteriol. 1981, 147, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Choudhary, A.; Bavishi, A.; Ogbonna, N.; Maddux, S.; Choudhary, M. Use of the sucrose gradient method for bacterial cell cycle synchronization. J. Microbiol. Biol. Educ. 2012, 13, 50–53. [Google Scholar] [CrossRef]
- Schrader, J.M.; Shapiro, L. Synchronization of Caulobacter crescentus for investigation of the bacterial cell cycle. J. Vis. Exp. 2015, 98, e52633. [Google Scholar] [CrossRef]
- Wilmaerts, D.; Etthel, M.W.; Verstraeten, N.; Michiels, A.J. General Mechanisms Leading to Persister Formation and Awakening. Trends Genet. 2019, 35, 401–411. [Google Scholar] [CrossRef]
- Kim, J.S.; Chowdhury, N.; Yamasaki, R.; Wood, T.K. Viable but non-culturable and persistence describe the same bacterial stress state. Env. Microbiol. 2018, 20, 2038–2048. [Google Scholar] [CrossRef]
- Johnson, M.E.; Montoro Bustos, A.R.; Winchester, M.R. Practical utilization of spICP-MS to study sucrose density gradient centrifugation for the separation of nanoparticles. Anal. Bioanal. Chem. 2016, 408, 7629–7640. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chandra, K.; Kumar, V.; Werner, S.E.; Odom, T.W. Separation of Stabilized MOPS Gold Nanostars by Density Gradient Centrifugation. ACS Omega 2017, 2, 4878–4884. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-López, V.; Fernández-Romero, J.M.; Gómez-Hens, A. Evaluation of liposome populations using a sucrose density gradient centrifugation approach coupled to a continuous flow system. Anal. Chim. Acta 2009, 645, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.M. Isolation of Golgi membranes from tissues and cells by differential and density gradient centrifugation. Curr. Protoc. Cell Biol. 2001, 3.9, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Hadjiolov, A.A.; Venkov, P.V.; Tsanev, R.G. Ribonucleic acids fractionation by density-gradient centrifugation and by agar gel electrophoresis: A comparison. Anal. Biochem. 1966, 17, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Zwartouw, H.T.; Westwood, J.C.; Appleyard, G. Purification of pox viruses by density gradient centrifugation. J. Gen. Microbiol. 1962, 29, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Brakke, M.K. Nonideal sedimentation and the capacity of sucrose gradient columns for virus in density-gradient centrifugation. Arch. Biochem. Biophys. 1964, 107, 388–403. [Google Scholar] [CrossRef]
- Parlet, C.P.; Brown, M.M.; Horswill, A.R. Commensal Staphylococci Influence Staphylococcus aureus Skin Colonization and Disease. Trends Microbiol. 2019, 27, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Filkins, L.M.; Graber, J.A.; Olson, D.G.; Dolben, E.L.; Lynd, L.R.; Bhuju, S.; O’Toole, G.A. Coculture of Staphylococcus aureus with Pseudomonas aeruginosa Drives S. aureus towards Fermentative Metabolism and Reduced Viability in a Cystic Fibrosis Model. J. Bacteriol. 2015, 197, 2252–2264. [Google Scholar] [CrossRef]
- Ciupescu, L.M.; Auvray, F.; Nicorescu, I.M.; Meheut, T.; Ciupescu, V.; Lardeux, A.L.; Tanasuica, R.; Hennekinne, J.A. Characterization of Staphylococcus aureus strains and evidence for the involvement of non-classical enterotoxin genes in food poisoning outbreaks. FEMS Microbiol. Lett. 2018, 365, fny139. [Google Scholar] [CrossRef]
- Pollitt, E.J.; Crusz, S.A.; Diggle, S.P. Staphylococcus aureus forms spreading dendrites that have characteristics of active motility. Sci. Rep. 2015, 5, 17698. [Google Scholar] [CrossRef] [PubMed]
- Cartwright, J.H.E. Stokes’ law, viscometry, and the Stokes falling sphere clock. Philos. Trans. A Math. Phys. Eng. Sci. 2020, 378, 20200214. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Lee, S.W.; Carnicelli, J.; Jiang, Z.; Ren, D. Antibiotic Susceptibility of Escherichia coli Cells during Early-Stage Biofilm Formation. J. Bacteriol. 2019, 201, e00034-19. [Google Scholar] [CrossRef]
- Naghili, H.; Tajik, H.; Mardani, K.; Razavi Rouhani, S.M.; Ehsani, A.; Zare, P. Validation of drop plate technique for bacterial enumeration by parametric and nonparametric tests. Vet. Res. Forum 2013, 4, 179–183. [Google Scholar]
- O’Toole, G.A. Microtiter dish biofilm formation assay. J. Vis. Exp. 2011, 47, e2437. [Google Scholar] [CrossRef]
- Schichnes, D.; Nemson, J.A.; Ruzin, S.E. Fluorescent Staining Method for Bacterial Endospores. Microscope 2006, 54, 91–93. [Google Scholar]
- Sutton, S. The Most Probable Number Method and Its Uses in Enumeration, Qualification, and Validation. J. Valid. Technol. 2010, 16, 35–38. [Google Scholar]
- Idrees, M.; Sawant, S.; Karodia, N.; Rahman, A. Biofilm: Morphology, Genetics, Pathogenesis and Treatment Strategies. Int. J. Env. Res. Public Health 2021, 18, 7602. [Google Scholar] [CrossRef]
- Hindy, J.R.; Haddad, S.F.; Kanj, S.S. New drugs for methicillin-resistant Staphylococcus aureus skin and soft tissue infections. Curr. Opin. Infect. Dis. 2022, 35, 112–119. [Google Scholar] [CrossRef]
- Arciola, C.R.; Campoccia, D.; Montanaro, L. Implant infections: Adhesion, biofilm formation and immune evasion. Nat. Rev. Microbiol. 2018, 16, 397–409. [Google Scholar] [CrossRef]
- Mah, T.F.; O’Toole, G.A. Mechanisms of Biofilm Resistance to Antimicrobial Agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Stewart, P.S. Mechanisms of antibiotic resistance in bacterial biofilms. Int. J. Med. Microbiol. 2002, 292, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Mahto, K.U.; Das, S. Bacterial biofilm and extracellular polymeric substances in the moving bed biofilm reactor for wastewater treatment: A review. Bioresour. Technol. 2022, 345, 126476. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, M.H.; Idris, A.L.; Fan, X.; Guo, Y.; Yu, Y.; Jin, X.; Qiu, J.; Guan, X.; Huang, T. Beyond Risk: Bacterial Biofilms and Their Regulating Approaches. Front. Microbiol. 2020, 11, 928. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Ren, D. A novel inductively coupled capacitor wireless sensor system for rapid antibiotic susceptibility testing. J. Biol. Eng. 2023, 17, 54. [Google Scholar] [CrossRef]
- Gu, H.; Lee, S.W.; Carnicelli, J.; Zhang, T.; Ren, D. Magnetically driven active topography for long-term biofilm control. Nat. Commun. 2020, 11, 2211. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Lee, S.W.; Buffington, S.L.; Henderson, J.H.; Ren, D. On-Demand Removal of Bacterial Biofilms via Shape Memory Activation. ACS Appl. Mater. Interfaces 2016, 8, 21140–21144. [Google Scholar] [CrossRef]
- Johnson, M.E.; Bustos, A.R.M.; Hanna, S.K.; Petersen, E.J.; Murphy, K.E.; Yu, L.L.; Nelson, B.C.; Winchester, M.R. Sucrose density gradient centrifugation for efficient separation of engineered nanoparticles from a model organism, Caenorhabditis elegans. NIST Special Publication 1200-24. Natl. Inst. Stand. Technol. 2017. [Google Scholar] [CrossRef]
- Damas-Souza, D.M.; Nunes, R.; Carvalho, H.F. An improved acridine orange staining of DNA/RNA. Acta Histochem. 2019, 121, 450–454. [Google Scholar] [CrossRef]
- Lee, S.W.; Gu, H.; Kilberg, J.B.; Ren, D. Sensitizing bacterial cells to antibiotics by shape recovery triggered biofilm dispersion. Acta Biomater. 2018, 81, 93–102. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kispert, S.; Liguori, M.; Velikaneye, C.; Qiu, C.; Wang, S.; Zhang, N.; Gu, H. Role of Staphylococcus aureus’s Buoyant Density in the Development of Biofilm Associated Antibiotic Susceptibility. Microorganisms 2024, 12, 759. https://doi.org/10.3390/microorganisms12040759
Kispert S, Liguori M, Velikaneye C, Qiu C, Wang S, Zhang N, Gu H. Role of Staphylococcus aureus’s Buoyant Density in the Development of Biofilm Associated Antibiotic Susceptibility. Microorganisms. 2024; 12(4):759. https://doi.org/10.3390/microorganisms12040759
Chicago/Turabian StyleKispert, Sarah, Madison Liguori, Cody Velikaneye, Chong Qiu, Shue Wang, Nan Zhang, and Huan Gu. 2024. "Role of Staphylococcus aureus’s Buoyant Density in the Development of Biofilm Associated Antibiotic Susceptibility" Microorganisms 12, no. 4: 759. https://doi.org/10.3390/microorganisms12040759
APA StyleKispert, S., Liguori, M., Velikaneye, C., Qiu, C., Wang, S., Zhang, N., & Gu, H. (2024). Role of Staphylococcus aureus’s Buoyant Density in the Development of Biofilm Associated Antibiotic Susceptibility. Microorganisms, 12(4), 759. https://doi.org/10.3390/microorganisms12040759