
Effects of an EPS Biosynthesis Gene Cluster of Paenibacillus polymyxa WLY78 on Biofilm Formation and Nitrogen Fixation under Aerobic Conditions

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Plasmids, Primers, and Media
2.2. Construction of ∆pep-1 and ∆pep-2 Mutants
2.3. Biofilm Assay
2.4. Isolation, Extraction and Analysis of EPS
2.5. Colonial Morphology and Motility Assays
2.6. Acetylene Reduction Assay for Nitrogenase Activity
2.7. Statistical Analysis
3. Results
3.1. The Effects of Carbon Sources and Nitrogen Sources on Biofilm in P. polymyxa WLY78
3.2. Identification of Polysaccharide Biosynthesis Gene Clusters in P. polymyxa WLY78
3.3. The pep-2 Gene Cluster Is Involved in Biofilm Formation and Polysaccharide Biosynthesis
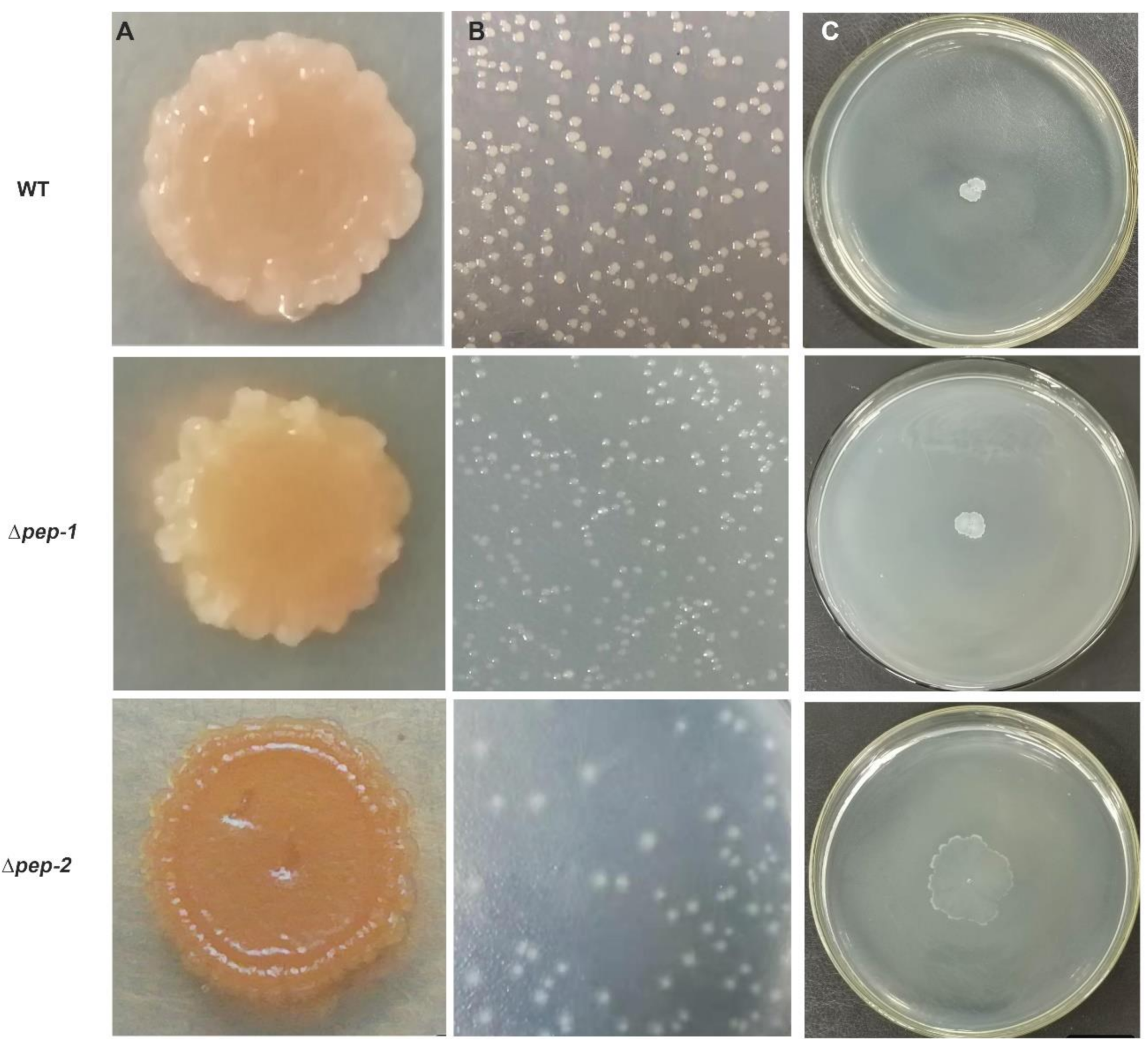
3.4. EPS Biosynthesis Defect Is Effect on Colony Biofilm, Colony Morphology and Motility
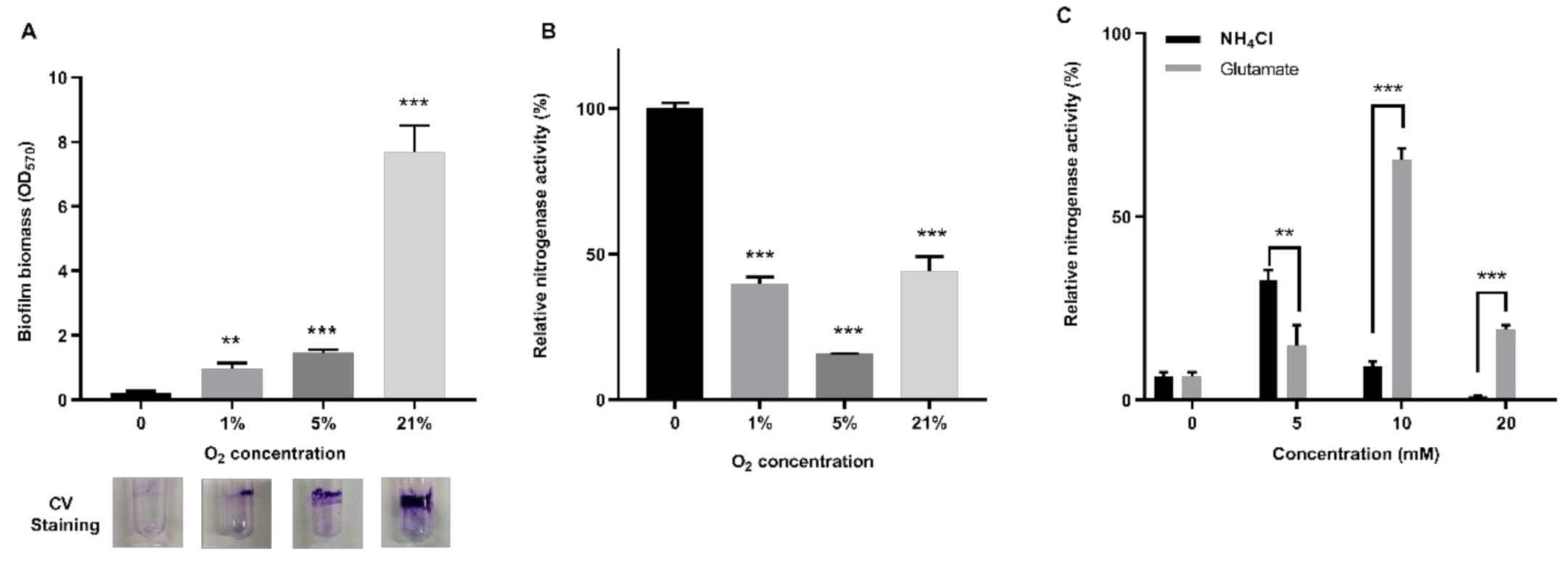
3.5. Biofilm Formation Mediated by EPS Enables Bacteria to Fix Nitrogen in Air
3.6. Impacts of Nitrogen and Oxygen Content on Nitrogenase Activity in Biofilm
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [Green Version]
- Hall-Stoodley, L.; Stoodley, P. Biofilm formation and dispersal and the transmission of human pathogens. Trends Microbiol. 2005, 13, 7–10. [Google Scholar] [CrossRef]
- Voloshin, S.; Shleeva, M.; Syroeshkin, A.; Kaprelyants, A. The role of intercellular contacts in the initiation of growth and in the development of a transiently nonculturable state by cultures of Rhodococcus rhodochrous grown in poor media. Microbiology 2005, 74, 420–427. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, Z.; Huang, Z.; Zhang, X.; Li, D.; Sun, L.; You, J.; Pan, X.; Yang, H. A putative LysR-type transcriptional regulator inhibits biofilm synthesis in Pseudomonas aeruginosa. Biofouling 2019, 35, 541–550. [Google Scholar] [CrossRef]
- Liu, J.; Guo, L.; Liu, J.; Zhang, J.; Zeng, H.; Ning, Y.; Wei, X. Identification of an efflux transporter lmrb regulating stress response and extracellular polysaccharide synthesis in Streptococcus mutans. Front. Microbiol. 2017, 8, 962. [Google Scholar] [CrossRef] [Green Version]
- Mahdhi, A.; Leban, N.; Chakroun, I.; Bayar, S.; Mahdouani, K.; Majdoub, H.; Kouidhi, B. Use of extracellular polysaccharides, secreted by Lactobacillus plantarum and Bacillus spp. as reducing indole production agents to control biofilm formation and efflux pumps inhibitor in Escherichia coli. Microb. Pathog. 2018, 125, 448–453. [Google Scholar] [CrossRef]
- Kearns, D.B.; Chu, F.; Branda, S.S.; Kolter, R.; Losick, R. A master regulator for biofilm formation by Bacillus subtilis. Mol. Microbiol. 2005, 55, 739–749. [Google Scholar] [CrossRef]
- Colledge, V.L.; Fogg, M.J.; Levdikov, V.M.; Leech, A.; Dodson, E.J.; Wilkinson, A.J. Structure and organisation of SinR, the master regulator of biofilm formation in Bacillus subtilis. J. Mol. Biol. 2011, 411, 597–613. [Google Scholar] [CrossRef] [Green Version]
- Newman, J.A.; Rodrigues, C.; Lewis, R.J. Molecular basis of the activity of SinR protein, the master regulator of biofilm formation in Bacillus subtilis. J. Biol. Chem. 2013, 288, 10766–10778. [Google Scholar] [CrossRef] [Green Version]
- Vlamakis, H.; Chai, Y.; Beauregard, P.; Losick, R.; Kolter, R. Sticking together: Building a biofilm the Bacillus subtilis way. Nat. Rev. Microbiol. 2013, 11, 157. [Google Scholar] [CrossRef] [Green Version]
- Mielichsuss, B.; Lopez, D. Molecular mechanisms involved in Bacillus subtilis biofilm formation. Environ. Microbiol. 2015, 17, 555–565. [Google Scholar] [CrossRef] [Green Version]
- Vu, B.; Chen, M.; Crawford, R.J.; Ivanova, E.P. Bacterial extracellular polysaccharides involved in biofilm formation. Molecules 2009, 14, 2535–2554. [Google Scholar] [CrossRef]
- Chai, Y.; Chu, F.; Kolter, R.; Losick, R. Bistability and biofilm formation in Bacillus subtilis. Mol. Microbiol. 2008, 67, 254–263. [Google Scholar] [CrossRef] [Green Version]
- Chai, Y.; Norman, T.; Kolter, R.; Losick, R. An epigenetic switch governing daughter cell separation in Bacillus subtilis. Genes Dev. 2010, 24, 754–765. [Google Scholar] [CrossRef] [Green Version]
- Chu, F.; Kearns, D.B.; Branda, S.S.; Kolter, R.; Losick, R. Targets of the master regulator of biofilm formation in Bacillus subtilis. Mol. Microbiol. 2006, 59, 1216–1228. [Google Scholar] [CrossRef]
- Zivkovic, M.; Miljkovic, M.; Ruas-Madiedo, P.; Strahinic, I.; Tolinacki, M.; Golic, N.; Kojic, M. Exopolysaccharide production and ropy phenotype are determined by two gene clusters in putative probiotic strain Lactobacillus paraplantarum BGCG11. Appl. Environ. Microbiol. 2015, 81, 1387–1396. [Google Scholar] [CrossRef] [Green Version]
- Schäper, S.; Wendt, H.; Bamberger, J.; Sieber, V.; Schmid, J.; Becker, A. A bifunctional UDP-sugar 4-epimerase supports biosynthesis of multiple cell surface polysaccharides in Sinorhizobium meliloti. J. Bacteriol. 2019, 201, e00801–e00818. [Google Scholar] [CrossRef] [Green Version]
- Downie, J.A. The roles of extracellular proteins, polysaccharides and signals in the interactions of rhizobia with legume roots. FEMS Microbiol. Rev. 2010, 34, 150–170. [Google Scholar] [CrossRef]
- Lerner, A.; Castro-Sowinski, S.; Valverde, A.; Lerner, H.; Dror, R.; Okon, Y.; Burdman, S. The Azospirillum brasilense Sp7 noeJ and noeL genes are involved in extracellular polysaccharide biosynthesis. Microbiology 2009, 155, 4058–4068. [Google Scholar] [CrossRef] [Green Version]
- Timmusk, S.; Grantcharova, N.; Wagner, E.G.H. Paenibacillus polymyxa invades plant roots and forms biofilms. Appl. Environ. Microbiol. 2005, 71, 7292–7300. [Google Scholar] [CrossRef] [Green Version]
- Timmusk, S.; Copolovici, D.; Copolovici, L.; Teder, T.; Nevo, E.; Behers, L. Paenibacillus polymyxa biofilm polysaccharides antagonise Fusarium graminearum. Sci. Rep. 2019, 9, 662. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Mishra, A.; Chauhan, P.; Nautiyal, C. Induction of Paenibacillus lentimorbus biofilm by sodium alginate and CaCl2 alleviates drought stress in chickpea. Ann. Appl. Biol. 2011, 159, 372–386. [Google Scholar] [CrossRef]
- Yegorenkova, I.V.; Tregubova, K.V.; Matora, L.Y.; Burygin, G.L.; Ignatov, V.V. Biofilm formation by Paenibacillus polymyxa strains differing in the production and rheological properties of their exopolysaccharides. Curr. Microbiol. 2011, 62, 1554–1559. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.-H.; Jung, H.K. Antioxidant and antitumor activities of β-glucan-rich exopolysaccharides with different molecular weight from Paenibacillus polymyxa JB115. J. Korean Soc. Appl. Biol. Chem. 2014, 57, 105–112. [Google Scholar] [CrossRef]
- Liu, J.; Luo, J.; Ye, H.; Sun, Y.; Lu, Z.; Zeng, X. Production, characterization and antioxidant activities in vitro of exopolysaccharides from endophytic bacterium Paenibacillus polymyxa EJS-3. Carbohydr. Polym. 2009, 78, 275–281. [Google Scholar] [CrossRef]
- Mokaddem, H.; Sadaoui, Z.; Boukhelata, N.; Azouaou, N.; Kaci, Y. Removal of Cadmium from aqueous solution by polysaccharide produced from Paenibacillus polymyxa. J. Hazard. Mater. 2009, 172, 1150–1155. [Google Scholar] [CrossRef]
- Tang, J.; Qi, S.; Li, Z.; An, Q.; Xie, M.; Yang, B.; Wang, Y. Production, purification and application of polysaccharide-based bioflocculant by Paenibacillus mucilaginosus. Carbohydr. Polym. 2014, 113, 463–470. [Google Scholar] [CrossRef]
- Debois, D.; Ongena, M.; Cawoy, H.; De Pauw, E. MALDI-FTICR MS imaging as a powerful tool to identify Paenibacillus antibiotics involved in the Inhibition of plant pathogens. J. Am. Soc. Mass Spectrom. 2013, 24, 1202–1213. [Google Scholar] [CrossRef]
- Rubio, L.M.; Ludden, P.W. Biosynthesis of the iron-molybdenum cofactor of nitrogenase. Annu. Rev. Microbiol. 2008, 62, 93–111. [Google Scholar] [CrossRef] [Green Version]
- Dixon, R.; Kahn, D. Genetic regulation of biological nitrogen fixation. Nat. Rev. Microbiol. 2004, 2, 621–631. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, L.; Liu, Z.; Zhao, D.; Liu, X.; Zhang, B.; Xie, J.; Hong, Y.; Li, P.; Chen, S. A minimal nitrogen fixation gene cluster from Paenibacillus sp. WLY78 enables expression of active nitrogenase in Escherichia coli. PLoS Genet. 2013, 9, e1003865. [Google Scholar] [CrossRef]
- Xie, J.; Shi, H.; Du, Z.; Wang, T.; Liu, X.; Chen, S. Comparative genomic and functional analysis reveal conservation of plant growth promoting traits in Paenibacillus polymyxa and its closely related species. Sci. Rep. 2016, 6, 21329. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.-B.; Du, Z.; Bai, L.; Tian, C.; Zhang, Y.; Xie, J.-Y.; Wang, T.; Liu, X.; Chen, X.; Cheng, Q. Comparative genomic analysis of N2-fixing and non-N2-fixing Paenibacillus spp.: Organization, evolution and expression of the nitrogen fixation genes. PLoS Genet. 2014, 10, e1004231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villafane, R.; Bechhofer, D.; Narayanan, C.; Dubnau, D. Replication control genes of plasmid pE194. J. Bacteriol. 1987, 169, 4822–4829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.; Kumar, M.; Pandey, A.; Thakur, I.S. Genomic analysis of carbon dioxide sequestering bacterium for exopolysaccharides production. Sci. Rep. 2019, 9, 4270. [Google Scholar] [CrossRef]
- Adebayo-tayo, B.C.; Onilude, A.A. Screening of lactic acid bacteria strains isolated from some Nigerian fermented foods for EPS production. World Appl. Sci. J. 2008, 4, 741–747. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Rice, K.C.; Mann, E.E.; Endres, J.L.; Weiss, E.C.; Cassat, J.E.; Smeltzer, M.S.; Bayles, K.W. The cidA murein hydrolase regulator contributes to DNA release and biofilm development in Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2007, 104, 8113–8118. [Google Scholar] [CrossRef] [Green Version]
- Jackson, K.D.; Starkey, M.; Kremer, S.; Parsek, M.R.; Wozniak, D.J. Identification of psl, a locus encoding a potential exopolysaccharide that is essential for Pseudomonas aeruginosa PAO1 biofilm formation. J. Bacteriol. 2004, 186, 4466–4475. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Xu, A.; Elmerich, C.; Ma, L.Z. Biofilm formation enables free-living nitrogen-fixing rhizobacteria to fix nitrogen under aerobic conditions. ISME J. 2017, 11, 1602. [Google Scholar] [CrossRef] [Green Version]
- Rütering, M.; Cress, B.F.; Schilling, M.; Rühmann, B.; Koffas, M.A.; Sieber, V.; Schmid, J. Tailor-made exopolysaccharides—CRISPR-Cas9 mediated genome editing in Paenibacillus polymyxa. Synth. Biol. 2017, 2, ysx007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Cole, R.A.; Reeves, P.R. An O-antigen processing function for Wzx (RfbX): A promising candidate for O-unit flippase. J. Bacteriol. 1996, 178, 2102–2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodward, R.; Yi, W.; Li, L.; Zhao, G.; Eguchi, H.; Sridhar, P.R.; Guo, H.; Song, J.K.; Motari, E.; Cai, L. In vitro bacterial polysaccharide biosynthesis: Defining the functions of Wzy and Wzz. Nat. Chem. Biol. 2010, 6, 418. [Google Scholar] [CrossRef] [PubMed]
- Elsholz, A.K.; Wacker, S.A.; Losick, R. Self-regulation of exopolysaccharide production in Bacillus subtilis by a tyrosine kinase. Genes Dev. 2014, 28, 1710–1720. [Google Scholar] [CrossRef] [Green Version]
- Leiman, S.A.; Arboleda, L.C.; Spina, J.S.; Mcloon, A.L. SinR is a mutational target for fine-tuning biofilm formation in laboratory-evolved strains of Bacillus subtilis. BMC Microbiol. 2014, 14, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Bai, L.; Wang, L.; Chen, S. Phylogeny of 16S rRNA and nifH genes and regulation of nitrogenase activity by oxygen and ammonium in the genus Paenibacillus. Microbiology 2012, 81, 702–709. [Google Scholar] [CrossRef]
- Péant, B.; LaPointe, G.; Gilbert, C.; Atlan, D.; Ward, P.; Roy, D. Comparative analysis of the exopolysaccharide biosynthesis gene clusters from four strains of Lactobacillus rhamnosus. Microbiology 2005, 151, 1839–1851. [Google Scholar] [CrossRef] [Green Version]
- Mijakovic, I.; Deutscher, J. Protein-tyrosine phosphorylation in Bacillus subtilis: A 10-year retrospective. Front. Microbiol. 2015, 6, 18. [Google Scholar] [CrossRef]
- Gerwig, J.; Kiley, T.B.; Gunka, K.; Stanley-Wall, N.; Stülke, J. The protein tyrosine kinases EpsB and PtkA differentially affect biofilm formation in Bacillus subtilis. Microbiology 2014, 160 Pt 4, 682. [Google Scholar] [CrossRef] [Green Version]
- Guttenplan, S.B.; Blair, K.M.; Kearns, D.B. The EpsE flagellar clutch is bifunctional and synergizes with EPS biosynthesis to promote Bacillus subtilis biofilm formation. PLoS Genet. 2010, 6, e1001243. [Google Scholar] [CrossRef]
- Branda, S.S.; Chu, F.; Kearns, D.B.; Losick, R.; Kolter, R. A major protein component of the Bacillus subtilis biofilm matrix. Mol. Microbiol. 2006, 59, 1229–1238. [Google Scholar] [CrossRef]
- Molle, V.; Fujita, M.; Jensen, S.T.; Eichenberger, P.; González-Pastor, J.E.; Liu, J.S.; Losick, R. The Spo0A regulon of Bacillus subtilis. Mol. Microbiol. 2003, 50, 1683–1701. [Google Scholar] [CrossRef] [Green Version]
- Lemon, K.P.; Higgins, D.E.; Kolter, R. Flagellar motility is critical for Listeria monocytogenes biofilm formation. J. Bacteriol. 2007, 189, 4418–4424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barken, K.B.; Pamp, S.J.; Yang, L.; Gjermansen, M.; Bertrand, J.J.; Klausen, M.; Givskov, M.; Whitchurch, C.B.; Engel, J.N.; Tolker-Nielsen, T. Roles of type IV pili, flagellum-mediated motility and extracellular DNA in the formation of mature multicellular structures in Pseudomonas aeruginosa biofilms. Environ. Microbiol. 2008, 10, 2331–2343. [Google Scholar] [CrossRef] [PubMed]
- Wood, T.K.; Barrios, A.F.G.; Herzberg, M.; Lee, J. Motility influences biofilm architecture in Escherichia coli. Appl. Microbiol. Biotechnol. 2006, 72, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Parsek, M.R.; Tolker-Nielsen, T. Pattern formation in Pseudomonas aeruginosa biofilms. Curr. Opin. Microbiol. 2008, 11, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Merritt, P.M.; Danhorn, T.; Fuqua, C. Motility and chemotaxis in Agrobacterium tumefaciens surface attachment and biofilm formation. J. Bacteriol. 2007, 189, 8005–8014. [Google Scholar] [CrossRef] [Green Version]
- Sabra, W.; Zeng, A.-P.; Lünsdorf, H.; Deckwer, W.-D. Effect of oxygen on formation and structure of Azotobacter vinelandii alginate and its role in protecting nitrogenase. Appl. Environ. Microbiol. 2000, 66, 4037–4044. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predicted Gene Products | Length [aa] | Predicted Function | Predicted Protein Family | Identity with Those of P. polymyxa DSM 365 |
|---|---|---|---|---|
| pep-2 gene cluster | ||||
| PepA (GM004746) | 247 | Capsular biosynthesis protein | YveK | 100% |
| PepB (GM004745) | 212 | tyrosine protein kinase | P-loop_NTPase | 100% |
| GalU (GM004744) | 300 | UTP-glucose-1-phosphate uridilytransferase | Glyco_tranf_GTA_type (Glycosyltransferase family A) | 100% |
| PepC (GM004743) | 232 | undecaprenyl-phosphate galactose phosphotransferase | Bac_transf (Bacterial sugar transferase) | 99.57% |
| PepD (GM004742) | 311 | undecaprenyl-phosphate galactose phosphotransferase | Glyco_tranf_GTA_type | 99.36% |
| PepE (GM004741) | 445 | polysaccharide polymerase | Wzy_C | 99.78% |
| PepF (GM004740) | 251 | UDP-N-acetyl-D-mannosaminuronic acid transferase | Glycosyltransferase WecG/TagA | 100% |
| PepG (GM004739) | 480 | polysaccharide polymerase | Wzy_C | 99.38% |
| PepH (GM004738) | 472 | flippase | MATE (Multidrug and toxic compound extrusion) like | 99.79% |
| PepI (GM004737) | 276 | glycosyltransferase GT2 | Beta4 Glucosyltransferase | 99.64% |
| PepJ (GM004736) | 395 | glycosyltransferase GT4 | Glycosyltransferase_GTB-type | 100% |
| PepK (GM004735) | 382 | glycosyltransferase GT1 | Glycosyltransferase_GTB-type | 100% |
| PepL (GM004734) | 354 | glycosyltransferase GT4 | Glycosyltransferase_GTB-type | 100% |
| UgdH1 (GM004733) | 445 | UDP-glucose 6-dehydrogenase | NADB[NAD(P)H/NAD(P)(+) binding]_Rossmann | 100% |
| ManC (GM004732) | 458 | mannose-1-phosphate guanylyltransferase | Glyco_tranf_GTA_type | 99.56% |
| PepM (GM004731) | 535 | glycoside hydrolase | Glycosyl hydrolase family 26 | 96.45% |
| PepN (GM004730) | 741 | glycoside hydrolase | CRISPR/Cas system-associated protein Cas4 | 97.98% |
| pep-1 gene cluster | ||||
| PepO (GM004727) | 249 | polysaccharide chain length regulation protein | YveK | 97.59% |
| PepP (GM004726) | 228 | tyrosine protein kinase | P-loop_NTPase | 98.68% |
| PepQ (GM004725) | 192 | undecaprenyl-phosphate glucose phosphotransferase | Bac_transf | 98.96% |
| Fcl (GM004724) | 312 | GDP-L-fucose synthase | NADB_Rossmann | 98.40% |
| Gmd (GM004723) | 329 | GDP-mannose 4,6-dehydratase | NADB_Rossmann | 99.39% |
| PepR (GM004722) | 447 | putative flippase | MATE_like | 99.78% |
| PepS (GM004721) | 420 | putative flippase | No domains | 100% |
| PepT (GM004719) | 416 | glycosyltransferase GT1 | Glycosyltransferase_GTB-type | 96.88% |
| PepU (GM004718) | 405 | glycosyltransferase GT1 | Glycosyltransferase_GTB-type | 97.78% |
| PepV (GM004717) | 269 | UDP-N-acetyl-D-mannosaminuronic acid transferase | Glycosyltransferase WecG/TagA | 95.54% |
| UgdH2 (GM004716) | 456 | UDP-glucose 6-dehydrogenase | NADB_Rossmann | 99.12% |
| LytR (GM004715) | 325 | transcriptional regulator | LytR_cpsA_psr | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, X.; Li, Q.; Wang, N.; Chen, S. Effects of an EPS Biosynthesis Gene Cluster of Paenibacillus polymyxa WLY78 on Biofilm Formation and Nitrogen Fixation under Aerobic Conditions. Microorganisms 2021, 9, 289. https://doi.org/10.3390/microorganisms9020289
He X, Li Q, Wang N, Chen S. Effects of an EPS Biosynthesis Gene Cluster of Paenibacillus polymyxa WLY78 on Biofilm Formation and Nitrogen Fixation under Aerobic Conditions. Microorganisms. 2021; 9(2):289. https://doi.org/10.3390/microorganisms9020289
Chicago/Turabian StyleHe, Xiaojuan, Qin Li, Nan Wang, and Sanfeng Chen. 2021. "Effects of an EPS Biosynthesis Gene Cluster of Paenibacillus polymyxa WLY78 on Biofilm Formation and Nitrogen Fixation under Aerobic Conditions" Microorganisms 9, no. 2: 289. https://doi.org/10.3390/microorganisms9020289