1. Introduction
Seasonal influenza vaccines contain influenza A and B viruses, providing optimal protection against circulating influenza strains. The need for influenza vaccines with broad protection is urgent to reduce the influenza burden worldwide. Live attenuated influenza vaccines (LAIVs) provide broader cross-protection compared to inactivated vaccines due to the considerable involvement of T-cell immunity, the generation of resident memory T cells targeted at conserved viral epitopes in respiratory airways, and the induction of mucosal secretory antibody responses [
1,
2,
3,
4].
Influenza B viruses are less variable than influenza A viruses [
5,
6]. Two lineages of influenza B are specified on the basis of hemagglutinin (HA) antigenic properties: sera against one lineage do not cross-react with viruses of another lineage [
6], although some level of cross-protection has been demonstrated [
7,
8]. The highest burden of influenza B virus infection is registered in children, but these viruses can also cause complicated cases in the elderly population [
9,
10]. Natural reassortment between the influenza A and B viruses does not occur, which is restricted by the specificity of viral RNA packaging signals [
11,
12]. For effective replication of a cross-type reassortant, specific features of the polymerase have to match the features of the virus subtype, first of all, the structure of promoter elements in the noncoding regions of RNA segments. The impact of other viral regions on polymerase machinery activity has also been shown [
13]. Nevertheless, the capacity of the influenza genome to express foreign proteins has been extensively studied in multiple experiments with reporter genes and chimeric constructs [
14,
15,
16,
17,
18,
19]. These studies have shown which gene fragments are necessary for packaging genome segments into virions [
14,
16]. Engineered reassortants between the influenza A and B viruses have been previously obtained by several study groups using a H1N1 influenza virus backbone [
11,
12,
20,
21]. Horimoto et al. [
20] studied different variants of A/B chimeric HAs on the base of the A/WSN/33 strain with the insertion of B/Lee/40 fragments. In Flandorfer et al. [
12], the reassortants with both HA and neuraminidase (NA) were rescued on the base of the A/WSN/33 and B/Yamagata/16/88 strains. The same influenza B strain was used by Baker et al. [
11]; as a backbone, A/PR/8/34 virus was used. Some of the chimeric constructions were tested for biological characteristics and immunogenicity in animal models [
21,
22]. In a study by Horimoto et al. [
22], immunogenicity of the chimeric A/B virus was investigated by an hemagglutination inhibition (HAI) assay with influenza B antigen. For live viral vaccines, it was established that one of the major mechanisms providing protection against respiratory viruses is the formation of resident memory T-cell responses in the respiratory airways, targeted at epitopes of different pathogen’s proteins [
23]. These epitopes may arise from the fragments of conservative protein domains, providing basic protection against different virus subtypes in the future. Therefore, experimental assessment of immunogenicity and protective potential of a chimeric live vaccine based on influenza A and B reassortants can be informative for the development of universal cross-type influenza vaccines.
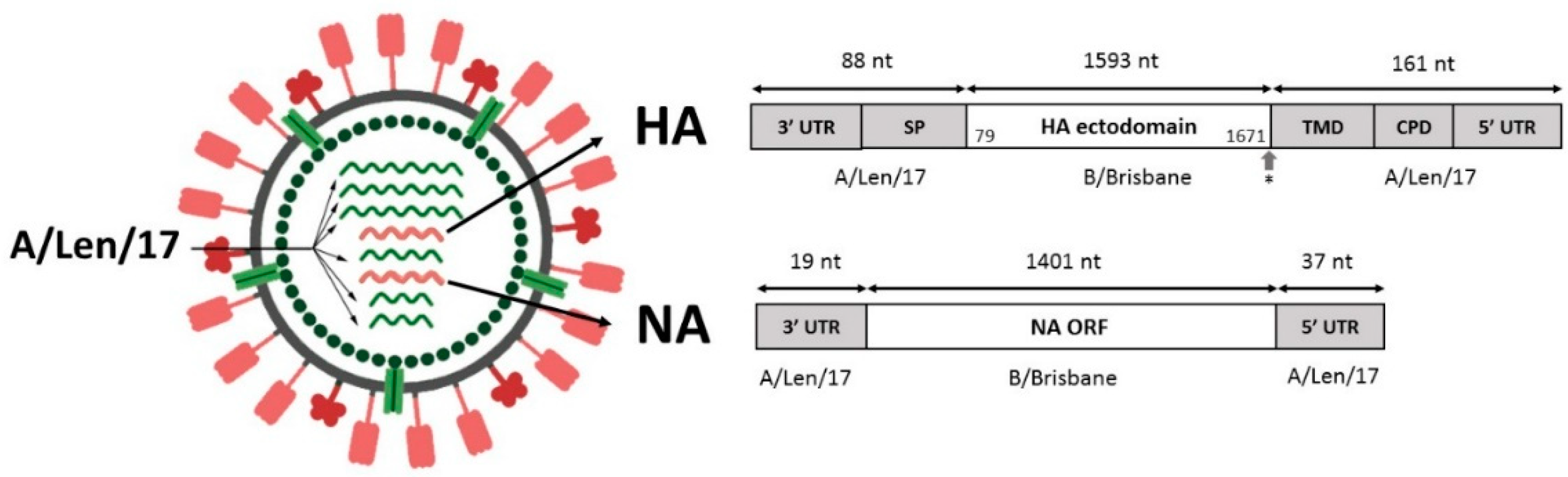
The A/Leningrad/134/17/57 (H2N2) strain is licensed for influenza A LAIV strain development [
24]. Since this LAIV backbone virus has a strong safety record and has been on the market for decades, its use for designing a new generation of influenza vaccines is a very promising strategy. All previously studied chimeric constructions were obtained on the base of H1N1 strains. However, those viruses cannot be widely used for the development of human live attenuated vaccines due to the lack of safety data. We aimed to study if the approach of engineering intertype reassortants can be applied to the cold-adapted attenuated influenza A virus of the H2N2 subtype.
In this study, we rescued an engineered influenza A/B reassortant on the backbone of a licensed Russian LAIV master donor virus—the strain A/Leningrad/134/17/57 (H2N2)—and assessed its biological characteristics, stability, immunogenicity, and protective efficacy in a mouse model.
4. Discussion
The influenza virus’s high variability requires an annual revision of influenza vaccines composition. In a common trivalent formulation, one influenza B strain is included while two strains of different genetic lineages of influenza B are compounds of a quadrivalent formulation. Despite the burden of influenza B in high-risk groups [
9,
10], in seasons with antigenic mismatch, the rate of influenza B in overall morbidity can reach 70% [
9,
56], so the role of the influenza B vaccine should not be underestimated. During mixed infection, influenza viruses can interfere, and the impact of this process on live vaccine effectiveness is not fully understood. Influenza B has lower variability and evolution rate [
5,
57,
58]. On the other hand, the suppression of influenza A replication by influenza B NP has been found [
59]. The lower variability of influenza B could provide a cross-reactivity of immunity factors and consequently a faster immunity activation after boost with a new vaccine dose; this is discussed as a possible reason for the low vaccine effectiveness of the influenza A component (reviewed in [
60]). The ability to prepare strains of both subtypes on the one backbone, in theory, could be a key to solving the interference problem.
Experiments with intertype reassortant rescue have been successfully performed by several groups of scientists [
11,
12,
20,
21]. As a backbone, two H1N1 viruses were used: A/WSN/33 [
12,
20,
21] and A/Puerto Rico/8/34 [
11]. As a source of influenza B genes, the B/Lee/40 strain was used [
20], or B/Yamagata/16/88 [
11,
12]. However, the aforementioned H1N1 viruses cannot be widely used for the development of human live attenuated vaccines due to the lack of safety data. In our study, the intertype reassortant was rescued on the H2N2 virus backbone with the influenza B strain of B/Victoria lineage, demonstrating that the strategy of the intertype reassortants’ design is universal for influenza viruses of different subtypes. The backbone strain used in this study, A/Len/17, is a master donor virus for a licensed live attenuated influenza vaccine, which has been extensively studied in multiple clinical and epidemiological trials (reviewed in [
61]), with established safety, genetic stability, immunogenicity, and effectiveness. In the current study, we performed proof-of-concept experiments with a rescue of an A/Len/17-based A/B reassortant and tested its replication capacity, phenotype, immunogenicity, and ability to protect animals in challenge experiments.
In other studies, A/B chimeric viruses have been shown to have reduced titers and attenuated phenotype in animals [
12,
20,
21]. The H2B chimeric strain on A/Len/17 backbone inherited
ts,
ca, and attenuated phenotypes from the vaccine backbone.
The in vitro characteristics of the H2B virus stock were standard for LAIV strains [
24,
30]. Directly after rescue, the virus had a low titer and low HA activity, but serial passaging in eggs improved its growth characteristics. Sequencing revealed heterogeneity in the virus stock population with an appearance of variants with the substitution G141E in HA specific for influenza B strain egg adaptation [
41,
62]. The observed changes in PB1 and NS1 could also impact growth characteristics, but mostly important, these substitutions did not alter the
ts phenotype of the virus and its attenuation in mouse experiment. Nevertheless, the H2B chimeric virus had lower titers in MDCK cells than both parental viruses (A/Len/17 and B/Brisbane/60/08). Of note, the control A/LAIV was also unable to efficient replication in MDCK cells, but still, this vaccine was effective in clinical trials [
63], indicating that the virus titer in MDCK is not the main correlate of the vaccine quality. A more significant observation is that the H2B LAIV tends to replicate to lower titers in the upper respiratory tract of mice compared to control vaccines, which could be the reason for lower immunogenicity and decreased protection against influenza A virus. Although these differences were not significant, the tendency for the chimeric vaccine to be less immunogenic than the control B/LAIV could be related to the synergistic effect of the attenuating properties of the backbone and consequences of the viral genetic modification.
In this study, a prime-boost immunization schedule was used to induce optimal immune responses. Although we did not assess antibody and T cell-based immunity after prime immunization, previous studies suggest that a booster LAIV dose indeed significantly enhances adaptive immunity in mice [
64,
65,
66]. The antigenic properties of the constructed chimeric virus assessed in HAI with different sera confirmed that heterologous TMD and CPD in influenza HA did not affect the antigenic properties of the molecule. The sera of H2B-immunized animals reacted with the influenza B antigen in the HAI test and neutralized infectious influenza B virus replication. These data are in line with experiments by Horimoto et al. [
22]: after mice infection with an A/WSN/33-based chimeric virus with flu B HA, antibodies to the B antigen in the HAI test were detected. In the current study, HAI and MNT titers after chimeric virus immunization were relatively low, but they were sufficient to protect animals against influenza B virus challenge. It is possible that T-cell responses to HA and NA could have contributed to protection, since serum antibodies are not the only correlate of protection after LAIV immunization [
67].
The HA transmembrane domain plays a significant role during fusion, thus affecting the overall stability of the HA molecule [
35,
68,
69]. Low immunogenicity of viruses with unstable HA has been previously reported [
36]. During the development of chimeric construction design, we took into consideration the fact that chimeric HAs have to inherit fragments of coding regions from the backbone virus for successful rescue [
12,
18,
19,
20]. On the other hand, the stalk region of HA is a significant determinant for cross-reactive antibody response induction [
70,
71]. Thus, we tested the strategy with TMD and CPD inherited from a backbone virus and other HA domains from a wild-type virus. The possible consequence of inconsistency of the HA transmembrane and stalk domains inherited from viruses of different subtypes could be an unstable HA structure, so we tested this hypothesis in an HA thermostability test. The inhibition of hemagglutination after incubation at high temperatures is an indirect method of assessing HA stability [
36,
72]. In this test, the chimeric HA has been shown to be stable despite the inconsistency of the ectodomain and transmembrane part of the molecule.
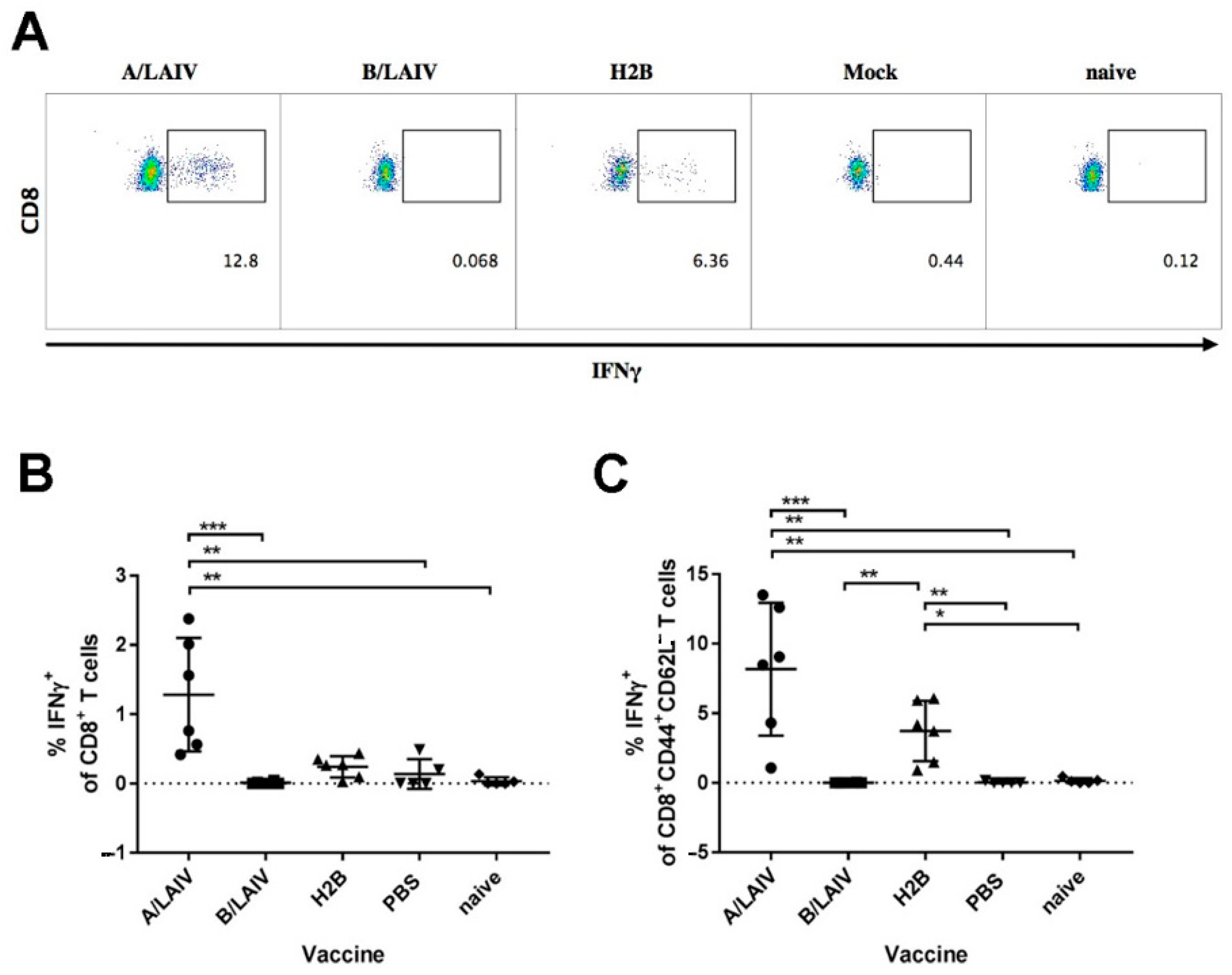
The development of reassortant with influenza B HA and NA on the influenza A backbone gave us a unique opportunity to assess the immunity against influenza A without the impact of influenza A HA and NA proteins in experiments with a live replicating virus. As expected, influenza A neutralizing antibodies and hemagglutination inhibition by the sera of chimera-immunized animals were not detected. IgG antibodies against influenza A were detected by ELISA, and these could be antibody targeted at NP, M1, or M2 proteins. Though antibodies in the internal proteins have no direct virus neutralizing activity, their role in protection, such as NK-cell activation, has been discussed [
73]. Importantly, the chimeric vaccine induced significant levels of T
EM cells (CD44
+CD62L
− phenotype) in spleens specific to A/Len/17 NP
366-374 peptide. Although we did not assess T-cell immunity in the whole influenza A virus, stimulation of the splenocytes with the most immunodominant epitope reflects the overall cell-mediated immune responses to vaccination, and stimulation with the whole virus yields similar results in terms of intergroup comparison [
52].
In Horimoto et al.’s experiments [
47] with A/B chimeras, the protective response to the backbone influenza A virus was shown. In these chimeras, only HA was transferred to the chimeric virus. NA was from the backbone virus. In addition, the backbone that was used to prepare the chimeric viruses was not attenuated, and the same virus was used for challenge. In our study, we used more relevant influenza A backbone for the construction of a chimeric vaccine, a LAIV master donor virus of H2N2 subtype, which is routinely used for preparation of seasonal influenza vaccines against circulating H1N1 and H3N2 influenza strains. Here, we were unable to challenge mice with the original H2N2 1957 pandemic virus due to safety reasons. However, influenza viruses share common T-cell epitopes that, in theory, could provide partial protection against infection with drifted influenza viruses [
74]. Therefore, we selected a recent heterologous influenza A/H1N1 to assess protective effect of the chimeric vaccine against circulating strains. Despite effective T-cell immunity stimulation by the H2B chimeric vaccine, it was not sufficient to protect animals against a high-dose influenza A virus of the heterologous subtype. It is of special note that the most immunodominant epitopes of influenza A virus internal proteins have significantly evolved since 1957 (the year of A/Len/17 strain isolation) and are not present in the currently circulating viruses anymore. Furthermore, the differences even in one immunodominant epitope, such as NP
366, can significantly alter the protective efficacy of the LAIVs in C57BL/6J mice [
46]. Indeed, matching of the internal proteins’ sequences revealed that a high proportion of H-2
b-restricted T-cell epitopes of A/Len/17 virus differ from that of A/California/07/09 virus, so the protective role of T-cell immunity formed after immunization in C57BL/6J mice could be decreased.
We discussed earlier the issue of mismatch between the T-cell epitope composition in reassortant viruses for live and inactivated influenza vaccines and currently the circulating influenza viruses and made recommendations reconsidering the genome composition of the vaccines by the replacement of either NP or M genes of the old attenuated backbone virus with that of a recent influenza strain, which would enhance the cross-protective properties of the vaccines in terms of optimizing T-cell immunity [
75]. Our current study further emphasized the necessity for optimizing genome composition of the chimeric vaccine construct to enhance the cross-reactivity of T-cell responses against influenza A viruses. Further experiments with optimized construction designs, vaccine dosages, and immunization schedules are warranted, as they could improve immunogenicity and the protective potential of the chimeric vaccine.


 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}