
Reversal of Polymicrobial Biofilm Tolerance to Ciprofloxacin by Blue Light plus Carvacrol


Abstract
:1. Introduction
2. Materials and Methods
2.1. Light Source, Compounds, and Microorganisms
2.2. Anti-Planktonic Bacteria Assay
2.3. Polymicrobial Biofilm
2.4. Quantitative PCR (qPCR) to Measure Biofilm Bacterial Composition
2.5. Anti-Biofilm Assays
2.6. Measure Bacterial Membrane Permeability
2.7. Statistical Analysis
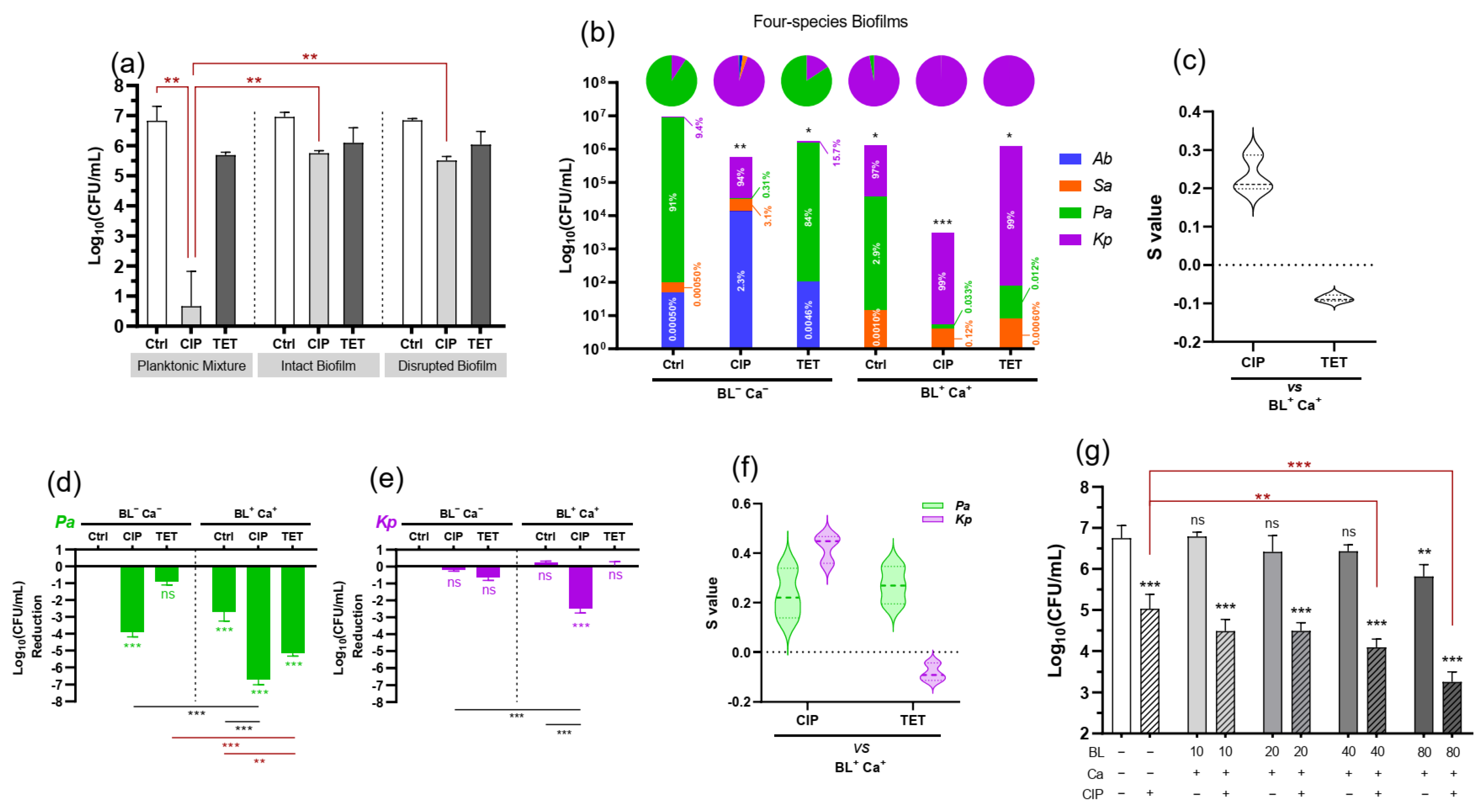
3. Results
3.1. P. aeruginosa Dominate the Five-Species Polymicrobial Biofilm
3.2. Polymicrobial Biofilms Tolerate Ciprofloxacin
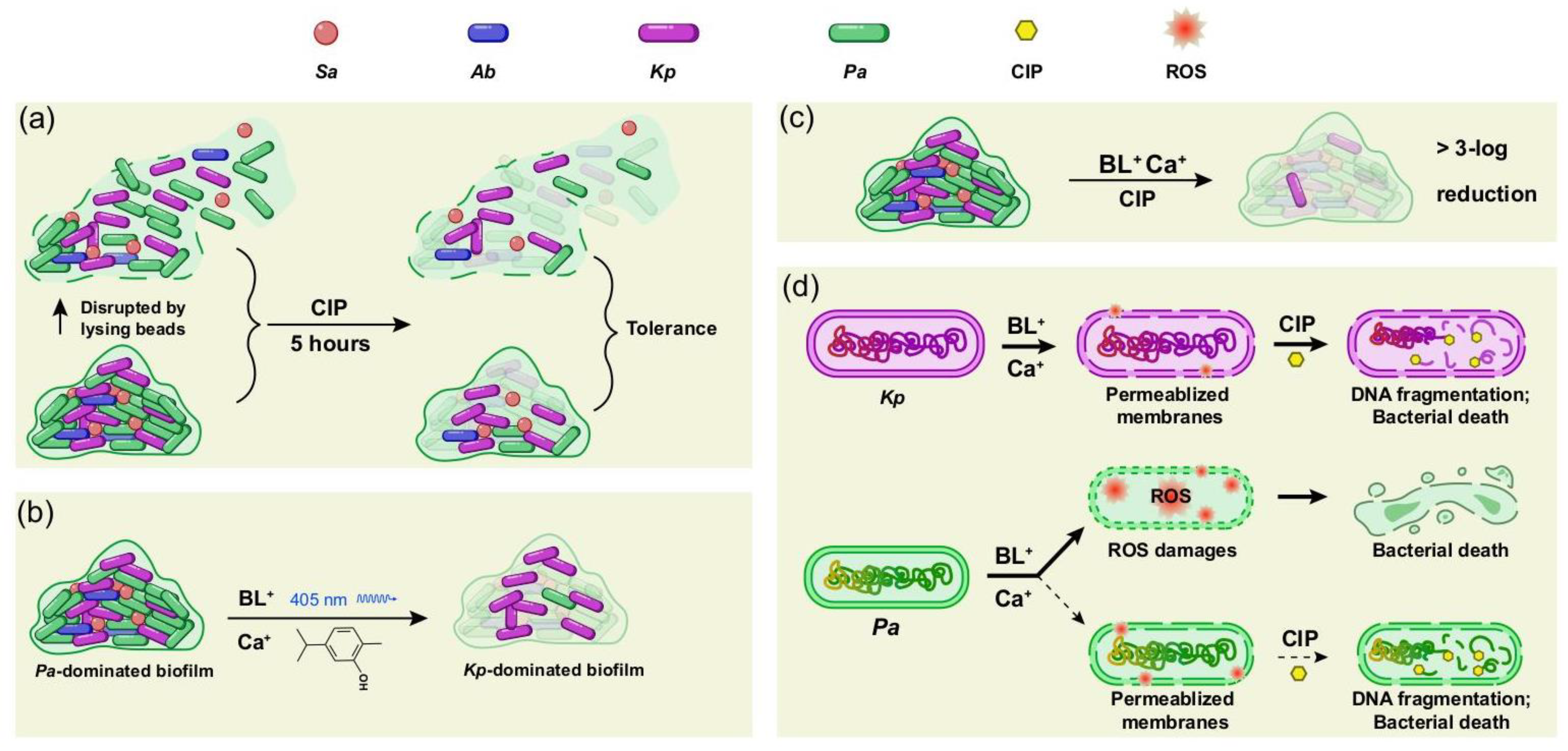
3.3. BL-Ca Dual Treatment Potentiates Anti-Biofilm Activities of Ciprofloxacin and Reshapes the Biofilm Community
3.4. Biofilm-Associated P. aeruginosa or K. pneumoniae Respond Differently to BL-Ca Treatment
3.5. BL-Ca Treatment Permeabilizes Bacterial Membranes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Stewart, P.S.; Costerton, J.W. Antibiotic resistance of bacteria in biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Grant, S.S.; Hung, D.T. Persistent bacterial infections, antibiotic tolerance, and the oxidative stress response. Virulence 2013, 4, 273–283. [Google Scholar] [CrossRef] [Green Version]
- Hall, C.W.; Mah, T.-F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef]
- Shigeta, M.; Tanaka, G.; Komatsuzawa, H.; Sugai, M.; Suginaka, H.; Usui, T. Permeation of antimicrobial agents through Pseudomonas aeruginosa biofilms: A simple method. Chemotherapy 1997, 43, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K. Multidrug tolerance of biofilms and persister cells. Bact. Biofilms 2008, 322, 107–131. [Google Scholar]
- Jensen, P.Ø.; Briales, A.; Brochmann, R.P.; Wang, H.; Kragh, K.N.; Kolpen, M.; Hempel, C.; Bjarnsholt, T.; Høiby, N.; Ciofu, O. Formation of hydroxyl radicals contributes to the bactericidal activity of ciprofloxacin against Pseudomonas aeruginosa biofilms. Pathog. Dis. 2014, 70, 440–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reza, A.; Sutton, J.M.; Rahman, K.M. Effectiveness of efflux pump inhibitors as biofilm disruptors and resistance breakers in gram-negative (ESKAPEE) bacteria. Antibiotics 2019, 8, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, K. Riddle of biofilm resistance. Antimicrob. Agents Chemother. 2001, 45, 999–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harriott, M.M.; Noverr, M.C. Importance of Candida–bacterial polymicrobial biofilms in disease. Trends Microbiol. 2011, 19, 557–563. [Google Scholar] [CrossRef] [Green Version]
- Elias, S.; Banin, E. Multi-species biofilms: Living with friendly neighbors. FEMS Microbiol. Rev. 2012, 36, 990–1004. [Google Scholar] [CrossRef]
- Wolcott, R.; Costerton, J.; Raoult, D.; Cutler, S. The polymicrobial nature of biofilm infection. Clin. Microbiol. Infect. 2013, 19, 107–112. [Google Scholar] [CrossRef] [Green Version]
- James, G.A.; Swogger, E.; Wolcott, R.; Pulcini, E.d.; Secor, P.; Sestrich, J.; Costerton, J.W.; Stewart, P.S. Biofilms in chronic wounds. Wound Repair Regen. 2008, 16, 37–44. [Google Scholar] [CrossRef]
- Dalton, T.; Dowd, S.E.; Wolcott, R.D.; Sun, Y.; Watters, C.; Griswold, J.A.; Rumbaugh, K.P. An in vivo polymicrobial biofilm wound infection model to study interspecies interactions. PLoS ONE 2011, 6, e27317. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, L.R.; Déziel, E.; d’Argenio, D.A.; Lépine, F.; Emerson, J.; McNamara, S.; Gibson, R.L.; Ramsey, B.W.; Miller, S.I. Selection for Staphylococcus aureus small-colony variants due to growth in the presence of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2006, 103, 19890–19895. [Google Scholar] [CrossRef] [Green Version]
- Lebeaux, D.; Chauhan, A.; Rendueles, O.; Beloin, C. From in vitro to in vivo models of bacterial biofilm-related infections. Pathogens 2013, 2, 288–356. [Google Scholar] [CrossRef] [Green Version]
- Ferrer-Espada, R.; Liu, X.; Goh, X.S.; Dai, T. Antimicrobial blue light inactivation of polymicrobial biofilms. Front. Microbiol. 2019, 10, 721. [Google Scholar] [CrossRef]
- Woods, J.; Boegli, L.; Kirker, K.R.; Agostinho, A.M.; Durch, A.M.; Delancey Pulcini, E.; Stewart, P.S.; James, G.A. Development and application of a polymicrobial, in vitro, wound biofilm model. J. Appl. Microbiol. 2012, 112, 998–1006. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Dowd, S.E.; Smith, E.; Rhoads, D.D.; Wolcott, R.D. In vitro multispecies Lubbock chronic wound biofilm model. Wound Repair Regen. 2008, 16, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Smith, E.; Wolcott, R.; Dowd, S. Propagation of anaerobic bacteria within an aerobic multi-species chronic wound biofilm model. J. Wound Care 2009, 18, 426–431. [Google Scholar] [CrossRef] [PubMed]
- DeLeon, S.; Clinton, A.; Fowler, H.; Everett, J.; Horswill, A.R.; Rumbaugh, K.P. Synergistic interactions of Pseudomonas aeruginosa and Staphylococcus aureus in an in vitro wound model. Infect. Immun. 2014, 82, 4718–4728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, C.N.; Davies, D.G.; Sauer, K. Control of biofilms with the fatty acid signaling molecule cis-2-decenoic acid. Pharmaceuticals 2015, 8, 816–835. [Google Scholar] [CrossRef]
- Bjarnsholt, T.; Ciofu, O.; Molin, S.; Givskov, M.; Høiby, N. Applying insights from biofilm biology to drug development—Can a new approach be developed? Nat. Rev. Drug Discov. 2013, 12, 791–808. [Google Scholar] [CrossRef]
- Sultana, S.T.; Call, D.R.; Beyenal, H. Eradication of Pseudomonas aeruginosa biofilms and persister cells using an electrochemical scaffold and enhanced antibiotic susceptibility. NPJ Biofilms Microbiomes 2016, 2, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Wang, S.; Wang, T.; Hu, S.; Bhayana, B.; Ishii, M.; Kong, Y.; Cai, Y.; Dai, T.; Cui, W. Bacteria-specific phototoxic reactions triggered by blue light and phytochemical carvacrol. Sci. Transl. Med. 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhu, Y.; Gupta, A.; Huang, Y.; Murray, C.K.; Vrahas, M.S.; Sherwood, M.E.; Baer, D.G.; Hamblin, M.R.; Dai, T. Antimicrobial blue light therapy for multidrug-resistant Acinetobacter baumannii infection in a mouse burn model: Implications for prophylaxis and treatment of combat-related wound infections. J. Infect. Dis. 2014, 209, 1963–1971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamblin, M.R.; Viveiros, J.; Yang, C.; Ahmadi, A.; Ganz, R.A.; Tolkoff, M.J. Helicobacter pylori accumulates photoactive porphyrins and is killed by visible light. Antimicrob. Agents Chemother. 2005, 49, 2822–2827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, M.; Li, Y.; Wu, M.X. Bacteria-specific pro-photosensitizer kills multidrug-resistant Staphylococcus aureus and Pseudomonas aeruginosa. Commun. Biol. 2021, 4, 1–12. [Google Scholar] [CrossRef]
- Halstead, F.D.; Thwaite, J.E.; Burt, R.; Laws, T.R.; Raguse, M.; Moeller, R.; Webber, M.A.; Oppenheim, B.A. Antibacterial activity of blue light against nosocomial wound pathogens growing planktonically and as mature biofilms. Appl. Environ. Microbiol. 2016, 82, 4006–4016. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Dai, T.; Murray, C.K.; Wu, M.X. Bactericidal property of oregano oil against multidrug-resistant clinical isolates. Front. Microbiol. 2018, 9, 2329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courtney, C.M.; Goodman, S.M.; Nagy, T.A.; Levy, M.; Bhusal, P.; Madinger, N.E.; Detweiler, C.S.; Nagpal, P.; Chatterjee, A. Potentiating antibiotics in drug-resistant clinical isolates via stimuli-activated superoxide generation. Sci. Adv. 2017, 3, e1701776. [Google Scholar] [CrossRef] [Green Version]
- Sahl, J.W.; Johnson, J.K.; Harris, A.D.; Phillippy, A.M.; Hsiao, W.W.; Thom, K.A.; Rasko, D.A. Genomic comparison of multi-drug resistant invasive and colonizing Acinetobacter baumannii isolated from diverse human body sites reveals genomic plasticity. BMC Genom. 2011, 12, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Moser, C.; Wang, H.-Z.; Høiby, N.; Song, Z.-J. Strategies for combating bacterial biofilm infections. Int. J. Oral Sci. 2015, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Silva, F.; Lourenço, O.; Queiroz, J.A.; Domingues, F.C. Bacteriostatic versus bactericidal activity of ciprofloxacin in Escherichia coli assessed by flow cytometry using a novel far-red dye. J. Antibiot. 2011, 64, 321–325. [Google Scholar] [CrossRef]
- Zhang, L.; Mah, T.-F. Involvement of a novel efflux system in biofilm-specific resistance to antibiotics. J. Bacteriol. 2008, 190, 4447–4452. [Google Scholar] [CrossRef] [Green Version]
- Dos Anjos, C.; Sabino, C.P.; Sellera, F.P.; Esposito, F.; Pogliani, F.C.; Lincopan, N. Hypervirulent and hypermucoviscous strains of Klebsiella pneumoniae challenged by antimicrobial strategies using visible light. Int. J. Antimicrob. Agents 2020, 56, 106025. [Google Scholar] [CrossRef] [PubMed]
- Hoenes, K.; Bauer, R.; Meurle, T.; Spellerberg, B.; Hessling, M. Inactivation Effect of Violet and Blue Light on ESKAPE Pathogens and Closely Related Non-pathogenic Bacterial Species—A Promising Tool against Antibiotic-Sensitive and Antibiotic-Resistant Microorganisms. Front. Microbiol. 2021, 11, 3429. [Google Scholar] [CrossRef]
- Amin, R.M.; Bhayana, B.; Hamblin, M.R.; Dai, T. Antimicrobial blue light inactivation of Pseudomonas aeruginosa by photo-excitation of endogenous porphyrins: In vitro and in vivo studies. Lasers Surg. Med. 2016, 48, 562–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wu, X.; Chen, J.; Amin, R.; Lu, M.; Bhayana, B.; Zhao, J.; Murray, C.K.; Hamblin, M.R.; Hooper, D.C. Antimicrobial blue light inactivation of gram-negative pathogens in biofilms: In vitro and in vivo studies. J. Infect. Dis. 2016, 213, 1380–1387. [Google Scholar] [CrossRef] [Green Version]
- Dubois-Brissonnet, F.; Trotier, E.; Briandet, R. The biofilm lifestyle involves an increase in bacterial membrane saturated fatty acids. Front. Microbiol. 2016, 7, 1673. [Google Scholar] [CrossRef] [Green Version]
- Hill, K.E.; Malic, S.; McKee, R.; Rennison, T.; Harding, K.G.; Williams, D.W.; Thomas, D.W. An in vitro model of chronic wound biofilms to test wound dressings and assess antimicrobial susceptibilities. J. Antimicrob. Chemother. 2010, 65, 1195–1206. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence 5′ to 3′ | Tm (°C) | Target Gene | Size (bp) | Source |
|---|---|---|---|---|---|
| Sa-F | ATTTGGTCCCAGTGGTGTGGGTAT | 60.4 | hypothetical protein | 143 | Sun et al. [18] |
| Sa-R | GCTGTGACAATTGCCGTTTGTCGT | 60.7 | |||
| Ec-F | CGCGGTGATACATATCCAGCCA | 59.1 | uidA | 179 | This study |
| Ec-R | CGATTTGGAAACGGCAGAGAAGGT | 59.6 | |||
| Kp-F | GGCTTATACCGATAGAGAACTCGAACTG | 58.8 | ureR_1 | 162 | This study |
| Kp-R | TCGATAAAGCCATGAGAATGCAACGG | 59.4 | |||
| Ab-F | GGTTCTTCGCTCCTTTGTCCG | 58.3 | hypothetical protein | 164 | Sahl et al. [31] |
| Ab-R | CTCGGTAATGCTGTTTAACTTCAGGAATATC | 57.3 | |||
| Pa-F | TAAGGACAGCCAGGACTACGAGAA | 59.0 | lasR | 159 | Sun et al. [18] |
| Pa-R | TGGTAGATGGACGGTTCCCAGAAA | 59.8 | |||
| Universal-F | CACAAGCGGTGGAGCATGT | 58.8 | 16S rRNA gene | 145 | This study |
| Universal-R | ACGACACGAGCTGACGACA | 58.8 |
| Planktonic Cells | Tetracycline | Ciprofloxacin | Carvacrol |
|---|---|---|---|
| S. aureus IQ0064 | 0.25; S * | 1; S | 200 |
| K. pneumoniae IQ0035 | 2–4; S | 0.13–0.25; S | 250 |
| A. baumannii 17978 | 1–2; S | 0.13–0.25; S | 100 |
| P. aeruginosa PAO1 | 2–4; S | 0.06; S | 500 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Wu, M.X. Reversal of Polymicrobial Biofilm Tolerance to Ciprofloxacin by Blue Light plus Carvacrol. Microorganisms 2021, 9, 2074. https://doi.org/10.3390/microorganisms9102074
Li Y, Wu MX. Reversal of Polymicrobial Biofilm Tolerance to Ciprofloxacin by Blue Light plus Carvacrol. Microorganisms. 2021; 9(10):2074. https://doi.org/10.3390/microorganisms9102074
Chicago/Turabian StyleLi, Yongli, and Mei X. Wu. 2021. "Reversal of Polymicrobial Biofilm Tolerance to Ciprofloxacin by Blue Light plus Carvacrol" Microorganisms 9, no. 10: 2074. https://doi.org/10.3390/microorganisms9102074
APA StyleLi, Y., & Wu, M. X. (2021). Reversal of Polymicrobial Biofilm Tolerance to Ciprofloxacin by Blue Light plus Carvacrol. Microorganisms, 9(10), 2074. https://doi.org/10.3390/microorganisms9102074