
A Comparison of Different Protocols for the Extraction of Microbial DNA Inhabiting Synthetic Mars Simulant Soil

 , , and
, , and 
Abstract
:1. Introduction
2. Material and Methods
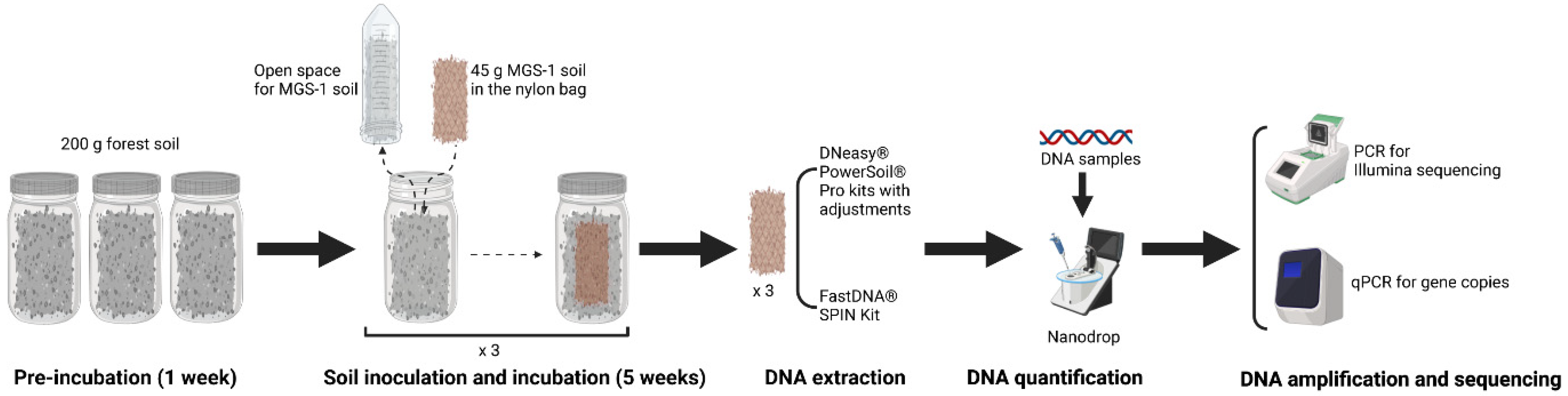
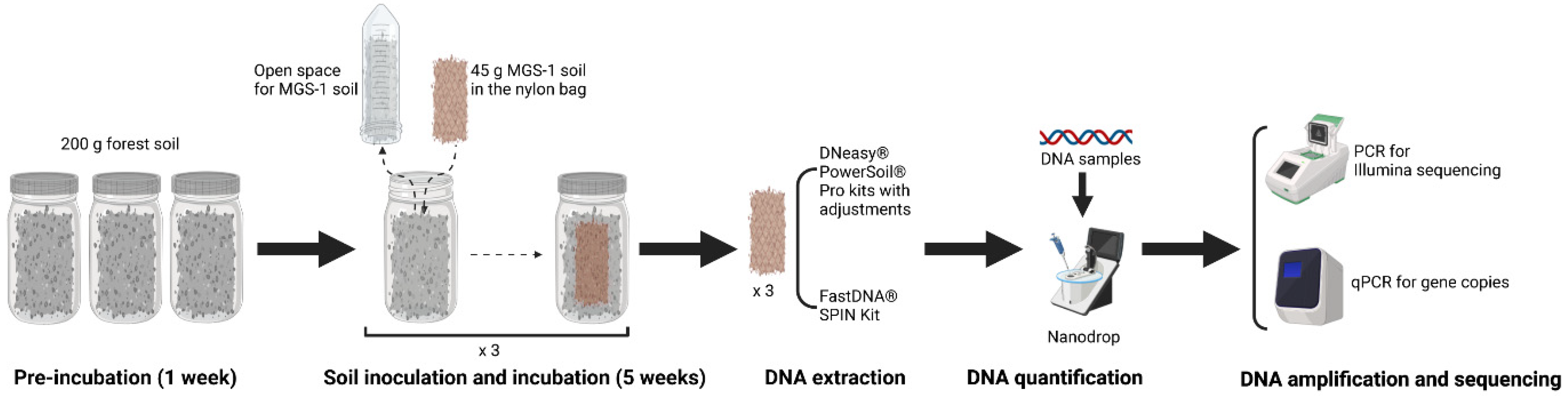
2.1. Soil Sampling
2.2. Soil Sample Preparation
2.3. Soil DNA Extraction
2.3.1. PowerSoil DNA Isolation Kit
2.3.2. FastDNA SPIN Kit
2.4. High-Throughput Sequencing of Bacterial Community Composition
2.5. Sequence Read Processing and Data Analysis
2.6. Real-Time Quantitative PCR (qPCR)
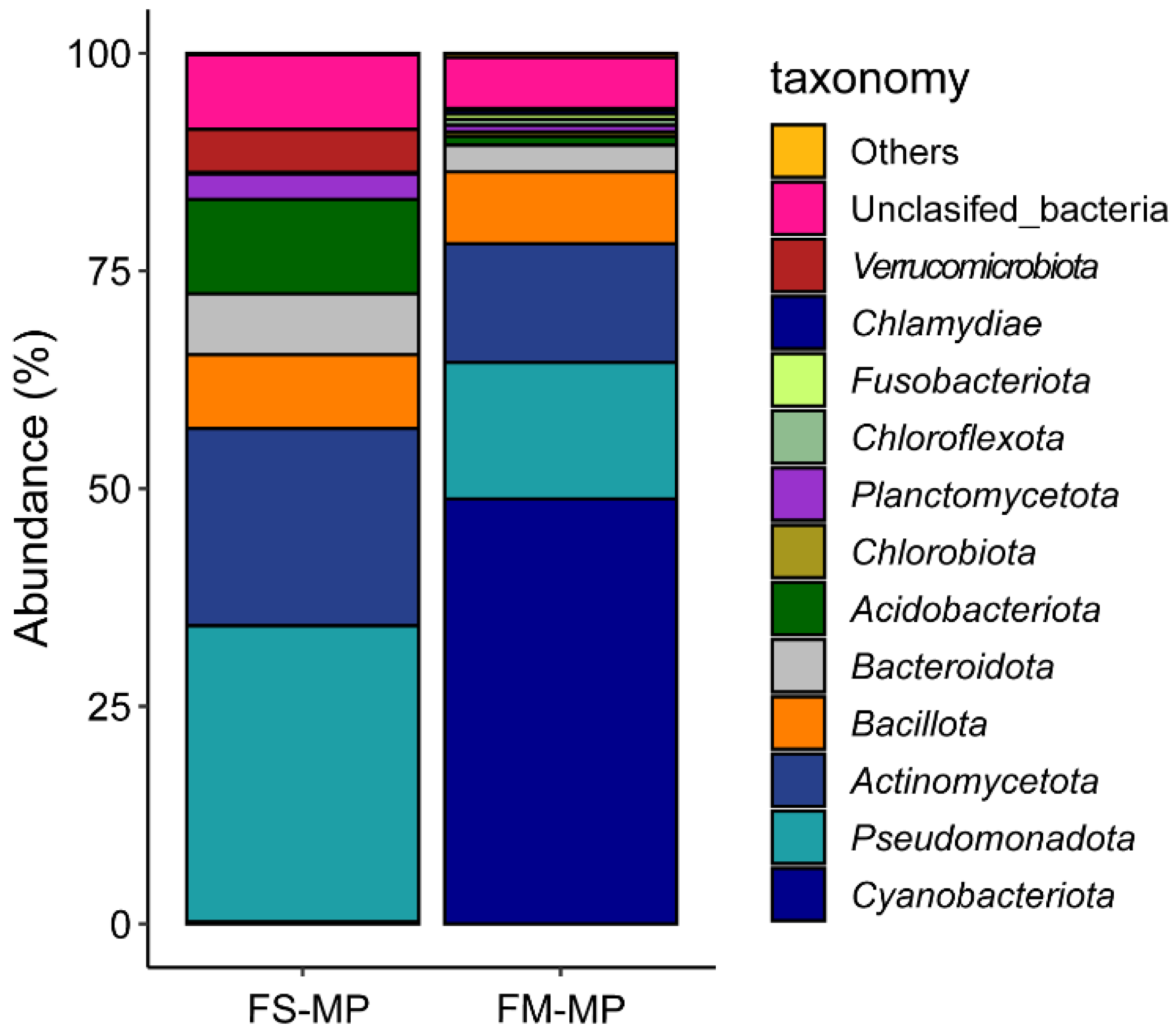
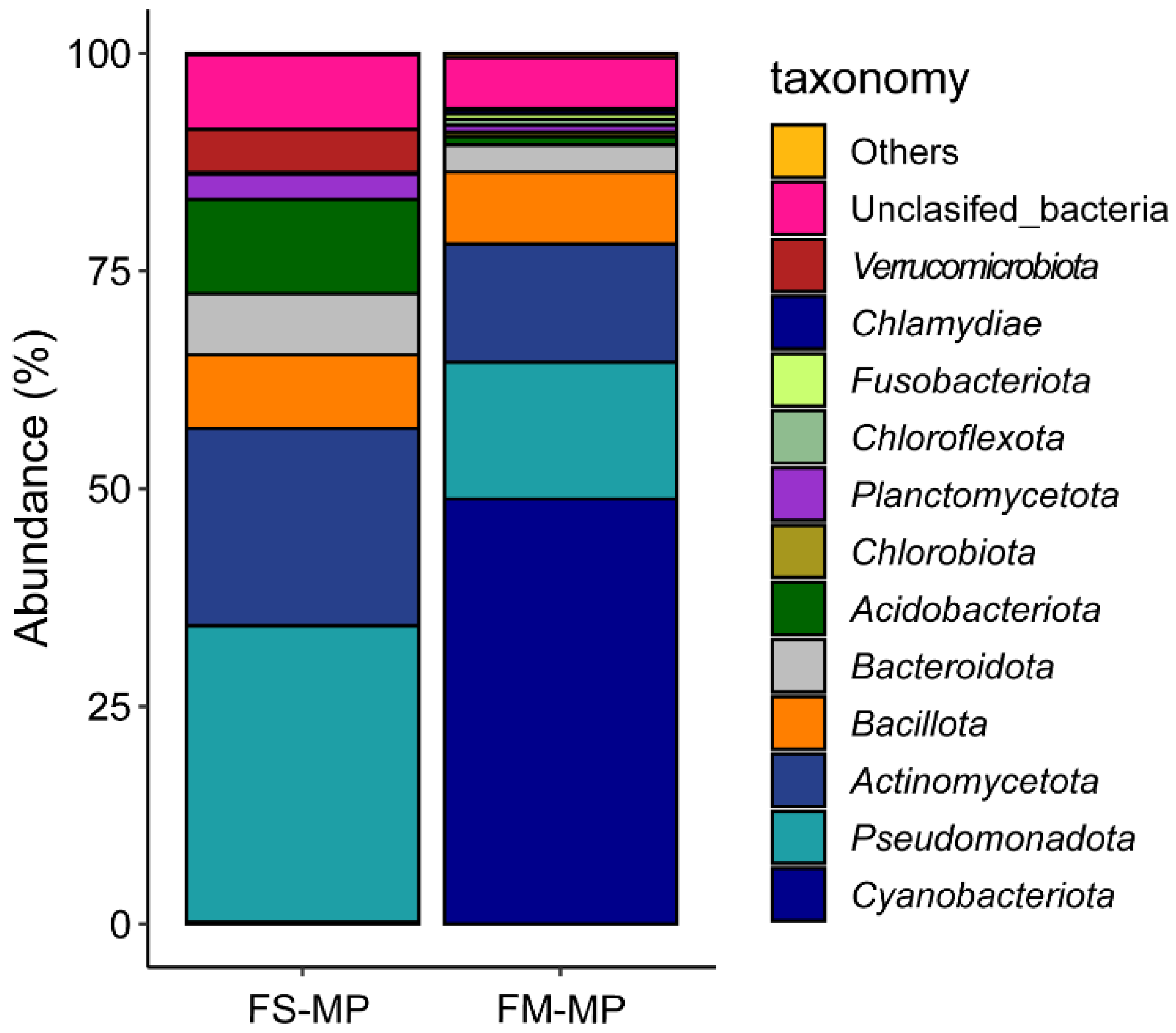
3. Result
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marlow, J.J.; Martins, Z.; A Sephton, M. Mars on Earth: Soil analogues for future Mars missions. Astron. Geophys. 2008, 49, 2.20–2.23. [Google Scholar] [CrossRef]
- Morris, R.V.; Klingelhöfer, G.; Schröder, C.; Rodionov, D.S.; Yen, A.; Ming, D.W.; de Souza, P.A.; Fleischer, I.; Wdowiak, T.; Gellert, R.; et al. Mössbauer mineralogy of rock, soil, and dust at Gusev crater, Mars: Spirit’s journey through weakly altered olivine basalt on the plains and pervasively altered basalt in the Columbia Hills. J. Geophys. Res. Planets 2006, 111, E02S13. [Google Scholar] [CrossRef]
- Morris, R.V.; Klingelhöfer, G.; Schröder, C.; Rodionov, D.S.; Yen, A.; Ming, D.W.; de Souza, P.A.; Wdowiak, T.; Fleischer, I.; Gellert, R.; et al. Mössbauer mineralogy of rock, soil, and dust at Meridiani Planum, Mars: Opportunity’s journey across sulfate-rich outcrop, basaltic sand and dust, and hematite lag deposits. J. Geophys. Res. Planets 2006, 111, E12S15. [Google Scholar] [CrossRef]
- Yen, A.S.; Gellert, R.; Schröder, C.; Morris, R.V.; Bell, J.F.; Knudson, A.T.; Clark, B.C.; Ming, D.W.; Crisp, J.A.; Arvidson, R.E.; et al. An integrated view of the chemistry and mineralogy of martian soils. Nature 2005, 436, 49–54. [Google Scholar] [CrossRef]
- Zeng, X.; Li, X.; Wang, S.; Li, S.; Spring, N.; Tang, H.; Li, Y.; Feng, J. JMSS-1: A new Martian soil simulant. Earth Planets Space 2015, 67, 72. [Google Scholar] [CrossRef]
- Westall, F.; Foucher, F.; Bost, N.; Bertrand, M.; Loizeau, D.; Vago, J.L.; Kminek, G.; Gaboyer, F.; Campbell, K.A.; Bréhéret, J.-G.; et al. Biosignatures on Mars: What, Where, and How? Implications for the Search for Martian Life. Astrobiology 2015, 15, 998–1029. [Google Scholar] [CrossRef]
- Vago, J.L.; Westall, F.; Coates, A.J.; Jaumann, R.; Korablev, O.; Ciarletti, V.; Mitrofanov, I.; Josset, J.-L.; De Sanctis, M.C.; Bibringet, J.-P.; et al. Habitability on Early Mars and the Search for Biosignatures with the ExoMars Rover. Astrobiology 2017, 17, 471–510. [Google Scholar] [CrossRef]
- Carrier, B.; Beaty, D.; Meyer, M.; Blank, J.; Chou, L.; DasSarma, S.; Marais, D.D.; Eigenbrode, J.; Grefenstette, N.; Lanza, N.; et al. Mars Extant Life: What’s Next? Conference Report. Astrobiology 2020, 20, 785–814. [Google Scholar] [CrossRef]
- ElShafie, A.; Chevrier, V.; Dennis, N. Application of planetary analog mechanical properties to subsurface geological investigations. Planet. Space Sci. 2012, 73, 224–232. [Google Scholar] [CrossRef]
- Moroz, L.; Basilevsky, A.; Hiroi, T.; Rout, S.; Baither, D.; van der Bogert, C.; Yakovlev, O.; Fisenko, A.; Semjonova, L.; Rusakov, V.; et al. Spectral properties of simulated impact glasses produced from martian soil analogue JSC Mars-1. Icarus 2009, 202, 336–353. [Google Scholar] [CrossRef]
- Perko, H.A.; Nelson, J.D.; Green, J.R. Mars Soil Mechanical Properties and Suitability of Mars Soil Simulants. J. Aerosp. Eng. 2006, 19, 169–176. [Google Scholar] [CrossRef]
- Yeomans, B.; Saaj, C.M.; Van Winnendael, M. Walking planetary rovers—Experimental analysis and modelling of leg thrust in loose granular soils. J. Terramechanics 2013, 50, 107–120. [Google Scholar] [CrossRef]
- Cannon, K.M.; Britt, D.T.; Smith, T.M.; Fritsche, R.F.; Batcheldor, D. Mars global simulant MGS-1: A Rocknest-based open standard for basaltic martian regolith simulants. Icarus 2019, 317, 470–478. [Google Scholar] [CrossRef]
- Wamelink, G.; Frissel, J.; Krijnen, W.; Verwoert, M. Crop growth and viability of seeds on Mars and Moon soil simulants. Open Agric. 2019, 4, 509–516. [Google Scholar] [CrossRef]
- Wamelink, G.W.W.; Frissel, J.Y.; Krijnen, W.H.J.; Verwoert, M.R.; Goedhart, P.W. Can Plants Grow on Mars and the Moon: A Growth Experiment on Mars and Moon Soil Simulants. PLoS ONE 2014, 9, e103138. [Google Scholar] [CrossRef]
- Harris, F.; Dobbs, J.; Atkins, D.; Ippolito, J.A.; Stewart, J.E. Soil fertility interactions with Sinorhizobium-legume symbiosis in a simulated Martian regolith; effects on nitrogen content and plant health. PLoS ONE 2021, 16, e0257053. [Google Scholar] [CrossRef]
- Duri, L.G.; Caporale, A.G.; Rouphael, Y.; Vingiani, S.; Palladino, M.; De Pascale, S.; Adamo, P. The Potential for Lunar and Martian Regolith Simulants to Sustain Plant Growth: A Multidisciplinary Overview. Front. Astron. Space Sci. 2022, 8, 747821. [Google Scholar] [CrossRef]
- Kasiviswanathan, P.; Swanner, E.D.; Halverson, L.J.; Vijayapalani, P. Farming on Mars: Treatment of basaltic regolith soil and briny water simulants sustains plant growth. PLoS ONE 2022, 17, e0272209. [Google Scholar] [CrossRef]
- Raghavendra, J.B.; Zorzano, M.-P.; Kumaresan, D.; Martin-Torres, J. DNA sequencing at the picogram level to investigate life on Mars and Earth. Sci. Rep. 2023, 13, 15277. [Google Scholar] [CrossRef]
- Mojarro, A.; Hachey, J.; Bailey, R.; Brown, M.; Doebler, R.; Ruvkun, G.; Zuber, M.T.; Carr, C.E. Nucleic Acid Extraction and Sequencing from Low-Biomass Synthetic Mars Analog Soils forIn SituLife Detection. Astrobiology 2019, 19, 1139–1152. [Google Scholar] [CrossRef]
- Mojarro, A.; Ruvkun, G.; Zuber, M.T.; Carr, C.E. Nucleic Acid Extraction from Synthetic Mars Analog Soils for in situ Life Detection. Astrobiology 2017, 17, 747–760. [Google Scholar] [CrossRef]
- Adhikari, S.; Timms, W.; Mahmud, M.P. Optimising water holding capacity and hydrophobicity of biochar for soil amendment—A review. Sci. Total Environ. 2022, 851, 158043. [Google Scholar] [CrossRef]
- Wang, L.; Yuan, L.; Li, Z.-H.; Zhang, X.; Sheng, G.-P. Quantifying the occurrence and transformation potential of extracellular polymeric substances (EPS)-associated antibiotic resistance genes in activated sludge. J. Hazard. Mater. 2021, 408, 124428. [Google Scholar] [CrossRef]
- Zakaria, B.S.; Dhar, B.R. Characterization and significance of extracellular polymeric substances, reactive oxygen species, and extracellular electron transfer in methanogenic biocathode. Sci. Rep. 2021, 11, 7933. [Google Scholar] [CrossRef]
- Kamble, A.; Singh, H. Different Methods of Soil DNA Extraction. Bio-Protocol 2020, 10, e3521. [Google Scholar] [CrossRef]
- Debeljak, P.; Pinto, M.; Proietti, M.; Reisser, J.; Ferrari, F.F.; Abbas, B.; van Loosdrecht, M.C.M.; Slat, B.; Herndl, G.J. Extracting DNA from ocean microplastics: A method comparison study. Anal. Methods 2017, 9, 1521–1526. [Google Scholar] [CrossRef]
- Xie, K.; Deng, Y.; Zhang, X.; Wang, X.; Kang, G.; Bai, L.; Huang, H. Biases in Prokaryotic Community Amplicon Sequencing Affected by DNA Extraction Methods in Both Saline and Non-saline Soil. Front. Microbiol. 2018, 9, 1796. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-Learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Anonymous. Biomformat: An Interface Package for the BIOM File Format Version 1.18.0 from Bioconductor. Available online: https://rdrr.io/bioc/biomformat/ (accessed on 10 October 2023).
- Schloss, P.D. Rarefaction is currently the best approach to control for uneven sequencing effort in amplicon sequence analyses. mSphere 2024, 9, e0035423. [Google Scholar] [CrossRef]
- Louca, S.; Jacques, S.M.S.; Pires, A.P.F.; Leal, J.S.; Srivastava, D.S.; Parfrey, L.W.; Farjalla, V.F.; Doebeli, M. High taxonomic variability despite stable functional structure across microbial communities. Nat. Ecol. Evol. 2016, 1, 0015. [Google Scholar] [CrossRef]
- Mitsuta, A.; Ishige, N.; Tatsumi, C.; Madegwa, Y.M.; Kuramae, E.E.; Uchida, Y. Stability of ammonia oxidizer communities upon nitrogen fertilizer pulse disturbances is dependent on diversity. Geoderma 2023, 439, 116685. [Google Scholar] [CrossRef]
- Direito, S.O.; Marees, A.; Röling, W.F. Sensitive life detection strategies for low-biomass environments: Optimizing extraction of nucleic acids adsorbing to terrestrial and Mars analogue minerals. FEMS Microbiol. Ecol. 2012, 81, 111–123. [Google Scholar] [CrossRef]
- Xia, Z.; Patchin, M.; McKay, C.P.; Drndić, M. Deoxyribonucleic Acid Extraction from Mars Analog Soils and Their Characterization with Solid-State Nanopores. Astrobiology 2022, 22, 992–1008. [Google Scholar] [CrossRef]
- Ramírez, G.A.; Graham, D.; D’Hondt, S. Influence of commercial DNA extraction kit choice on prokaryotic community metrics in marine sediment. Limnol. Oceanogr. Methods 2018, 16, 525–536. [Google Scholar] [CrossRef]
- Santos, H.; Carmo, F.; Leite, D.; Jesus, H.; Maalouf, P.D.C.; Almeida, C.; Soriano, A.; Altomari, D.; Suhett, L.; Vólaro, V.; et al. Comparison of different protocols for the extraction of microbial DNA from reef corals. Braz. J. Microbiol. 2012, 43, 517–527. [Google Scholar] [CrossRef]
- Peters, G.H.; Abbey, W.; Bearman, G.H.; Mungas, G.S.; Smith, J.A.; Anderson, R.C.; Douglas, S.; Beegle, L.W. Mojave Mars simulant—Characterization of a new geologic Mars analog. Icarus 2008, 197, 470–479. [Google Scholar] [CrossRef]
- Quinn, R.C.; Martucci, H.F.; Miller, S.R.; Bryson, C.E.; Grunthaner, F.J.; Grunthaner, P.J. Perchlorate Radiolysis on Mars and the Origin of Martian Soil Reactivity. Astrobiology 2013, 13, 515–520. [Google Scholar] [CrossRef]
- Clark, B.C.; Kounaves, S.P. Evidence for the distribution of perchlorates on Mars. Int. J. Astrobiol. 2016, 15, 311–318. [Google Scholar] [CrossRef]
- Sutter, B.; Quinn, R.C.; Archer, P.D.; Glavin, D.P.; Glotch, T.D.; Kounaves, S.P.; Osterloo, M.M.; Rampe, E.B.; Ming, D.W. Measurements of Oxychlorine species on Mars. Int. J. Astrobiol. 2017, 16, 203–217. [Google Scholar] [CrossRef]
- Direito, S.O.; Ehrenfreund, P.; Marees, A.; Staats, M.; Foing, B.; Röling, W.F. A wide variety of putative extremophiles and large beta-diversity at the Mars Desert Research Station (Utah). Int. J. Astrobiol. 2011, 10, 191–207. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| ID | MGS-1 | Mars |
|---|---|---|
| SiO2 | 45.57% | 43.52% |
| Fe2O3 | 16.85% | 18.28% |
| Al2O3 | 9.43% | 8.64% |
| CaO | 4.03% | 6.09% |
| MgO | 16.50% | 6.54% |
| SO3 | 2.63% | 6.42% |
| Na2O | 3.66% | 2.57% |
| P2O5 | 0.37% | 0.79% |
| TiO2 | 0.30% | 0.78% |
| K2O | 0.43% | 0.35% |
| MnO | 0.10% | 0.32% |
| Cr2O3 | 0.12% | 0.37% |
| Total | 99.99% | 94.67% |
| Sample ID | DNA Yield (ng/µL) | A260/280 * | A260/230 * | Method | Number of Reads | 16S rRNA Gene Copies/g | |
|---|---|---|---|---|---|---|---|
| FM-1 | 1.8 | 1.74 | 0.23 | Normal | 0 | 0 | |
| FM-2 | 1.8 | 1.11 | 0.33 | Normal | 0 | 0 | |
| FM-3 | 3.2 | 1.2 | 0.17 | Normal | 0 | 0 | |
| FM-B1 | 68.8 | 1.55 | 0.4 | Beating | 0 | 0 | |
| FM-B2 | 12.5 | 1.61 | 0.18 | Beating | 0 | 0 | |
| FM-B3 | 68.8 | 1.55 | 0.4 | Beating | 0 | 0 | |
| FM-H1 | 0.5 | 1.91 | 0.05 | Heating | 0 | 0 | |
| FM-H2 | 1.6 | 1.21 | 0.08 | Heating | 0 | 0 | |
| PowerSoil | FM-H3 | 1.2 | 1.56 | 0.01 | Heating | 0 | 0 |
| FM-CER1 | 5 | 0.75 | 0.03 | CER | 0 | 0 | |
| FM-CER2 | 2.4 | 2.25 | 0.02 | CER | 0 | 0 | |
| FM-CER3 | 6 | 0.87 | 0.06 | CER | 0 | 0 | |
| FM-P1 | 9 | 1.62 | 0.03 | PBS | 0 | 0 | |
| FM-P2 | 13 | 1.89 | 0.13 | PBS | 0 | 0 | |
| FM-P3 | 9 | 3.15 | 0.03 | PBS | 0 | 0 | |
| FM-PC1 | 5 | 5.31 | 0.04 | PBS+CER | 0 | 0 | |
| FM-PC2 | 9 | 1.28 | 0.01 | PBS+CER | 0 | 0 | |
| FM-PC3 | 19 | 1.39 | 0.01 | PBS+CER | 0 | 0 | |
| FM-N1 | 1.3 | 1.59 | 0.04 | N2 | 0 | 0 | |
| FM-N2 | 1.1 | 1.93 | 0.03 | N2 | 0 | 0 | |
| FM-N3 | 3.2 | 1.47 | 0.08 | N2 | 0 | 0 | |
| FM-MP1 | 9.144 | 2.47 | 0.01 | Normal | 28,642 | 30,380 | |
| MP | FM-MP2 | 9.331 | 2.7 | 0.01 | Normal | 29,743 | 38,550 |
| FM-MP3 | 8.391 | 2.62 | 0.01 | Normal | 37,805 | 42,070 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Pijl, A.; Liu, B.; Wamelink, W.; Korthals, G.W.; Costa, O.Y.A.; Kuramae, E.E. A Comparison of Different Protocols for the Extraction of Microbial DNA Inhabiting Synthetic Mars Simulant Soil. Microorganisms 2024, 12, 760. https://doi.org/10.3390/microorganisms12040760
Wang H, Pijl A, Liu B, Wamelink W, Korthals GW, Costa OYA, Kuramae EE. A Comparison of Different Protocols for the Extraction of Microbial DNA Inhabiting Synthetic Mars Simulant Soil. Microorganisms. 2024; 12(4):760. https://doi.org/10.3390/microorganisms12040760
Chicago/Turabian StyleWang, Han, Agata Pijl, Binbin Liu, Wieger Wamelink, Gerard W. Korthals, Ohana Y. A. Costa, and Eiko E. Kuramae. 2024. "A Comparison of Different Protocols for the Extraction of Microbial DNA Inhabiting Synthetic Mars Simulant Soil" Microorganisms 12, no. 4: 760. https://doi.org/10.3390/microorganisms12040760
APA StyleWang, H., Pijl, A., Liu, B., Wamelink, W., Korthals, G. W., Costa, O. Y. A., & Kuramae, E. E. (2024). A Comparison of Different Protocols for the Extraction of Microbial DNA Inhabiting Synthetic Mars Simulant Soil. Microorganisms, 12(4), 760. https://doi.org/10.3390/microorganisms12040760