
Oral Administration of Lactobacillus acidophilus LA5 Prevents Alveolar Bone Loss and Alters Oral and Gut Microbiomes in a Murine Periodontitis Experimental Model
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Group Allocation
2.2. Sample Size
2.3. Bacteria Cultures
2.4. Experimental Periodontitis and Probiotic Administration
2.5. Sample Collection
2.6. Microcomputed Tomography (μCT) Analysis
2.7. Oral and Gut Microbiome Sequencing
2.8. Statistical Analysis
3. Results
3.1. Effect of L. acidophilus on Controlling Alveolar Bone Loss
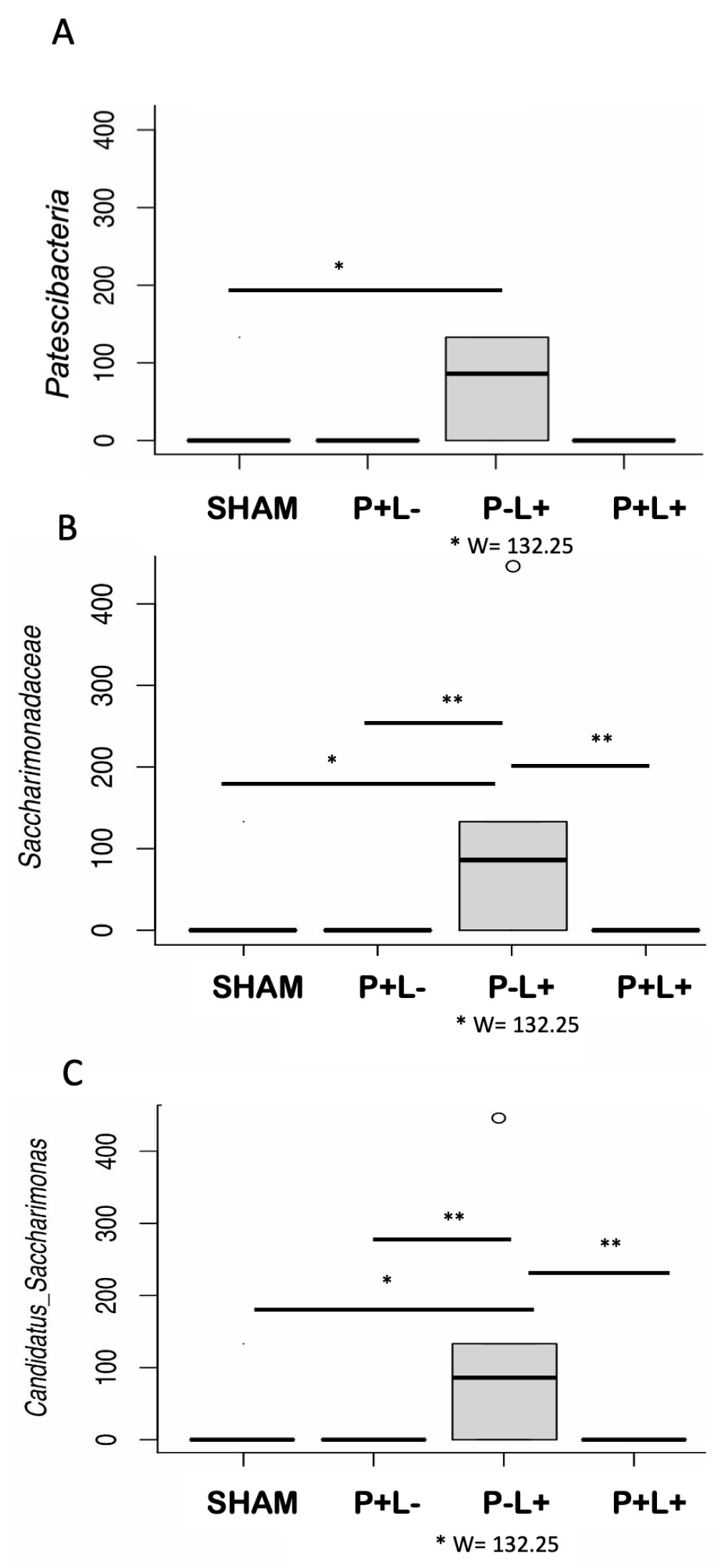
3.2. Effect of L. acidophilus on Reshaping Oral and Gut Microbiomes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hajishengallis, G.; Lamont, R.J. Beyond the Red Complex and into More Complexity: The Polymicrobial Synergy and Dysbiosis (PSD) Model of Periodontal Disease Etiology. Mol. Oral Microbiol. 2012, 27, 409–419. [Google Scholar] [CrossRef]
- Amado, P.P.P.; Kawamoto, D.; Albuquerque-Souza, E.; Franco, D.C.; Saraiva, L.; Casarin, R.C.V.; Horliana, A.C.R.T.; Mayer, M.P.A. Oral and Fecal Microbiome in Molar-Incisor Pattern Periodontitis. Front. Cell Infect. Microbiol. 2020, 10, 583761. [Google Scholar] [CrossRef]
- Arimatsu, K.; Yamada, H.; Miyazawa, H.; Minagawa, T.; Nakajima, M.; Ryder, M.I.; Gotoh, K.; Motooka, D.; Nakamura, S.; Iida, T.; et al. Oral Pathobiont Induces Systemic Inflammation and Metabolic Changes Associated with Alteration of Gut Microbiota. Sci. Rep. 2014, 4, 4828. [Google Scholar] [CrossRef]
- Kawamoto, D.; Borges, R.; Ribeiro, R.A.; de Souza, R.F.; Amado, P.P.P.; Saraiva, L.; Horliana, A.C.R.T.; Faveri, M.; Mayer, M.P.A. Oral Dysbiosis in Severe Forms of Periodontitis Is Associated with Gut Dysbiosis and Correlated with Salivary Inflammatory Mediators: A Preliminary Study. Front. Oral Health 2021, 2, 722495. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Raff, A.; Hunt, L.C. Probiotics for Periodontal Health: A Review of the Literature. J. Dent. Hyg. 2012, 86, 71–81. [Google Scholar]
- Gomes, A.C.; Bueno, A.A.; de Souza, R.G.M.; Mota, J.F. Gut Microbiota, Probiotics and Diabetes. Nutr. J. 2014, 13, 60. [Google Scholar] [CrossRef]
- Mousquer, C.R.; Della Bona, A.; Milani, D.C.; Callegari-Jacques, S.M.; Ishikawa, K.; Mayer, M.P.A.; Rösing, C.K.; Fornari, F. Are Lactobacillus salivarius G60 and Inulin More Efficacious to Treat Patients with Oral Halitosis and Tongue Coating than the Probiotic Alone and Placebo? A Randomized Clinical Trial. J. Periodontol. 2020, 91, 775–783. [Google Scholar] [CrossRef]
- Wolowczuk, I.; Verwaerde, C.; Viltart, O.; Delanoye, A.; Delacre, M.; Pot, B.; Grangette, C. Feeding Our Immune System: Impact on Metabolism. Clin. Dev. Immunol. 2008, 2008, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Laleman, I.; Pauwels, M.; Quirynen, M.; Teughels, W. A Dual-strain Lactobacilli reuteri Probiotic Improves the Treatment of Residual Pockets: A Randomized Controlled Clinical Trial. J. Clin. Periodontol. 2020, 47, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Alanzi, A.; Honkala, S.; Honkala, E.; Varghese, A.; Tolvanen, M.; Söderling, E. Effect of Lactobacillus rhamnosus and Bifidobacterium lactis on Gingival Health, Dental Plaque, and Periodontopathogens in Adolescents: A Randomised Placebo-Controlled Clinical Trial. Benef. Microbes 2018, 9, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Gheisary, Z.; Mahmood, R.; Harri Shivanantham, A.; Liu, J.; Lieffers, J.R.L.; Papagerakis, P.; Papagerakis, S. The Clinical, Microbiological, and Immunological Effects of Probiotic Supplementation on Prevention and Treatment of Periodontal Diseases: A Systematic Review and Meta-Analysis. Nutrients 2022, 14, 1036. [Google Scholar] [CrossRef]
- Li, J.; Zhao, G.; Zhang, H.M.; Zhu, F.F. Probiotic Adjuvant Treatment in Combination with Scaling and Root Planing in Chronic Periodontitis: A Systematic Review and Meta-Analysis. Benef. Microbes 2023, 14, 95–108. [Google Scholar] [CrossRef]
- Asgharian, H.; Homayouni-Rad, A.; Mirghafourvand, M.; Mohammad-Alizadeh-Charandabi, S. Effect of Probiotic Yoghurt on Plasma Glucose in Overweight and Obese Pregnant Women: A Randomized Controlled Clinical Trial. Eur. J. Nutr. 2020, 59, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Rezazadeh, L.; Gargari, B.P.; Jafarabadi, M.A.; Alipour, B. Effects of Probiotic Yogurt on Glycemic Indexes and Endothelial Dysfunction Markers in Patients with Metabolic Syndrome. Nutrition 2019, 62, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Zarrati, M.; Shidfar, F.; Nourijelyani, K.; Mofid, V.; Hossein Zadeh-Attar, M.J.; Bidad, K.; Najafi, F.; Gheflati, Z.; Chamari, M.; Salehi, E. Lactobacillus acidophilus La5, Bifidobacterium BB12, and Lactobacillus casei DN001 Modulate Gene Expression of Subset Specific Transcription Factors and Cytokines in Peripheral Blood Mononuclear Cells of Obese and Overweight People. BioFactors 2013, 39, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Nabavi, S.; Rafraf, M.; Somi, M.H.; Homayouni-Rad, A.; Asghari-Jafarabadi, M. Effects of Probiotic Yogurt Consumption on Metabolic Factors in Individuals with Nonalcoholic Fatty Liver Disease. J. Dairy Sci. 2014, 97, 7386–7393. [Google Scholar] [CrossRef]
- Albuquerque-Souza, E.; Balzarini, D.; Ando-Suguimoto, E.S.; Ishikawa, K.H.; Simionato, M.R.L.; Holzhausen, M.; Mayer, M.P.A. Probiotics Alter the Immune Response of Gingival Epithelial Cells Challenged by Porphyromonas gingivalis. J. Periodontal Res. 2019, 54, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Bueno, M.R.; Ishikawa, K.H.; Almeida-Santos, G.; Ando-Suguimoto, E.S.; Shimabukuro, N.; Kawamoto, D.; Mayer, M.P.A. Lactobacilli Attenuate the Effect of Aggregatibacter actinomycetemcomitans Infection in Gingival Epithelial Cells. Front. Microbiol. 2022, 13, 846192. [Google Scholar] [CrossRef]
- Ishikawa, K.H.; Mita, D.; Kawamoto, D.; Nicoli, J.R.; Albuquerque-Souza, E.; Lorenzetti Simionato, M.R.; Mayer, M.P.A. Probiotics Alter Biofilm Formation and the Transcription of Porphyromonas gingivalis Virulence-Associated Genes. J. Oral Microbiol. 2020, 12, 1805553. [Google Scholar] [CrossRef]
- Ishikawa, K.H.; Bueno, M.R.; Kawamoto, D.; Simionato, M.R.L.; Mayer, M.P.A. Lactobacilli Postbiotics Reduce Biofilm Formation and Alter Transcription of Virulence Genes of Aggregatibacter actinomycetemcomitans. Mol. Oral Microbiol. 2021, 36, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Bueno, M.R.; Martins, F.H.; Rocha, C.M.; Kawamoto, D.; Ishikawa, K.H.; Ando-Suguimoto, E.S.; Carlucci, A.R.; Arroteia, L.S.; Casarin, R.V.; Mayer, M.P.A. Lactobacillus acidophilus LA-5 Ameliorates Inflammation and Alveolar Bone Loss Promoted by A. actinomycetemcomitans and S. gordonii in Mice and Impacts Oral and Gut Microbiomes. Microorganisms 2024, 12, 836. [Google Scholar] [CrossRef] [PubMed]
- Medina, M.; Izquierdo, E.; Ennahar, S.; Sanz, Y. Differential Immunomodulatory Properties of Bifidobacterium logum Strains: Relevance to Probiotic Selection and Clinical Applications. Clin. Exp. Immunol. 2007, 150, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Shimabukuro, N.; Cataruci, A.C.d.S.; Ishikawa, K.H.; de Oliveira, B.E.; Kawamoto, D.; Ando-Suguimoto, E.S.; Albuquerque-Souza, E.; Nicoli, J.R.; Ferreira, C.M.; de Lima, J.; et al. Bifidobacterium Strains Present Distinct Effects on the Control of Alveolar Bone Loss in a Periodontitis Experimental Model. Front. Pharmacol. 2021, 12, 713595. [Google Scholar] [CrossRef] [PubMed]
- Charan, J.; Kantharia, N.D. How to Calculate Sample Size in Animal Studies? J. Pharmacol. Pharmacother. 2013, 4, 303–306. [Google Scholar] [CrossRef]
- Barbosa, G.M.; Colombo, A.V.; Rodrigues, P.H.; Simionato, M.R.L. Intraspecies Variability Affects Heterotypic Biofilms of Porphyromonas gingivalis and Prevotella intermedia: Evidences of Strain-Dependence Biofilm Modulation by Physical Contact and by Released Soluble Factors. PLoS ONE 2015, 10, e0138687. [Google Scholar] [CrossRef]
- Rickard, A.H.; Palmer, R.J.; Blehert, D.S.; Campagna, S.R.; Semmelhack, M.F.; Egland, P.G.; Bassler, B.L.; Kolenbrander, P.E. Autoinducer 2: A Concentration-dependent Signal for Mutualistic Bacterial Biofilm Growth. Mol. Microbiol. 2006, 60, 1446–1456. [Google Scholar] [CrossRef] [PubMed]
- Pakula, R. Factors Regulating Competence in Transformation of Streptococci. J. Bacteriol. 1965, 90, 1320–1324. [Google Scholar] [CrossRef] [PubMed]
- Gatej, S.M.; Marino, V.; Bright, R.; Fitzsimmons, T.R.; Gully, N.; Zilm, P.; Gibson, R.J.; Edwards, S.; Bartold, P.M. Probiotic Lactobacillus rhamnosus GG Prevents Alveolar Bone Loss in a Mouse Model of Experimental Periodontitis. J. Clin. Periodontol. 2018, 45, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; de Brito Bezerra, B.; Pacios, S.; Andriankaja, O.; Li, Y.; Tsiagbe, V.; Schreiner, H.; Fine, D.H.; Graves, D.T. Aggregatibacter actinomycetemcomitans Infection Enhances Apoptosis In Vivo through a Caspase-3-Dependent Mechanism in Experimental Periodontitis. Infect. Immun. 2012, 80, 2247–2256. [Google Scholar] [CrossRef]
- Herlemann, D.P.; Labrenz, M.; Jürgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in Bacterial Communities along the 2000 Km Salinity Gradient of the Baltic Sea. ISME J. 2011, 5, 1571–1579. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.-R.; Shin, J.; Guevarra, R.B.; Lee, J.H.; Kim, D.W.; Seol, K.-H.; Lee, J.-H.; Kim, H.B.; Isaacson, R.E. Deciphering Diversity Indices for a Better Understanding of Microbial Communities. J. Microbiol. Biotechnol. 2017, 27, 2089–2093. [Google Scholar] [CrossRef]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An Effective Distance Metric for Microbial Community Comparison. ISME J. 2011, 5, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Van Treuren, W.; White, R.A.; Eggesbø, M.; Knight, R.; Peddada, S.D. Analysis of Composition of Microbiomes: A Novel Method for Studying Microbial Composition. Microb. Ecol. Health Dis. 2015, 26, 27663. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Peddada, S. Das Analysis of Compositions of Microbiomes with Bias Correction. Nat. Commun. 2020, 11, 3514. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G. Immunomicrobial Pathogenesis of Periodontitis: Keystones, Pathobionts, and Host Response. Trends Immunol. 2014, 35, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Payne, M.A.; Hashim, A.; Alsam, A.; Joseph, S.; Aduse-Opoku, J.; Wade, W.G.; Curtis, M.A. Horizontal and Vertical Transfer of Oral Microbial Dysbiosis and Periodontal Disease. J. Dent. Res. 2019, 98, 1503–1510. [Google Scholar] [CrossRef]
- Simas, A.M.; Kramer, C.D.; Weinberg, E.O.; Genco, C.A. Oral Infection with a Periodontal Pathogen Alters Oral and Gut Microbiomes. Anaerobe 2021, 71, 102399. [Google Scholar] [CrossRef]
- Tian, R.; Ning, D.; He, Z.; Zhang, P.; Spencer, S.J.; Gao, S.; Shi, W.; Wu, L.; Zhang, Y.; Yang, Y.; et al. Small and Mighty: Adaptation of Superphylum Patescibacteria to Groundwater Environment Drives Their Genome Simplicity. Microbiome 2020, 8, 51. [Google Scholar] [CrossRef]
- Lu, L.; Tang, M.; Li, J.; Xie, Y.; Li, Y.; Xie, J.; Zhou, L.; Liu, Y.; Yu, X. Gut Microbiota and Serum Metabolic Signatures of High-Fat-Induced Bone Loss in Mice. Front. Cell Infect. Microbiol. 2021, 11, 788576. [Google Scholar] [CrossRef] [PubMed]
- Tremlett, H.; Zhu, F.; Arnold, D.; Bar-Or, A.; Bernstein, C.N.; Bonner, C.; Forbes, J.D.; Graham, M.; Hart, J.; Knox, N.C.; et al. The Gut Microbiota in Pediatric Multiple Sclerosis and Demyelinating Syndromes. Ann. Clin. Transl. Neurol. 2021, 8, 2252–2269. [Google Scholar] [CrossRef]
- Wen, Y.; Feng, L.; Wang, H.; Zhou, H.; Li, Q.; Zhang, W.; Wang, M.; Li, Y.; Luan, X.; Jiang, Z.; et al. Association Between Oral Microbiota and Human Brain Glioma Grade: A Case-Control Study. Front. Microbiol. 2021, 12, 746568. [Google Scholar] [CrossRef]
- Luo, L.; Luo, J.; Cai, Y.; Fu, M.; Li, W.; Shi, L.; Liu, J.; Dong, R.; Xu, X.; Tu, L.; et al. Inulin-Type Fructans Change the Gut Microbiota and Prevent the Development of Diabetic Nephropathy. Pharmacol. Res. 2022, 183, 106367. [Google Scholar] [CrossRef]
- Wang, J.; Xiang, Q.; Gu, S.; Gu, Y.; Yao, M.; Huang, W.; Gao, W.; Tang, L.-L. Short- and Long-Term Effects of Different Antibiotics on the Gut Microbiota and Cytokines Level in Mice. Infect. Drug Resist. 2022, 15, 6785–6797. [Google Scholar] [CrossRef] [PubMed]
- Bull, M.; Plummer, S.; Marchesi, J.; Mahenthiralingam, E. The Life History of Lactobacillus acidophilus as a Probiotic: A Tale of Revisionary Taxonomy, Misidentification and Commercial Success. FEMS Microbiol. Lett. 2013, 349, 77–87. [Google Scholar] [CrossRef]
- Yli-Knuuttila, H.; Snäll, J.; Kari, K.; Meurman, J.H. Colonization of Lactobacillus rhamnosus GG in the Oral Cavity. Oral Microbiol. Immunol. 2006, 21, 129–131. [Google Scholar] [CrossRef]
- Collado, M.; Isolauri, E.; Salminen, S.; Sanz, Y. The Impact of Probiotic on Gut Health. Curr. Drug Metab. 2009, 10, 68–78. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cataruci, A.C.S.; Kawamoto, D.; Shimabukuro, N.; Ishikawa, K.H.; Ando-Suguimoto, E.S.; Ribeiro, R.A.; Nicastro, G.G.; Albuquerque-Souza, E.; de Souza, R.F.; Mayer, M.P.A. Oral Administration of Lactobacillus acidophilus LA5 Prevents Alveolar Bone Loss and Alters Oral and Gut Microbiomes in a Murine Periodontitis Experimental Model. Microorganisms 2024, 12, 1057. https://doi.org/10.3390/microorganisms12061057
Cataruci ACS, Kawamoto D, Shimabukuro N, Ishikawa KH, Ando-Suguimoto ES, Ribeiro RA, Nicastro GG, Albuquerque-Souza E, de Souza RF, Mayer MPA. Oral Administration of Lactobacillus acidophilus LA5 Prevents Alveolar Bone Loss and Alters Oral and Gut Microbiomes in a Murine Periodontitis Experimental Model. Microorganisms. 2024; 12(6):1057. https://doi.org/10.3390/microorganisms12061057
Chicago/Turabian StyleCataruci, Amalia C. S., Dione Kawamoto, Natali Shimabukuro, Karin H. Ishikawa, Ellen S. Ando-Suguimoto, Rodolfo A. Ribeiro, Gianlucca G. Nicastro, Emanuel Albuquerque-Souza, Robson F. de Souza, and Marcia P. A. Mayer. 2024. "Oral Administration of Lactobacillus acidophilus LA5 Prevents Alveolar Bone Loss and Alters Oral and Gut Microbiomes in a Murine Periodontitis Experimental Model" Microorganisms 12, no. 6: 1057. https://doi.org/10.3390/microorganisms12061057