
Candidate-Probiotic Lactobacilli and Their Postbiotics as Health-Benefit Promoters
 , ,
, ,  , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Microorganisms, Cell Line, Media, and Culture Conditions
Cell Line, Media, and Conditions
2.2. Production of Postmetabolites (Cell-Free Supernatants, CFSs) and Postbiotics for In Vitro Tests
2.2.1. Preparation of EPS/EPS–Protein Crude Extract from LAB Spent Cultures
2.2.2. Protein Estimation of Produced Postbiotics
2.2.3. Lyophilization and Sample Preparation for Electron Paramagnetic Resonance (EPR) Analysis
2.3. In Vitro Antimicrobial Activity of LAB
2.3.1. Antibacterial Activity
2.3.2. Antifungal Activity
2.4. Free Radical Scavenging Ability Using DPPH and ABTS Methods
2.5. Electron Paramagnetic Resonance Spectroscopy Analysis (EPR)
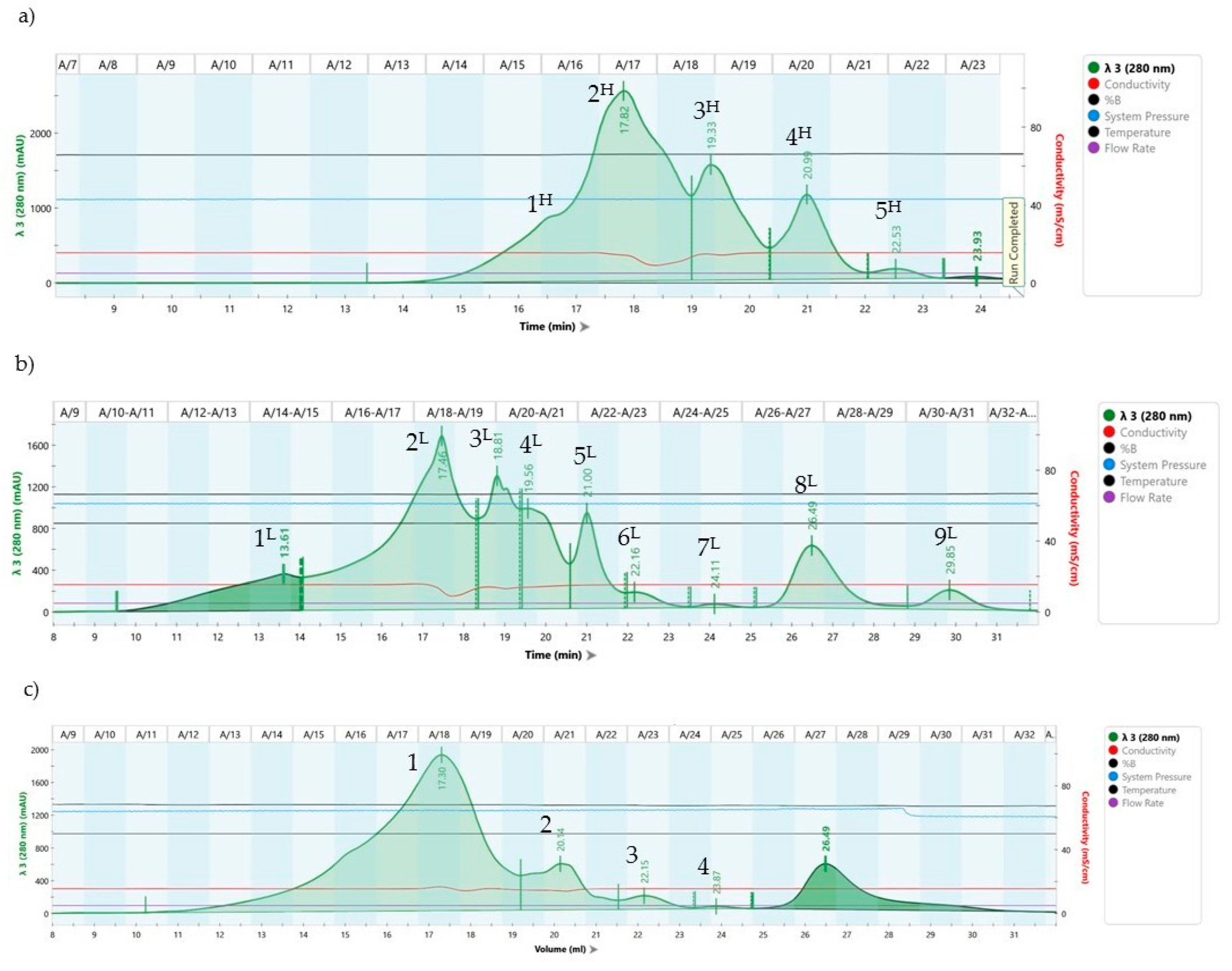
2.6. Fast Protein Liquid Chromatography (FPLC) Analysis
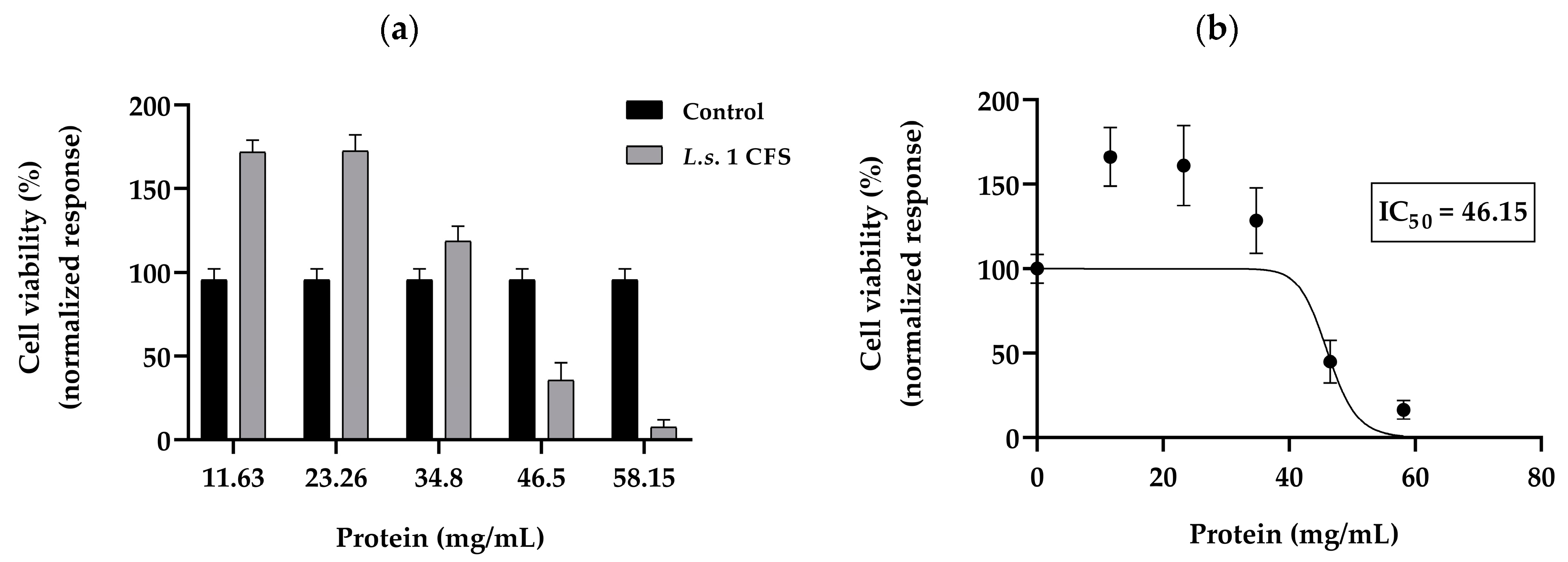
2.7. MTT Assay
2.8. Statistical Analysis
3. Results and Discussion
3.1. In Vitro Assessment of Four LAB Strains as Natural Antagonists of Different Pathogenic Bacteria and Molds
3.2. In Vitro Assessment of Antifungal Activity of LAB Strains
3.3. Radical Scavenging Activity of Metabiotics from LAB Cultures
3.4. EPR Analysis of Ready-to-Use Probiotics from Two Selected Strains
3.5. FPLC Chromatography Analyses
3.6. Inhibitory Effects of Postbiotics of LAB Cultures on the Growth of K562 Cell Lines Using the MTT Assay
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shenderov, B.A. Metabiotics: Novel idea or natural development of probiotic conception. Microb. Ecol. Health Dis. 2013, 24, 20399. [Google Scholar] [CrossRef] [PubMed]
- Moradi, M.; Kousheh, S.A.; Almasi, H.; Alizadeh, A.; Guimarães, J.T.; Yılmaz, N.; Lotfi, A. Postbiotics produced by lactic acid bacteria: The next frontier in food safety. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3390–3415. [Google Scholar] [CrossRef]
- Dobreva, L.; Borisova, D.; Paunova-Krasteva, T.; Dimitrova, P.D.; Hubenov, V.; Atanasova, N.; Ivanov, I.; Danova, S. From traditional dairy product “katak” to beneficial lactiplantibacillus plantarum strains. Microorganisms 2023, 11, 2847. [Google Scholar] [CrossRef] [PubMed]
- Tremonte, P.; Pannella, G.; Succi, M.; Luca, T.; Sturchio, M.; Coppola, R.; Luongo, D.; Sorrentino, E. Antimicrobial activity of Lactobacillus plantarum strains isolated from different environments: A preliminary study. Int. Food Res. J. 2017, 24, 852–859. [Google Scholar]
- Chen, H.; Yan, X.; Du, G.; Guo, Q.; Shi, Y.; Chang, J.; Wang, X.; Yuan, Y.; Yue, T. Recent developments in antifungal lactic acid bacteria: Application, screening methods, separation, purification of antifungal compounds and antifungal mechanisms. Crit. Rev. Food Sci. Nutr. 2023, 63, 2544–2558. [Google Scholar] [CrossRef]
- Ponzio, A.; Rebecchi, A.; Zivoli, R.; Morelli, L. Reuterin, phenyllactic acid, and exopolysaccharides as main antifungal molecules produced by lactic acid bacteria: A scoping review. Foods 2024, 13, 752. [Google Scholar] [CrossRef] [PubMed]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The international scientific association of probiotics and prebiotics (isapp) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Tuohy, K.M.; Scott, K.P. The microbiota of the human gastrointestinal tract: A molecular view. In Diet-Microbe Interactions in the Gut; Elsevier: Amsterdam, The Netherlands, 2015; pp. 1–15. [Google Scholar]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Núñez, G. Gut microbiota: Role in pathogen colonization, immune responses, and inflammatory disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef]
- dos Reis, S.A.; da Conceição, L.L.; Siqueira, N.P.; Rosa, D.D.; da Silva, L.L.; Peluzio, M.d.C.G. Review of the mechanisms of probiotic actions in the prevention of colorectal cancer. Nutr. Res. 2017, 37, 1–19. [Google Scholar] [CrossRef]
- Huynh, U.; Zastrow, M.L. Metallobiology of lactobacillaceae in the gut microbiome. J. Inorg. Biochem. 2023, 238, 112023. [Google Scholar] [CrossRef]
- Lin, C.; Cai, X.; Zhang, J.; Wang, W.; Sheng, Q.; Hua, H.; Zhou, X. Role of gut microbiota in the development and treatment of colorectal cancer. Digestion 2018, 100, 72–78. [Google Scholar] [CrossRef]
- Muegge, B.D.; Kuczynski, J.; Knights, D.; Clemente, J.C.; González, A.; Fontana, L.; Henrissat, B.; Knight, R.; Gordon, J.I. Diet drives convergence in gut microbiome functions across mammalian phylogeny and within humans. Science 2011, 332, 970–974. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Chang, H.-W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef]
- Vilhelmova-Ilieva, N.; Atanasov, G.; Simeonova, L.; Dobreva, L.; Mancheva, K.; Trepechova, M.; Danova, S. Anti-herpes virus activity of lactobacillus’ postbiotics. BioMedicine 2022, 12, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Das, T.K.; Pradhan, S.; Chakrabarti, S.; Mondal, K.C.; Ghosh, K. Current status of probiotic and related health benefits. Appl. Food Res. 2022, 2, 100185. [Google Scholar] [CrossRef]
- Divyashree, S.; Shruthi, B.; Vanitha, P.R.; Sreenivasa, M.Y. Probiotics and their postbiotics for the control of opportunistic fungal pathogens: A review. Biotechnol. Rep. 2023, 38, e00800. [Google Scholar] [CrossRef]
- World Health Organization. The Global Burden of Disease: 2004 Update; World Health Organisation: Geneva, Switzerland, 2008; Available online: https://www.who.int/publications/i/item/9789241563710 (accessed on 16 September 2024).
- Wild, C.P. The global cancer burden: Necessity is the mother of prevention. Nat. Rev. Cancer 2019, 19, 123–124. [Google Scholar] [CrossRef]
- National Cancer Institute. Leukemia. Available online: https://seer.cancer.gov/statfacts/html/leuks.html (accessed on 16 September 2024).
- Panebianco, C.; Latiano, T.; Pazienza, V. Microbiota manipulation by probiotics administration as emerging tool in cancer prevention and therapy. Front. Oncol. 2020, 10, 679. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J. 1984, 219, 1–14. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef]
- Kahouli, I.; Tomaro-Duchesneau, C.; Prakash, S. Probiotics in colorectal cancer (crc) with emphasis on mechanisms of action and current perspectives. J. Med. Microbiol. 2013, 62, 1107–1123. [Google Scholar] [CrossRef] [PubMed]
- Haghshenas, B.; Abdullah, N.; Nami, Y.; Radiah, D.; Rosli, R.; Khosroushahi, A.Y. Different effects of two newly-isolated probiotic Lactobacillus plantarum 15hn and Lactococcus lactis subsp. Lactis 44lac strains from traditional dairy products on cancer cell lines. Anaerobe 2014, 30, 51–59. [Google Scholar] [CrossRef] [PubMed]
- El-Deeb, N.M.; Yassin, A.M.; Al-Madboly, L.A.; El-Hawiet, A. A novel purified lactobacillus acidophilus 20079 exopolysaccharide, la-eps-20079, molecularly regulates both apoptotic and nf-κb inflammatory pathways in human colon cancer. Microb. Cell Factories 2018, 17, 29. [Google Scholar] [CrossRef]
- Górska, A.; Przystupski, D.; Niemczura, M.J.; Kulbacka, J. Probiotic bacteria: A promising tool in cancer prevention and therapy. Curr. Microbiol. 2019, 76, 939–949. [Google Scholar] [CrossRef]
- Hu, J.; Chen, F.; Kan, T.; Zhuang, H.; Zhang, J.; Han, X. Inhibition of fusarium solani infection in murine keratocytes by lactobacillus salivarius ssp. Salivarius jcm1231 culture filtrate in vitro. Curr. Eye Res. 2017, 42, 1339–1347. [Google Scholar] [CrossRef] [PubMed]
- Escamilla, J.; Lane, M.A.; Maitin, V. Cell-free supernatants from probiotic Lactobacillus casei and Lactobacillus rhamnosus gg decrease colon cancer cell invasion in vitro. Nutr. Cancer 2012, 64, 871–878. [Google Scholar] [CrossRef]
- Dobreva, L. Functional Characteristic of Lactic Acid Bacteria from Different Habitats. Ph.D. Dissertation, The Stephan Angeloff Institute of Microbiology, Bulgarian Academy of Sciences, Sofia, Bulgaria, 2023. [Google Scholar]
- Danova, S. Biodiversity and Probiotic Potential of Lactic Acid Bacteria from Different Ecological Niches. Ph.D. Thesis, The Stephan Angeloff Institute of Microbiology, Bulgarian Academy of Sciences, Sofia, Bulgaria, 2015. [Google Scholar]
- Schuelter, A.R.; Kroumov, A.D.; Hinterholz, C.L.; Fiorini, A.; Trigueros, D.E.G.; Vendruscolo, E.G.; Zaharieva, M.M.; Módenes, A.N. Isolation and identification of new microalgae strains with antibacterial activity on food-borne pathogens. Engineering approach to optimize synthesis of desired metabolites. Biochem. Eng. J. 2019, 144, 28–39. [Google Scholar] [CrossRef]
- Todorov, S.D.; Vaz-Velho, M.; de Melo Franco, B.D.G.; Holzapfel, W.H. Partial characterization of bacteriocins produced by three strains of lactobacillus sakei, isolated from salpicao, a fermented meat product from north-west of portugal. Food Control 2013, 30, 111–121. [Google Scholar] [CrossRef]
- Tropcheva, R.; Nikolova, D.; Evstatieva, Y.; Danova, S. Antifungal activity and identification of lactobacilli, isolated from traditional dairy product “katak”. Anaerobe 2014, 28, 78–84. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved abts radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. The World Health Report 2002: Reducing Risks, Promoting Healthy Life; World Health Organization: Geneva, Switzerland, 2002. [Google Scholar]
- Dobreva, L.; Danova, S.; Georgieva, V.; Georgieva, S.; Koprinarova, M. Anti-salmonella activity of lactobacilli from different habitats. Bulg. J. Vet. Med. 2020, 25, 2020. [Google Scholar] [CrossRef]
- Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Gancel, F.; Kempf, I.; Drider, D. Benefits and inputs from lactic acid bacteria and their bacteriocins as alternatives to antibiotic growth promoters during food-animal production. Front. Microbiol. 2019, 10, 57. [Google Scholar] [CrossRef] [PubMed]
- Gherardi, G. Staphylococcus aureus infection: Pathogenesis and antimicrobial resistance. Int. J. Mol. Sci. 2023, 24, 8182. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.F.; Nyachoti, M. Using probiotics to improve swine gut health and nutrient utilization. Anim. Nutr. (Zhongguo Xu Mu Shou Yi Xue Hui) 2017, 3, 331–343. [Google Scholar] [CrossRef]
- Sulijaya, B.; Takahashi, N.; Yamazaki, K. Lactobacillus-derived bioactive metabolites for the regulation of periodontal health: Evidences to clinical setting. Molecules 2020, 25, 2088. [Google Scholar] [CrossRef]
- Kang, M.-S.; Lim, H.-S.; Oh, J.-S.; Lim, Y.-j.; Wuertz-Kozak, K.; Harro, J.M.; Shirtliff, M.E.; Achermann, Y. Antimicrobial activity of Lactobacillus salivarius and Lactobacillus fermentum against Staphylococcus aureus. Pathog. Dis. 2017, 75, ftx009. [Google Scholar] [CrossRef]
- Chen, Y.T.; Hsieh, P.S.; Ho, H.H.; Hsieh, S.H.; Kuo, Y.W.; Yang, S.F.; Lin, C.W. Antibacterial activity of viable and heat-killed probiotic strains against oral pathogens. Lett. Appl. Microbiol. 2020, 70, 310–317. [Google Scholar] [CrossRef]
- Layus, B.I.; Gerez, C.L.; Rodriguez, A.V. Antibacterial activity of Lactobacillus plantarum crl 759 against methicillin-resistant Staphylococcus aureus and Pseudomonas aeruginosa. Arab. J. Sci. Eng. 2020, 45, 4503–4510. [Google Scholar] [CrossRef]
- Mao, Y.; Zhang, X.; Xu, Z. Identification of antibacterial substances of Lactobacillus plantarum dy-6 for bacteriostatic action. Food Sci. Nutr. 2020, 8, 2854–2863. [Google Scholar] [CrossRef]
- Lim, S.-M.; Lee, N.-K.; Paik, H.-D. Antibacterial and anticavity activity of probiotic Lactobacillus plantarum 200661 isolated from fermented foods against Streptococcus mutans. LWT 2020, 118, 108840. [Google Scholar] [CrossRef]
- Younes, Z.; Cheriguene, A.; Chougrani, F.; Merzouk, Y.; Marchetta, A.; Urzì, C.; De Leo, F. Antifungal activity of lactic acid bacteria against phytopathogenic alternaria alternata species and their molecular characterization. J. Food Nutr. Res. 2021, 60, 18–28. [Google Scholar]
- Maschmeyer, G.; Haas, A.; Cornely, O.A. Invasive aspergillosis. Drugs 2007, 67, 1567–1601. [Google Scholar] [CrossRef]
- van Diepeningen, A.D.; Brankovics, B.; Iltes, J.; van der Lee, T.A.J.; Waalwijk, C. Diagnosis of fusarium infections: Approaches to identification by the clinical mycology laboratory. Curr. Fungal Infect. Rep. 2015, 9, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zeng, X.; Fu, H.; Wang, X.; Guo, X.; Wang, M. Lactiplantibacillus plantarum: A comprehensive review of its antifungal and anti-mycotoxic effects. Trends Food Sci. Technol. 2023, 136, 224–238. [Google Scholar] [CrossRef]
- Crowley, S.; Mahony, J.; Van Sinderen, D. Current perspectives on antifungal lactic acid bacteria as natural bio-preservatives. Trends Food Sci. Technol. 2013, 33, 93–109. [Google Scholar] [CrossRef]
- Dong, A.-R.; Thuy Ho, V.T.; Lo, R.; Bansal, N.; Turner, M.S. A genetic diversity study of antifungal Lactobacillus plantarum isolates. Food Control 2017, 72, 83–89. [Google Scholar] [CrossRef]
- Russo, P.; Arena, M.P.; Fiocco, D.; Capozzi, V.; Drider, D.; Spano, G. Lactobacillus plantarum with broad antifungal activity: A promising approach to increase safety and shelf-life of cereal-based products. Int. J. Food Microbiol. 2017, 247, 48–54. [Google Scholar] [CrossRef]
- Vazquez-Munoz, R.; Dongari-Bagtzoglou, A. Anticandidal activities by lactobacillus species: An update on mechanisms of action. Front. Oral Health 2021, 2, 689382. [Google Scholar] [CrossRef]
- van Niel, E.W.; Hofvendahl, K.; Hahn-Hägerdal, B. Formation and conversion of oxygen metabolites by Lactococcus lactis subsp. Lactis atcc 19435 under different growth conditions. Appl. Environ. Microbiol. 2002, 68, 4350–4356. [Google Scholar] [CrossRef]
- Sánchez-Moreno, C. Methods used to evaluate the free radical scavenging activity in foods and biological systems. Food Sci. Technol. Int. 2002, 8, 121–137. [Google Scholar] [CrossRef]
- Elias, R.J.; Kellerby, S.S.; Decker, E.A. Antioxidant activity of proteins and peptides. Crit. Rev. Food Sci. Nutr. 2008, 48, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Kullisaar, T.; Zilmer, M.; Mikelsaar, M.; Vihalemm, T.; Annuk, H.; Kairane, C.; Kilk, A. Two antioxidative lactobacilli strains as promising probiotics. Int. J. Food Microbiol. 2002, 72, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Noureen, S.; Riaz, A.; Arshad, M.; Arshad, N. In vitro selection and in vivo confirmation of the antioxidant ability of lactobacillus brevis mg000874. J. Appl. Microbiol. 2019, 126, 1221–1232. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Wang, J. Oxidative stress tolerance and antioxidant capacity of lactic acid bacteria as probiotic: A systematic review. Gut Microbes 2020, 12, 1801944. [Google Scholar] [CrossRef]
- Tang, W.; Xing, Z.; Li, C.; Wang, J.; Wang, Y.J.F.C. Molecular mechanisms and in vitro antioxidant effects of Lactobacillus plantarum ma2. Food Chem. 2017, 221, 1642–1649. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, C.; Li, D.; Zhao, Y.; Zhang, X.; Zeng, X.; Yang, Z.; Li, S. Antioxidant activity of an exopolysaccharide isolated from Lactobacillus plantarum c88. Int. J. Biol. Macromol. 2013, 54, 270–275. [Google Scholar] [CrossRef]
- Eladawi, H.; Khalil, M.; El-Sheekh, M.; El-Deeb, N.; Hussein, M. Cytotoxicity assay and antioxidant activities of the lactic acid bacterial strains. Afr. J. Microbiol. Res. 2012, 6, 1700–1712. [Google Scholar]
- Chooruk, A.; Piwat, S.; Teanpaisan, R. Antioxidant activity of various oral lactobacillus strains. J. Appl. Microbiol. 2017, 123, 271–279. [Google Scholar] [CrossRef]
- Ding, W.; Wang, L.; Zhang, J.; Ke, W.; Zhou, J.; Zhu, J.; Guo, X.; Long, R. Characterization of antioxidant properties of lactic acid bacteria isolated from spontaneously fermented yak milk in the tibetan plateau. J. Funct. Foods 2017, 35, 481–488. [Google Scholar] [CrossRef]
- Bercu, V.; Negut, C.D.; Duliu, O.G. Epr investigation of some desiccated ascomycota and basidiomycota gamma-irradiated mushrooms. Radiat. Phys. Chem. 2010, 79, 1275–1278. [Google Scholar] [CrossRef]
- Brückner, A.; Bentrup, U.; Zanthoff, H.; Maschmeyer, D. The role of different ni sites in supported nickel catalysts for butene dimerization under industry-like conditions. J. Catal. 2009, 266, 120–128. [Google Scholar] [CrossRef]
- Bonneviot, L.; Che, M.; Dyrek, K.; Schollner, R.; Wendt, G. An epr study of the formation of nickel(1+) ions by photoreduction in hydrogen of nickel/alumina catalysts. J. Phys. Chem. 1986, 90, 2379–2384. [Google Scholar] [CrossRef]
- Poddar, D.; de Jonge, M.D.; Howard, D.L.; Palmer, J.; Ainscough, E.W.; Singh, H.; Haverkamp, R.G.; Jameson, G.B. Manganese accumulation in probiotic Lactobacillus paracasei atcc 55544 analyzed by synchrotron X-ray fluorescence microscopy and impact of accumulation on the bacterial viability following encapsulation. Food Res. Int. 2021, 147, 110528. [Google Scholar] [CrossRef]
- Archibald, F.S.; Fridovich, I. Manganese and defenses against oxygen toxicity in Lactobacillus plantarum. J. Bacteriol. 1981, 145, 442–451. [Google Scholar] [CrossRef]
- Lew, L.C.; Liong, M.T.; Gan, C.Y. Growth optimization of Lactobacillus rhamnosus ftdc 8313 and the production of putative dermal bioactives in the presence of manganese and magnesium ions. J. Appl. Microbiol. 2013, 114, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.; Zhai, Q.; Wang, G.; Zhang, Q.; Liu, X.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. System-wide analysis of manganese starvation-induced metabolism in key elements of Lactobacillus plantarum. RSC Adv. 2017, 7, 12959–12968. [Google Scholar] [CrossRef]
- Carman, M.D.; Schornagel, J.H.; Rivest, R.S.; Srimatkandada, S.; Portlock, C.S.; Duffy, T.; Bertino, J.R. Resistance to methotrexate due to gene amplification in a patient with acute leukemia. J. Clin. Oncol. 1984, 2, 16–20. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, T.; Du, J.; Li, Y.; Wang, X.; Zhou, Y.; Yu, X.; Fan, W.; Zhu, Q.; Tong, X.; et al. The critical role of pten/pi3k/akt signaling pathway in shikonin-induced apoptosis and proliferation inhibition of chronic myeloid leukemia. Cell. Physiol. Biochem. 2018, 47, 981–993. [Google Scholar] [CrossRef] [PubMed]
- Maghsood, F.; Johari, B.; Rohani, M.; Madanchi, H.; Saltanatpour, Z.; Kadivar, M. Anti-proliferative and anti-metastatic potential of high molecular weight secretory molecules from probiotic Lactobacillus reuteri cell-free supernatant against human colon cancer stem-like cells (ht29-she). Int. J. Pept. Res. Ther. 2020, 26, 2619–2631. [Google Scholar] [CrossRef]
- Kim, J.-H.; Kim, D.-M.; Baek, H.; Lee, S.-H.; Chung, M.-J. Anti-cancer effects of peptides purified from culture supernatant of Lactobacillus casei. J. Dairy Sci. Biotechnol. 2008, 26, 5–10. [Google Scholar]
- Chuah, L.O.; Foo, H.L.; Loh, T.C.; Mohammed Alitheen, N.B.; Yeap, S.K.; Abdul Mutalib, N.E.; Abdul Rahim, R.; Yusoff, K. Postbiotic metabolites produced by Lactobacillus plantarum strains exert selective cytotoxicity effects on cancer cells. BMC Complement. Altern. Med. 2019, 19, 114. [Google Scholar] [CrossRef] [PubMed]
- Tuo, Y.; Jiang, S.; Qian, F.; Mu, G.; Liu, P.; Guo, Y.; Ma, C. Short communication: Antiproliferative effect of 8 different lactobacillus strains on k562 cells. J. Dairy Sci. 2015, 98, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Pagani, I.S.; Poudel, G.; Wardill, H.R. A gut instinct on leukaemia: A new mechanistic hypothesis for microbiota-immune crosstalk in disease progression and relapse. Microorganisms 2022, 10, 713. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dobreva, L.; Atanasova, N.; Donchev, P.; Krumova, E.; Abrashev, R.; Karakirova, Y.; Mladenova, R.; Tolchkov, V.; Ralchev, N.; Dishliyska, V.; et al. Candidate-Probiotic Lactobacilli and Their Postbiotics as Health-Benefit Promoters. Microorganisms 2024, 12, 1910. https://doi.org/10.3390/microorganisms12091910
Dobreva L, Atanasova N, Donchev P, Krumova E, Abrashev R, Karakirova Y, Mladenova R, Tolchkov V, Ralchev N, Dishliyska V, et al. Candidate-Probiotic Lactobacilli and Their Postbiotics as Health-Benefit Promoters. Microorganisms. 2024; 12(9):1910. https://doi.org/10.3390/microorganisms12091910
Chicago/Turabian StyleDobreva, Lili, Nikoleta Atanasova, Petar Donchev, Ekaterina Krumova, Radoslav Abrashev, Yordanka Karakirova, Ralitsa Mladenova, Vladimir Tolchkov, Nikola Ralchev, Vladislava Dishliyska, and et al. 2024. "Candidate-Probiotic Lactobacilli and Their Postbiotics as Health-Benefit Promoters" Microorganisms 12, no. 9: 1910. https://doi.org/10.3390/microorganisms12091910
APA StyleDobreva, L., Atanasova, N., Donchev, P., Krumova, E., Abrashev, R., Karakirova, Y., Mladenova, R., Tolchkov, V., Ralchev, N., Dishliyska, V., & Danova, S. (2024). Candidate-Probiotic Lactobacilli and Their Postbiotics as Health-Benefit Promoters. Microorganisms, 12(9), 1910. https://doi.org/10.3390/microorganisms12091910