

The Isolation and Characterization of Novel Caulobacter and Non-Caulobacter Lysogenic Bacteria from Soil and the Discovery of Broad-Host-Range Phages Infecting Multiple Genera



Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strain Isolation and Growth
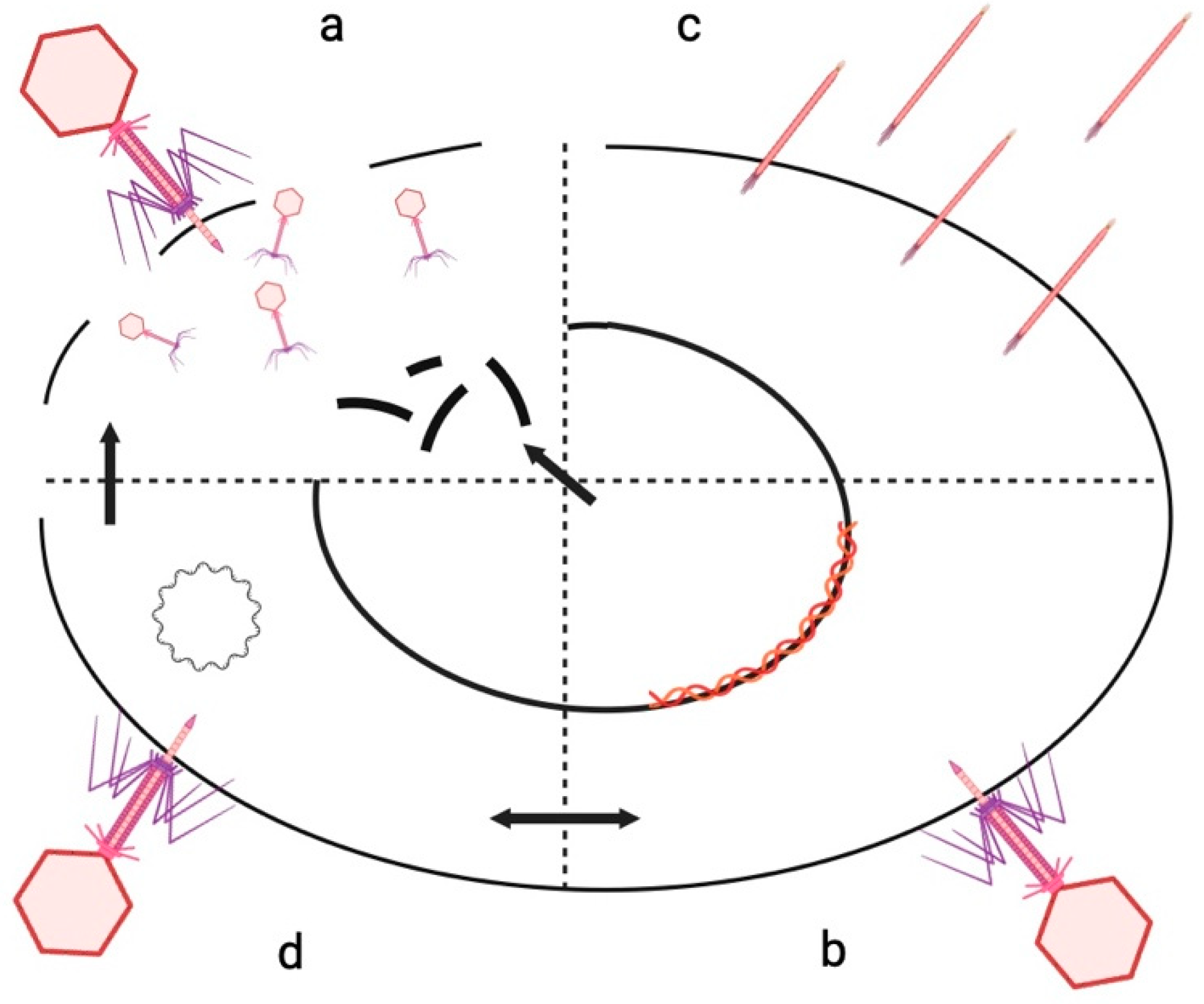
2.2. Lysogeny Test
2.3. Lysogenic Phage Induction with Mitomycin C
2.4. Host Range Determination
3. Results
3.1. Isolation and Characterization of Bacterial Strains from Soil and Plant Roots
3.2. Detection of Bacteriophages Capable of Lysing Caulobacter Strains
3.3. Characterization of Bacteriophage Host Ranges
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tsukanova, K.A.; Chebotar, V.K.; Meyer, J.J.M.; Bibikova, T.N. Effect of plant growth-promoting rhizobacteria on plant hormone homeostasis. S. Afr. J. Bot. 2017, 113, 91–102. [Google Scholar] [CrossRef]
- Richardson, A.E.; Barea, J.-M.; McNeill, A.M.; Prigent-Combaret, C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil 2009, 321, 305–339. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Vismans, G.; Yu, K.; Song, Y.; de Jonge, R.; Burgman, W.P.; Burmølle, M.; Herschend, J.; Bakker, P.A.H.M.; Pieterse, C.M.J. Disease-induced assemblage of a plant-beneficial bacterial consortium. ISME J. 2018, 12, 1496–1507. [Google Scholar] [CrossRef] [PubMed]
- Vacheron, J.; Desbrosses, G.; Bouffaud, M.-L.; Touraine, B.; Moënne-Loccoz, Y.; Muller, D.; Legendre, L.; Wisniewski-Dyé, F.; Prigent-Combaret, C. Plant growth-promoting rhizobacteria and root system functioning. Front. Plant. Sci 2013, 4, 356. [Google Scholar] [CrossRef] [PubMed]
- Gouda, S.; Kerry, R.G.; Das, G.; Paramithiotis, S.; Shin, H.-S.; Patra, J.K. Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiol. Res. 2018, 206, 131–140. [Google Scholar] [CrossRef]
- Luo, D.; Langendries, S.; Mendez, S.G.; De Ryck, J.; Liu, D.; Beirinckx, S.; Willems, A.; Russinova, E.; Debode, J.; Goormachtig, S. Plant growth promotion driven by a novel Caulobacter strain. Mol. Plant-Microbe Interact 2018, 10, 1162–1174. [Google Scholar] [CrossRef]
- Yang, E.; Sun, L.; Ding, X.; Sun, D.; Liu, J.; Wang, W. Complete genome sequence of Caulobacter flavus RHGG3 T, a type species of the genus Caulobacter with plant growth-promoting traits and heavy metal resistance. 3 Biotech 2019, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Agler, M.T.; Ruhe, J.; Kroll, S.; Morhenn, C.; Kim, S.T.; Weigel, D.; Kemen, E.M. Microbial hub taxa link host and abiotic factors to plant microbiome variation. PLoS Biol. 2016, 14, e1002352. [Google Scholar] [CrossRef]
- Poindexter, J.S. Biological properties and classification of the Caulobacter group. Bacteriol. Rev. 1964, 28, 231–295. [Google Scholar] [CrossRef]
- Wilhelm, R.C. Following the terrestrial tracks of Caulobacter—Redefining the ecology of a reputed aquatic oligotroph. ISME J. 2018, 12, 3025–3037. [Google Scholar] [CrossRef]
- Abraham, W.-R.; Strömpl, C.; Meyer, H.; Lindholst, S.; Moore, E.R.B.; Christ, R.; Vancanneyt, M.; Tindall, B.J.; Bennasar, A.; Smit, J.; et al. Phylogeny and polyphasic taxonomy of Caulobacter species. Proposal of Maricaulis gen. nov. with Maricaulis maris (Poindexter) comb. nov. as the type species, and emended description of the genera Brevundimonas and Caulobacter. Int. J. Syst. Bacteriol. 1999, 49, 1053–1073. [Google Scholar] [CrossRef] [PubMed]
- Reche, I.; D’Orta, G.; Mladenov, N.; Winget, D.M.; Suttle, C.A. Deposition rates of viruses and bacteria above the atmospheric boundary layer. ISME J. 2018, 12, 1154–1162. [Google Scholar] [CrossRef]
- Ely, B.; Lenski, J.; Mohammadi, T. Structural and genomic diversity of bacteriophages. In: Bacteriophages. Methods and Protocols. Methods Mol. Biol. 2024, 2738, 3–16. [Google Scholar] [PubMed]
- Ely, B.; Berrios, L.; Thomas, Q. S2B, a Temperate Bacteriophage That Infects Caulobacter Crescentus Strain CB15. Curr. Microbiol. 2022, 12, 98. [Google Scholar] [CrossRef] [PubMed]
- Paul, J.H. Prophages in marine bacteria: Dangerous molecular time bombs or the key to survival in the seas? ISME J. 2008, 2, 579. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann Berling, H.; Maze, R. Release of male-specific bacteriophages from surviving host bacteria. Virology 1964, 22, 305–313. [Google Scholar] [CrossRef]
- Ripp, S.; Miller, R.V. The role of pseudolysogeny in bacteriophage-host interactions in a natural freshwater environment. Microbiology 1997, 143, 2065–2070. [Google Scholar] [CrossRef]
- Ripp, S.; Miller, R.V. Dynamics of the pseudolysogenic response in slowly growing cells of Pseudomonas aeruginosa. Microbiology 1998, 144, 2225–2232. [Google Scholar] [CrossRef]
- Ackermann, H.W.; DuBow, M.S. Viruses of prokaryotes. In General Properties of Bacteriophages; CRC Press Inc.: Boca Raton, FL, USA, 1987; Volume 1, pp. 49–85. [Google Scholar]
- Weinbauer, M.G. Ecology of prokaryotic viruses. FEMS Microbiol. Rev. 2004, 28, 127–181. [Google Scholar] [CrossRef]
- Ross, A.; Ward, S.; Hyman, P. More is better: Selecting for broad host range bacteriophages. Front. Microbiol. 2016, 7, 1352. [Google Scholar] [CrossRef]
- Stewart, F.M.; Levin, B.R. The population biology of bacterial viruses—Why be temperate. Theor. Popul. Biol. 1984, 26, 93–117. [Google Scholar] [CrossRef] [PubMed]
- Marsh, P.; Wellington, E.M.H. Phage-host interactions in soil. FEMS Microbiol. Ecol. 1994, 15, 99–107. [Google Scholar] [CrossRef]
- Ely, B.; Johnson, R.C. Generalized transduction in Caulobacter crescentus. Genetics 1977, 87, 391–399. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Bacterial Name | Bacterial Strain | Colony Size | Pigmentation | Form | Elevation | Margin |
|---|---|---|---|---|---|---|
| RBW1 | Sphingomonas frigidaeris | Pinpoint | Yellow | Circular | Convex | Entire |
| RBW3 | Lysobacter | Large | Yellow | Irregular | Umbonate | Undulate |
| RBW6 | Pseudomonous | Large | White | Irregular | Flat | Undulate |
| RBW7 | Rhizobium | Large | White | Circular | Pulvinate | Entire |
| RBW11 | Sphingomonas sp. | Pinpoint | Yellow | Circular | Convex | Entire |
| RBW12 | Acidovorax sp. | Medium | White | Circular | Convex | Entire |
| RBW13 | Acidovorax sp. | Medium | White | Circular | Umbonate | Erose |
| RBW14 | Caulobacter rhizosphaerae | Large | White | Irregular | Umbonate | Undulate |
| RBW16 | Variovorax | Large | Yellow | Irregular | Umbonate | Undulate |
| RBW17 | Xanthomonas translucens | Large | Yellow | Irregular | Umbonate | Undulate |
| RBW18 | Brevundimonas | Large | Orange | Circular | Umbonate | Entire |
| RBW19 | Xanthomonas | Pinpoint | Yellow | Circular | Convex | Entire |
| RBW20 | Pseudomonous | Large | White | Irregular | Umbonate | Undulate |
| RBW21 | Caulobacter vibrioides | Large | White | Irregular | Umbonate | Undulate |
| RBW22 | Caulobacter vibrioides | Large | White | Circular | Umbonate | Undulate |
| RBW23 | Caulobacter rhizosphaerae | Large | Yellow | Irregular | Umbonate | Undulate |
| RBW25 | Caulobacter rhizosphaerae | Medium | Yellow | Circular | Convex | Entire |
| RBW26 | Caulobacter soil strain | Small | Yellow | Circular | Flat | Entire |
| RBW27 | Bacillus cereus | Large | White | Rhizoid | Umbonate | Lobate |
| RBW28 | Chromobacterium sp. | Large | White | Irregular | Umbonate | Undulate |
| RBW29 | Caulobacter vibrioides | Small | White | Circular | Convex | Entire |
| Lysogenic Bacteria | CB15 | CB13 |
|---|---|---|
| RBW1 | − | − |
| RBW3 | − | + |
| RBW6 | − | − |
| RBW7 | − | + |
| RBW11 | − | − |
| RBW12 | + | − |
| RBW13 | + | − |
| RBW14 | − | + |
| RBW16 | + | − |
| RBW17 | + | − |
| RBW18 | − | − |
| RBW19 | + | − |
| RBW20 | − | + |
| RBW21 | + | + |
| RBW22 | + | − |
| RBW23 | − | − |
| RBW25 | − | − |
| RBW26 | + | − |
| RBW27 | + | − |
| RBW28 | − | + |
| RBW29 | − | + |
| RBWs Bacteria | RBW1 | RBW6 | RBW11 | RBW18 | RBW23 | RBW25 |
|---|---|---|---|---|---|---|
| RBW1 | + | − | − | − | − | − |
| RBW3 | − | − | − | − | − | − |
| RBW6 | − | − | − | − | − | − |
| RBW7 | − | − | − | − | − | − |
| RBW11 | − | − | − | − | − | − |
| RBW12 | − | − | − | − | − | − |
| RBW13 | + | − | − | − | + | − |
| RBW14 | − | − | − | − | − | − |
| RBW16 | − | − | − | − | − | − |
| RBW17 | − | − | − | − | − | − |
| RBW18 | − | − | − | − | − | − |
| RBW19 | + | + | − | + | + | − |
| RBW20 | + | − | − | − | + | − |
| RBW21 | + | + | − | + | + | − |
| RBW22 | − | + | − | − | + | − |
| RBW23 | − | + | − | − | + | − |
| RBW25 | − | − | − | − | − | − |
| RBW26 | − | − | − | − | − | − |
| RBW27 | + | + | − | + | + | − |
| RBW28 | − | − | − | − | − | − |
| RBW29 | − | − | − | − | − | − |
| Bacteriophage Name | Original Host | Bacterial Strain | Plaque Morphology 1 | Positive Host Range | Self |
|---|---|---|---|---|---|
| RBC51 | RBW3 Lysobacter | CB13 | Turbid | CB15, CB13, CB2, RBW1, RBW14, RBW23 | − |
| RBC52 | RBW7 Rhizobium | CB13 | Turbid | CB15, CB13, CB2, RBW1, RBW13, RBW14, RBW22 | − |
| RBC53 | RBW12 Acidovorax | SC1004 | Turbid | CB15, CB13, CB2, FWC26, RBW1, RBW12, RBW14, RBW22, RBW23 | + |
| RBC54 | RBW12 Acidovorax | SC1004 | Clear | CB15, CB13, CB2, FWC26, RBW1, RBW12, RBW22, RBW23 | + |
| RBC55 | RBW13 Acidovorax | SC1004 | Turbid | CB15, CB13, CB2, CBR1, RBW14, RBW22 | − |
| RBC56 | RBW14 Caulobacter rhizosphaerae | CB13 | Turbid | CB15, CB13, CB2, FWC26, CBR1, RBW14, RBW12 | + |
| RBC57 | RBW16 Variovorax | SC1004 | Clear | CB15, CB13, CB2, CBR1, FWC26, TK0059, RBW12, RBW14, RBW22, RBW23, RBW26 | − |
| RBC58 | RBW16 Variovorax | SC1004 | Turbid | CB15, CB13, CB2, RBW12, RBW14, RBW22 | − |
| RBC59 | RBW17 Xanthomonas | SC1004 | Clear | CB15, CB13, CB2, TK0059, RBW12, RBW14, RBW22 | − |
| RBC60 | RBW19 Xanthomonas | SC1004 | Clear | CB15, CB13, CB2, CBR1, TK0059, RBW12, RBW14, RBW22, RBW23 | − |
| RBC61 | RBW20 Pseudomonas | SC1004 | Clear | CB15, CB13, CB2, RBW12, RBW14 | − |
| RBC62 | RBW21 Caulobacter vibrioides | SC1004 | Clear | CB15, CB13, CB2, RBW14 | − |
| RBC63 | RBW21 Caulobacter vibrioides | CB13 | Clear | CB15, CB13, CB2, FWC26, RBW12, RBW13, RBW14, RBW22 | − |
| RBC64 | RBW22 Caulobacter vibrioides | SC1004 | Clear | CB15, CB13, CB2, FWC26, RBW13, RBW14, RBW22 | + |
| RBC65 | RBW27 Bacillus cereus | SC1004 | Clear | CB15, CB13, CB2, RBW12, RBW13, RBW14, RBW22 | − |
| RBC66 | RBW28 Chromobacterium | CB13 | Turbid | CB15, CB13, CB2, FWC26, RBW1, RBW12, RBW13, RBW14, RBW22, RBW23 | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohammadi, T.; Ely, B. The Isolation and Characterization of Novel Caulobacter and Non-Caulobacter Lysogenic Bacteria from Soil and the Discovery of Broad-Host-Range Phages Infecting Multiple Genera. Microorganisms 2024, 12, 1894. https://doi.org/10.3390/microorganisms12091894
Mohammadi T, Ely B. The Isolation and Characterization of Novel Caulobacter and Non-Caulobacter Lysogenic Bacteria from Soil and the Discovery of Broad-Host-Range Phages Infecting Multiple Genera. Microorganisms. 2024; 12(9):1894. https://doi.org/10.3390/microorganisms12091894
Chicago/Turabian StyleMohammadi, Tannaz, and Bert Ely. 2024. "The Isolation and Characterization of Novel Caulobacter and Non-Caulobacter Lysogenic Bacteria from Soil and the Discovery of Broad-Host-Range Phages Infecting Multiple Genera" Microorganisms 12, no. 9: 1894. https://doi.org/10.3390/microorganisms12091894
APA StyleMohammadi, T., & Ely, B. (2024). The Isolation and Characterization of Novel Caulobacter and Non-Caulobacter Lysogenic Bacteria from Soil and the Discovery of Broad-Host-Range Phages Infecting Multiple Genera. Microorganisms, 12(9), 1894. https://doi.org/10.3390/microorganisms12091894