
Involvement of a Quorum Sensing Signal Molecule in the Extracellular Amylase Activity of the Thermophilic Anoxybacillus amylolyticus


 ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. A. amylolyticus Cultivation
2.3. Extraction of Spent Medium of A. amylolyticus
2.4. Identification of Autoinducer
2.4.1. TLC-Overlay Assay
2.4.2. LC-MS/MS Analysis
2.5. A. amylolyticus α-amylase Assay
3. Results
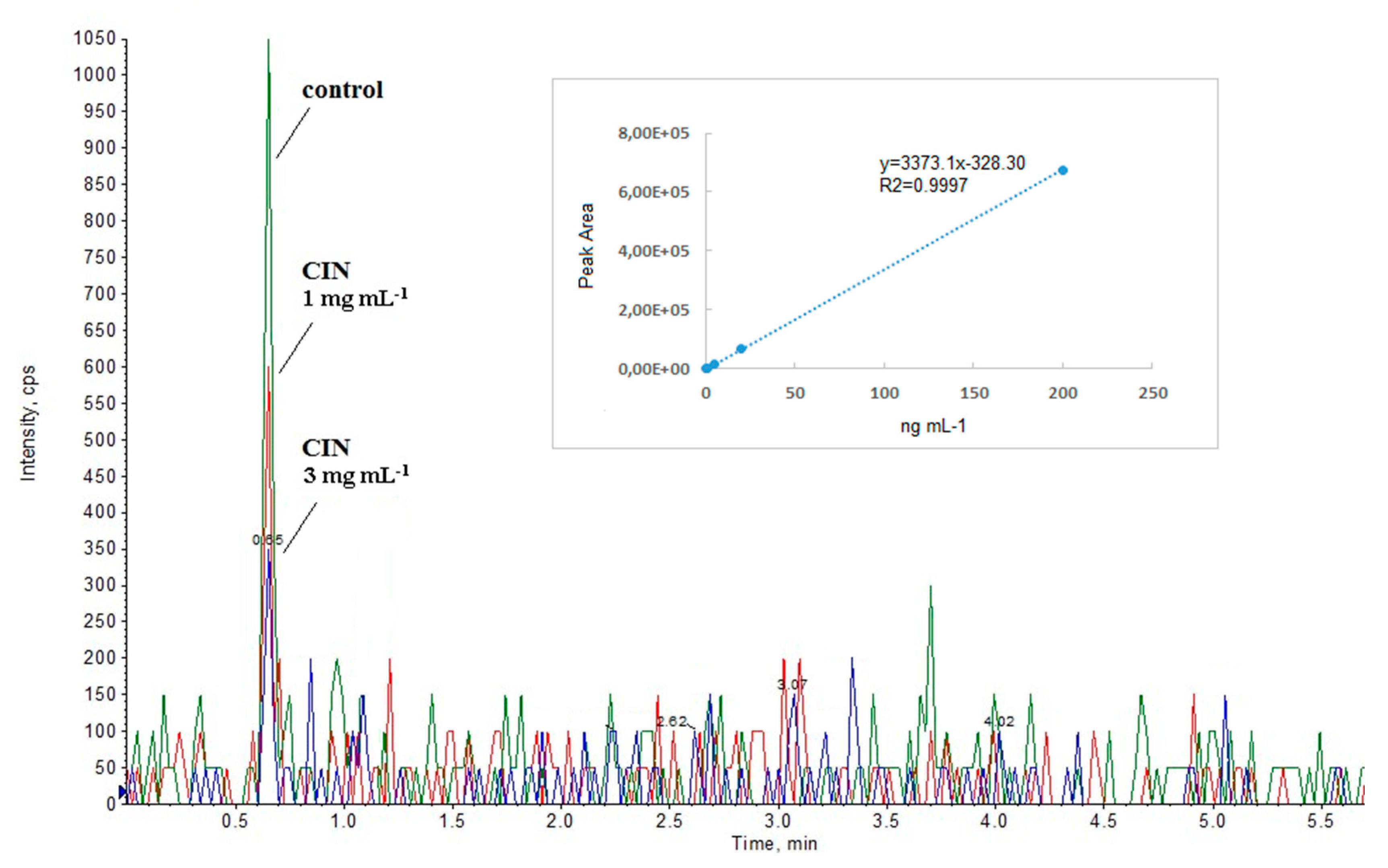
3.1. Identification of the Autoinducer
3.2. A. amylolyticus α-amylase Monitoring in Different Bacterial Growth Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Poli, A.; Esposito, E.; Lama, L.; Orlando, P.; Nicolaus, G.; de Appolonia, F.; Gambacorta, A.; Nicolaus, B. Anoxybacillus amylolyticus sp. nov., a thermophilic amylase producing bacterium isolated from Mount Rittmann (Antarctica). Syst. Appl. Microbiol. 2006, 29, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Fattah, Y.R.; Soliman, N.A.; El-Toukhy, N.M.; El-Gendi, H.; Ahmed, R.S. Production, purification and characterization of thermostable α-amylase produced by Bacillus licheniformis isolate AI20. J. Chem. 2013, 673173. [Google Scholar] [CrossRef] [Green Version]
- Asgher, M.; Asad, M.J.; Rahman, S.U.; Legge, R.L. A thermostable α-amylase from a moderately thermophilic Bacillus subtilis strain for starch processing. J. Food Eng. 2007, 79, 950–955. [Google Scholar] [CrossRef]
- Annamalai, N.; Thavasi, R.; Vij ayalakshmi, S.; Balasubramanian, T. Extraction, purification and characterization of thermostable, alkaline tolerant α-amylase from Bacillus cereus. Indian J. Microbiol. 2011, 51, 424–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, D.; Satyanarayana, T. Bacterial and Archaeal α-Amylases: Diversity and Amelioration of the Desirable Characteristics for Industrial Applications. Front. Microbiol. 2016, 7, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.; Nigam, P.; Soccol, C.R.; Soccol, V.T.; Sing, D.; Mohan, R. Advances in microbial amylases. Biotechnol. Appl. Biochem. 2000, 31, 135–152. [Google Scholar] [CrossRef]
- Finore, I.; Di Donato, P.; Poli, A.; Kirdar, B.; Kasavi, C.; Toksoy Oner, E.; Nicolaus, B.; Lama, L. Use of Agro Waste Biomass for α-Amylase Production by Anoxybacillus amylolyticus: Purification and Properties. J. Microb. Biochem. Technol. 2014, 6, 320–326. [Google Scholar]
- Williams, P. Quorum Sensing, communication and cross-kingdom signalling in the bacterial world. Microbiology 2007, 153, 3923–3938. [Google Scholar] [CrossRef] [Green Version]
- Abbamondi, G.R.; De Rosa, S.; Iodice, C.; Tommonaro, G. Cyclic dipeptides produced by marine sponge-associated bacteria as Quorum Sensing signals. Nat. Prod. Commun. 2014, 9, 229–232. [Google Scholar] [CrossRef] [Green Version]
- Tommonaro, G.; Abbamondi, G.R.; Iodice, C.; Tait, K.; De Rosa, S. Diketopiperazines produced by the halophilic archaeon, Haloterrigena hispanica, activate AHL bioreporters. Microb. Ecol. 2012, 63, 490–495. [Google Scholar] [CrossRef]
- Abbamondi, G.R.; Suner, S.; Cutignano, A.; Grauso, L.; Nicolaus, B.; Toksoy Oner, E.; Tommonaro, G. Identification of N-Hexadecanoyl-L-homoserine lactone (C16-AHL) as signal molecule in halophilic bacterium Halomonas smyrnensis AAD6. Ann. Microbiol. 2016, 66, 1329–1333. [Google Scholar] [CrossRef]
- Tommonaro, G.; Abbamondi, G.R.; Toksoy Oner, E.; Nicolaus, B. Investigating the Quorum Sensing system in halophilic bacteria. In Halophiles: Biodiversity and Sustainable Exploitation; Maheshwari, D.K., Saraf, M., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 189–207. [Google Scholar]
- Finore, I.; Lama, L.; Poli, A.; Di Donato, P.; Nicolaus, B. Biotechnology implications of extremophiles as life pioneers and wellspring of valuable biomolecules. In Microbial Factories: Biodiversity, Biopolymers, Bioactive Molecules; Kalia, V.C., Ed.; Springer: New Delhi, India, 2015; Volume 2, pp. 193–216. [Google Scholar]
- Chang, C.Y.; Krishnan, T.; Wang, H.; Chen, Y.; Yin, W.F.; Chong, Y.M.; Tan, L.Y.; Chong, T.M.; Chan, K.G. Non-antibiotic quorum sensing inhibitors acting against N-acyl homoserine lactone synthase as druggable target. Sci. Rep. 2014, 4, 1–8. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bernfeld, P. Amylases α and β. In Methods in Enzymology; Kaplan, N., Colowick, S., Eds.; Academic Press: New York, NY, USA, 1955; Volume 1, pp. 149–158. [Google Scholar]
- Tempé, J.; Petit, A.; Holsters, M.; van Montagu, M.; Schell, J. Thermosensitive step associated with transfer of the Ti plasmid during conjugation: Possible relation to transformation in crown gall. PNAS 1977, 74, 2848–2849. [Google Scholar] [CrossRef] [Green Version]
- Fuqua, C.; Winans, S.C. Conserved cis-acting promoter elements are required for density-dependent transcription of Agrobacterium tumefaciens conjugal transfer genes. J. Bacteriol. 1996, 178, 435–440. [Google Scholar] [CrossRef] [Green Version]
- Malheiro, J.F.; Maillard, J.-Y.; Borges, F.; Simões, M. Evaluation of cinnamaldehyde and cinnamic acid derivatives in microbial growth control. Int. Biodeterior. Biodegrad. 2019, 141, 71–78. [Google Scholar] [CrossRef]
- Llamas, I.; Quesada, E.; Martínez-Cánovas, M.J.; Gronquist, M.; Eberhard, A.; González, J.E. Quorum Sensing in halophilic bacteria: Detection of N-acyl-homoserine lactones in the exopolysaccharide-producing species of Halomonas. Extremophiles 2005, 9, 333–341. [Google Scholar] [CrossRef]
- Tahrioui, A.; Quesada, E.; Llamas, I. The hanR/hanI quorum-sensing system of Halomonas anticariensis, a moderately halophilic bacterium. Microbiology 2011, 157, 3378–3387. [Google Scholar] [CrossRef] [Green Version]
- Poli, A.; Nicolaus, B.; Denizci, A.A.; Yavuzturk, B.; Kazan, D. Halomonas smyrnensis sp. nov., a moderately halophilic, exopolysaccharide-producing bacterium. Int. J. Syst. Evol. Microbiol. 2013, 63, 10–18. [Google Scholar] [CrossRef]
- Rivas, M.; Seeger, M.; Jedlicki, E.; Holmes, D.S. Second acyl homoserine lactone production system in the extreme acidophile Acidithiobacillus ferrooxidans. Appl. Environ. Microbiol. 2007, 73, 3225–3231. [Google Scholar] [CrossRef] [Green Version]
- Paggi, R.A.; Martone, C.B.; Fuqua, C.; De Castro, R.E. Detection of Quorum Sensing signals in the haloalkaliphilic archaeon Natronococcus occultus. FEMS Microbiol. Lett. 2003, 221, 49–52. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Zhang, F.; Ding, G.; Li, J.; Guo, X.; Zhu, J.; Zhou, L.; Cai, S.; Liu, X.; Luo, Y.; et al. Acyl homoserine lactone-based Quorum Sensing in a methanogenic archaeon. ISME J. 2012, 6, 1336–1344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Megaw, J.; Gilmore, B.F. Archaeal persisters: Persister cell formation as a stress response in Haloferax volcanii. Front. Microbiol. 2017, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.R.; Montero, C.I.; Conners, S.B.; Shockley, K.R.; Bridger, S.L.; Kelly, R.M. Population density-dependent regulation of exopolysaccharide formation in the hyperthermophilic bacterium Thermotoga maritima. Mol. Microbiol. 2005, 55, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Nichols, J.D.; Johnson, M.R.; Chou, C.-J.; Kelly, R.M. Temperature, not LuxS, mediates AI-2 formation in hydrothermal habitats. FEMS Microbiol. Ecol. 2009, 68, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Shulami, S.; Shenker, O.; Langut, Y.; Lavid, N.; Gat, O.; Zaide, G.; Zehavi, A.; Sonenshein, A.L.; Shoham, Y. Multiple regulatory mechanisms control the expression of the Geobacillus stearothermophilus gene for extracellular xylanase. J. Biol. Chem. 2014, 289, 25957–25975. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Rodríguez, I.; Bolognini, M.; Ricci, J.; Bini, E.; Vetriani, C. From deep-sea volcanoes to human pathogens: A conserved quorum-sensing signal in Epsilon proteobacteria. ISME J. 2015, 9, 1222–1234. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; He, J.; Liu, J.; Yu, H.; Tang, J.; Zhang, J. Role of N-acyl-homoserine lactone (AHL) based quorum sensing on biofilm formation on packing media in wastewater treatment process. RSC Adv. 2016, 6, 11128. [Google Scholar] [CrossRef]
- Basaran, T.I.; Berber, D.; Gökalsın, B.; Tramice, A.; Tommonaro, G.; Abbamondi, G.R.; Erginer Hasköylü, M.; Toksoy Öner, E.; Iodice, C.; Sesal, N.C. Extremophilic Natrinema versiforme against Pseudomonas aeruginosa Quorum Sensing and Biofilm. Front. Microbiol. 2020, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Abbamondi, G.R.; Kambourova, M.; Poli, A.; Finore, I.; Nicolaus, B. Quorum Sensing in Extremophiles. In Quorum Sensing; Tommonaro, G., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 4, pp. 97–123. [Google Scholar]
- Cutignano, A. Analytical Approaches for the Identification of Quorum Sensing Molecules. In Quorum Sensing; Tommonaro, G., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 2, pp. 29–53. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tramice, A.; Cutignano, A.; Iodice, A.; Poli, A.; Finore, I.; Tommonaro, G. Involvement of a Quorum Sensing Signal Molecule in the Extracellular Amylase Activity of the Thermophilic Anoxybacillus amylolyticus. Microorganisms 2021, 9, 819. https://doi.org/10.3390/microorganisms9040819
Tramice A, Cutignano A, Iodice A, Poli A, Finore I, Tommonaro G. Involvement of a Quorum Sensing Signal Molecule in the Extracellular Amylase Activity of the Thermophilic Anoxybacillus amylolyticus. Microorganisms. 2021; 9(4):819. https://doi.org/10.3390/microorganisms9040819
Chicago/Turabian StyleTramice, Annabella, Adele Cutignano, Annalaura Iodice, Annarita Poli, Ilaria Finore, and Giuseppina Tommonaro. 2021. "Involvement of a Quorum Sensing Signal Molecule in the Extracellular Amylase Activity of the Thermophilic Anoxybacillus amylolyticus" Microorganisms 9, no. 4: 819. https://doi.org/10.3390/microorganisms9040819
APA StyleTramice, A., Cutignano, A., Iodice, A., Poli, A., Finore, I., & Tommonaro, G. (2021). Involvement of a Quorum Sensing Signal Molecule in the Extracellular Amylase Activity of the Thermophilic Anoxybacillus amylolyticus. Microorganisms, 9(4), 819. https://doi.org/10.3390/microorganisms9040819