
Polyphenols Content and In Vitro ?-Glycosidase Activity of Different Italian Monofloral Honeys, and Their Effect on Selected Pathogenic and Probiotic Bacteria

 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biochemical Analysis
2.1.1. Determination of Total Polyphenols Content
2.1.2. α-Glycosidase Inhibition Assay
2.2. Antibacterial Properties of the Honeys
2.2.1. Microorganisms and Culture Conditions
2.2.2. Minimal Inhibitory Concentration (MIC)
2.2.3. Biofilm Inhibitory Action of the Honeys
2.2.4. Effect of the Honeys on the Bacterial Adhesion Ability
2.2.5. Action of the Honeys on Mature Bacterial Biofilm
2.2.6. Metabolic Activity of Biofilm Cells
2.3. Effect of the Honeys of the Growth of Probiotics
2.4. Statistical Analysis
3. Results and Discussion
3.1. Total Polyphenol Content
3.2. α-Glycosidase Inhibitory Activity
3.3. Antibacterial Activity
3.4. Activity of Honeys on Biofilm
3.4.1. Activity of Honeys on Biofilm Formation and Bacterial Metabolism
3.4.2. Inhibitory Action of Honeys on the In Vitro Bacterial Adhesion and Bacterial Metabolism
3.4.3. Action of Honey on Mature Biofilm
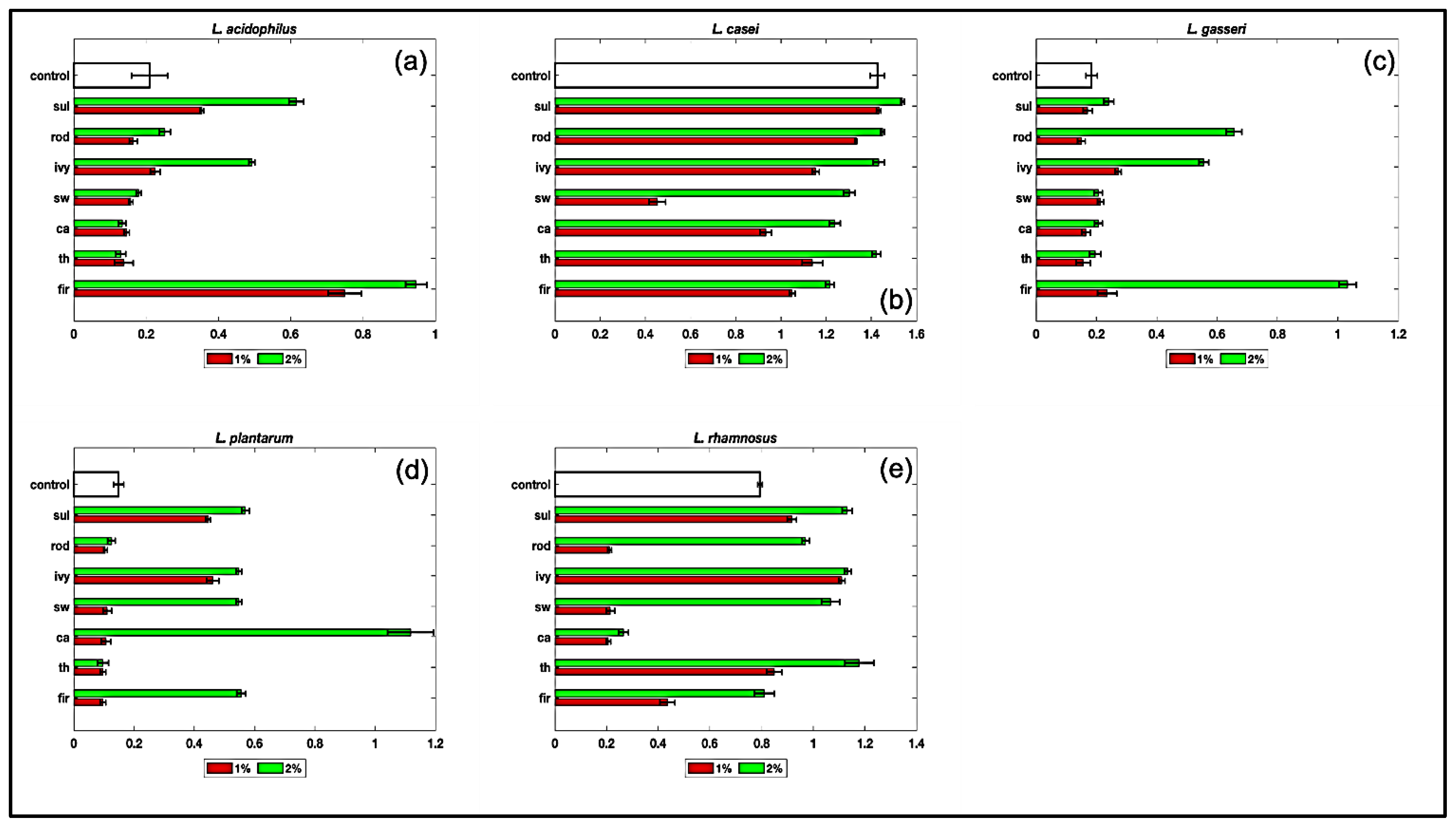
3.5. Effect of Monofloral Honeys on Probiotics Growth
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, J.R.; Ye, Y.L.; Lin, T.Y.; Wang, Y.W.; Peng, C.C. Effect of floral sources on the antioxidant, antimicrobial, and anti-inflammatory activities of honeys in Taiwan. Food Chem. 2013, 139, 938–943. [Google Scholar] [CrossRef]
- Available online: http://www.fao.org/faolex/results/details/en/c/LEX-FAOC108831/ (accessed on 24 May 2021).
- Available online: www.ismea.it (accessed on 24 May 2021).
- Eteraf-Oskouei, T.; Najafi, M. Traditional and modern uses of natural honey in human diseases: A review. Iran. J. Basic Med. Sci. 2013, 16, 731–742. [Google Scholar]
- Afroz, R.; Tanvir, E.M.; Zheng, W.; Little, P.J. Molecular Pharmacology of Honey. Clin. Exp. Pharmacol. 2016, 6, 3. [Google Scholar]
- Weston, R.J. The contribution of catalase and other natural products to the antibacterial activity of honey: A review. Food Chem. 2000, 71, 235–239. [Google Scholar] [CrossRef]
- Bogdanov, S.; Jurendic, T.; Sieber, R.; Gallmann, P. Honey for nutrition and health: A review. J. Am. Coll. Nutr. 2008, 27, 677–689. [Google Scholar] [CrossRef]
- Nasrolahi, O.; Heidari, R.; Rahmani, F.; Farokhi, F. Effect of natural honey from Ilam and metformin for improving glycemic control in streptozotocin-induced diabetic rats. Avicenna J. Phytomed. 2012, 2, 212–221. [Google Scholar] [PubMed]
- Atayoğlu, A.T.; Soylu, M.; Silici, S.; İnanç, N. Glycemic index values of monofloral Turkish honeys and the effect of their consumption on glucose metabolism. Turk. J. Med. Sci. 2016, 46, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Rakhman, A.; Hadju, V.; Bahar, B. Content of nutritional honey Moringa oleifera nutrition & glycemic index value on pregnancy rats (Rattus norvegicus). Hasanuddin Int. J. Health Res. 2020, 1, 47–57. [Google Scholar]
- Gourdomichali, T.; Papakonstantinou, E. Short-term effects of six Greek honey varieties on glycemic response: A randomized clinical trial in healthy subjects. Eur. J. Clin. Nutr. 2018, 72, 1709–1716. [Google Scholar] [CrossRef]
- Zaidi, H.; Ouchemoukh, S.; Amessis-Ouchemoukh, N.; Debbache, N.; Pacheco, R.; Serralheiro, M.L.; Araujo, M.E. Biological properties of phenolic compound extracts in selected Algerian honeys—The inhibition of acetylcholinesterase and α-glucosidase activities. Eur. J. Integr. Med. 2019, 25, 77–84. [Google Scholar] [CrossRef]
- Parada, J.; Santos, J.L.; Cañoles, P.; Manquián, N. Phenolic content of honey reduces in vitro starch digestibility. Czech J. Food Sci. 2016, 34, 217–223. [Google Scholar]
- Cianciosi, D.; Forbes-Hernández, T.Y.; Afrin, S.; Gasparrini, M.; Reboredo-Rodriguez, P.; Manna, P.P.; Zhang, J.; Bravo Lamas, L.; Florez, S.M.; Agudo Toyos, P.A.; et al. Phenolic compounds in honey and their associated health benefits: A review. Molecules 2018, 23, 2322. [Google Scholar] [CrossRef] [Green Version]
- Nazzaro, F.; Fratianni, F.; d’Acierno, A.; De Feo, V.; Ayala-Zavala, F.J.; Cruz, A.G.; Granato, D.; Coppola, R. Effect of polyphenols on microbial cell-cell communications. In Quorum Sensing; Tommonaro, G., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 195–223. [Google Scholar]
- Lewis, K. Riddle of biofilm resistance. Antimicr. Agentser Chemoth. 2001, 45, 999–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merckoll, P.; Jonassen, T.O.; Vad, M.E.; Jeansson, S.L.; Melby, K.K. Bacteria, biofilm and honey: A study of the effects of honey on ‘planktonic’ and biofilm-embedded chronic wound bacteria. Scand. J. Infect. Dis. 2009, 41, 341–347. [Google Scholar] [CrossRef]
- Nassar, H.M.; Li, M.; Gregory, R.L. Effect of honey on Streptococcus mutans growth and biofilm formation. Appl. Environ. Microbiol. 2012, 78, 536–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majtan, J.; Bohova, J.; Horniackova, M.; Klaudiny, J.; Majtan, V. Anti-biofilm Effects of honey against wound pathogens Proteus mirabilis and Enterobacter cloacae. Phytother. Res. 2014, 28, 69–75. [Google Scholar] [CrossRef]
- Stojkovska, J.; Petrovic, P.; Jancic, I.; Milenkovic, M.T.; Obradovic, B. Novel nano-composite hydrogels with honey effective against multi-resistant clinical strains of Acinetobacter baumannii and Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2019, 103, 8529–8543. [Google Scholar] [CrossRef]
- Jervis-Bardy, J.; Foreman, A.; Bray, S.; Tan, L.; Wormald, P.J. Methylglyoxal-infused honey mimics the anti-Staphylococcus aureus biofilm activity of Manuka honey: Potential implication in chronic rhinosinusitis. Laryngoscope 2011, 121, 1104–1107. [Google Scholar] [CrossRef]
- Tze Tan, H.T.; Rahman, R.A.; Gan, S.H.; Halim, A.S.; Hassan, S.A.; Sulaiman, S.A.; Kirnpal-Kaur, B.S. The antibacterial properties of Malaysian tualang honey against wound and enteric microorganisms in comparison to manuka honey. BMC Compl. Altern. Med. 2009, 9, 34. [Google Scholar]
- Celebioglu, H.U. Probiotic bacteria grown with chestnut honey enhance in vitro cytotoxicity on breast and colon cancer cells. Arch. Biol. Sci. 2020, 72, 329–338. [Google Scholar] [CrossRef]
- Celebioglu, H.U.; Erden, Y.; Ozel, H.B. In vitro cytotoxic effects of lactobacilli grown with lime honey on human breast and colon cancer cells. Food Biosci. 2021, 41, 101020. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; Cozzolino, R.; Martignetti, A.; Malorni, L.; De Feo, V.; Cruz, A.G.; d’Acierno, A. Antibacterial activity of three extra virgin olive oils of the Campania Region, Southern Italy, related to their polyphenol content and composition. Microorganisms 2019, 7, 321. [Google Scholar] [CrossRef] [Green Version]
- Sharp, H.; Hollinshead, J.; Bartholomew, B.B.; Oben, J.; Watson, A.; Nash, R.J. Inhibitory effects of Cissus quadrangularis L. derived components on lipase, amylase and α-glucosidase activity in vitro. Nat. Prod. Commun. 2007, 2, 817–822. [Google Scholar] [CrossRef] [Green Version]
- Watson, A.; Fleet, G.W.J.; Asano, N.; Molyneux, R.J.; Nash, R.J. Polyhydroxylated alkaloids—Natural occurrence and therapeutic applications. Phytochemistry 2001, 56, 265–295. [Google Scholar] [CrossRef]
- Sarker, S.D.; Nahar, L.; Kumarasamy, Y. Microtitre plate-based antibacterial assay incorporating resazurin as an indicator of cell growth, and its application in the in vitro antibacterial screening of phytochemicals. Methods 2007, 42, 321–324. [Google Scholar] [CrossRef]
- Caputo, L.; Amato, G.; Fratianni, F.; Coppola, R.; Candido, V.; De Feo, V.; Nazzaro, F. Chemical characterization and antibiofilm activities of bulbs and leaves of two aglione (Allium ampeloprasum var. holmense Asch. et Graebn.) landraces grown in Southern Italy. Molecules 2020, 25, 5486. [Google Scholar] [CrossRef] [PubMed]
- Kairo, S.K.; Bedwell, J.; Tyler, P.C.; Carter, A.; Corbel, M.J. Development of a tetrazolium salt assay for rapid determination of viability of BCG vaccines. Vaccine 1999, 17, 2423–2428. [Google Scholar] [CrossRef]
- Vela, L.; de Lorenzo, C.; Perez, R.A. Antioxidant capacity of Spanish honeys and its correlation with polyphenol content and other physicochemical properties. J. Sci. Food Agric. 2007, 87, 1069–1075. [Google Scholar] [CrossRef]
- Petretto, G.L.; Cossu, M.; Alamanni, M.C. Phenolic content, antioxidant and physico-chemical properties of Sardinian monofloral honeys. Int. J. Food Sci. Technol. 2015, 50, 482–491. [Google Scholar] [CrossRef]
- Perna, A.; Simonetti, A.; Intaglietta, I.; Sofo, A.; Gambacorta, E. Metal content of southern Italy honey of different botanical origins and its correlation with polyphenol content and antioxidant activity. Int. J. Food Sci. Technol. 2012, 47, 1909–1917. [Google Scholar] [CrossRef]
- Kavanagh, S.; Gunnoo, J.; Passos, T.M.; Stout, J.C.; White, B. Physicochemical properties and phenolic content of honey from different floral origins and from rural versus urban landscapes. Food Chem. 2019, 272, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Pichichero, E.; Canuti, L.; Canini, A. Characterisation of the phenolic and flavonoid fractions and antioxidant power of Italian honeys of different botanical origin. J. Sci. Food Agric. 2009, 89, 609–616. [Google Scholar] [CrossRef]
- Silici, S.; Sagdic, O.; Ekici, L. Total phenolic content, antiradical, antioxidant and antimicrobial activities of rhododendron honeys. Food Chem. 2010, 121, 238–243. [Google Scholar] [CrossRef]
- Gül, A.; Pehlivan, T. Antioxidant activities of some monofloral honey types produced across Turkey. Saudi J. Biol. Sci. 2018, 25, 1056–1065. [Google Scholar] [CrossRef]
- Alvarez-Suarez, J.M.; Tulipani, S.; Díaz, D.; Estevez, Y.; Romandini, S.; Giampieri, F.; Damiani, E.; Astolfi, P.; Bompadre, S.; Battino, M. Antioxidant and antimicrobial capacity of several monofloral Cuban honeys and their correlation with color, polyphenol content and other chemical compounds. Food. Chem. Toxicol. 2010, 48, 2490–2499. [Google Scholar] [CrossRef] [PubMed]
- Ademiluyi, A.O.; Oboh, G. Soybean phenolic-rich extracts inhibit key-enzymes linked to type 2 diabetes (α-amylase and α-glucosidase) and hypertension (angiotensin I converting enzyme) in vitro. Exp. Toxicol. Pathol. 2013, 65, 305–309. [Google Scholar] [CrossRef]
- Sharma, R.; Martins, N.; Chaudhary, A.; Garg, N.; Sharma, V.; Kuca, K.; Nepovimova, E.; Tuli, H.S.; Bishayee, A.; Chaudhary, A.; et al. Adjunct use of honey in diabetes mellitus: A consensus or conundrum? Trends Food. Sci. Technol. 2020, 107, 254–274. [Google Scholar] [CrossRef]
- Ali, H.; Bakar, M.F.A.; Majid, M.; Muhammad, N.; Lim, S.Y. In vitro anti-diabetic activity of stingless bee honey from different botanical origins. Food Res. 2020, 4, 1421–1426. [Google Scholar] [CrossRef]
- Kmieliūtė, K.; Batulevičienė, V. Correlation of botanical origin and quality of honey. Food Chem. Technol. 2014, 48, 29–38. [Google Scholar]
- Krishnasree, V.; Ukkuru, M.P. In vitro antidiabetic activity and glycemic index of bee honeys. Indian J. Tradit. Knowl. 2007, 16, 134–140. [Google Scholar]
- Carnwath, R.; Graham, E.; Reynolds, K.; Pollock, P. The antimicrobial activity of honey against common equine wound bacterial isolates. Vet. J. 2014, 199, 110–114. [Google Scholar] [CrossRef]
- Molan, P.C. The Antibacterial activity of honey. Bee World 1992, 73, 5–28. [Google Scholar] [CrossRef]
- Mandal, M.D.; Mandal, S. Honey: Its medicinal property and antibacterial activity. Asian Pac. J. Trop. Biomed. 2011, 1, 154–160. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Li, Q.X. Chemical composition, characterization, and differentiation of honey botanical and geographical origins. Adv. Food Nutr. Res. 2011, 62, 89–137. [Google Scholar]
- El-Aal, A.; El-Hadidy, M.; El-Mashad, N.; El-Sebaie, A. Antimicrobial effect of bee honey in comparison to antibiotics on organisms isolated from infected burns. Ann. Burns Fire Disasters 2007, 20, 83. [Google Scholar]
- Almasaudi, S.B.; Al-Nahari, A.A.M.; El Sayed, M.; El-Ghany, A.; Barbour, E.; Al Muhayawi, S.M.; Al-Jaouni, S.; Azhar, E.; Qari, M.; Qari, Y.A.; et al. Antimicrobial effect of different types of honey on Staphylococcus aureus. Saudi J. Biol. Sci. 2017, 24, 1255–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogdanov, S. Characterization of antibacterial substances in honey. Lebensm Wiss Technol. 1984, 17, 74–76. [Google Scholar]
- Nazzaro, F.; Fratianni, F.; Coppola, R. Quorum sensing and phytochemicals. Int. J. Mol. Sci. 2013, 14, 12607–12619. [Google Scholar] [CrossRef] [Green Version]
- Stagos, D.; Soulitsiotis, N.; Tsadila, C.; Papaeconomou, S.; Arvanitis, C.; Ntontos, A.; Karkanta, F.; Adamou-Androulaki, S.; Petrotos, K.; Spandidos, D.A.; et al. Antibacterial and antioxidant activity of different types of honey derived from Mount Olympus in Greece. Int. J. Mol. Med. 2018, 42, 726–734. [Google Scholar] [CrossRef] [Green Version]
- Melliou, E.; Chinou, I. Chemical constituents of selected unifloral Greek bee-honeys with antimicrobial activity. Food Chem. 2011, 129, 284–290. [Google Scholar] [CrossRef]
- Broznić, D.; Ratkaj, I.; Staver, M.; Kraljević Pavelić, S.; Žurga, P.; Bubalo, D.; Gobin, I. Evaluation of the Antioxidant Capacity, Antimicrobial and Antiproliferative Potential of Fir (Abies alba Mill.) Honeydew Honey Collected from Gorski kotar (Croatia). Food Technol. Biotechnol. 2018, 56, 533–545. [Google Scholar] [CrossRef]
- Tenesaca, M.; Navarrete, E.S.; Iturralde, G.A.; Villacrés Granda, I.M.; Tejera, E.; Beltrán-Ayala, P.; Giampieri, F.; Battino, M.; Alvarez-Suarez, J.M. Influence of botanical origin and chemical composition on the protective effect against oxidative damage and the capacity to reduce in vitro bacterial biofilms of monofloral honeys from the Andean Region of Ecuador. Int. J. Mol. Sci. 2018, 19, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, C.I.; Aazza, S.; Faleiro, M.L.; Miguel, M.D.G.; Neto, L. The antibacterial, anti-biofilm, anti-inflammatory and virulence inhibition properties of Portuguese honeys. J. Apicul. Res. 2016, 55, 292–304. [Google Scholar] [CrossRef] [Green Version]
- Henriques, A.F.; Jenkins, R.E.; Burton, N.F.; Cooper, R.A. The intracellular effects of manuka honey on Staphylococcus aureus. Eur. J. Clin. Microbiol. Infect. Dis. 2010, 29, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Brudzynski, K.; Sjaarda, C. Antibacterial compounds of Canadian honeys target bacterial cell wall inducing phenotype changes, growth inhibition and cell lysis that resemble action of β-lactam antibiotics. PLoS ONE 2014, 9, e106967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanley, N.R.; Lazazzera, B.A. Environmental signals and regulatory pathways that influence biofilm formation. Mol. Microbiol. 2004, 52, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Longo, F.; Vuotto, C.; Donelli, G. Biofilm formation in Acinetobacter baumannii. New Microbiol. 2014, 37, 119–127. [Google Scholar] [PubMed]
- Donlan, R.M. Biofilms and device-associated infections. Emerg. Infect. Dis. 2001, 7, 277–281. [Google Scholar] [CrossRef]
- Bodey, G.P.; Bolivar, R.; Fainstein, V.; Jadeja, L. Infections caused by Pseudomonas aeruginosa. Rev. Infec. Dis. 1983, 5, 279–313. [Google Scholar] [CrossRef]
- Wolcott, R.D.; Rhoads, D.D.; Dowd, S.E. Biofilms and chronic wound inflammation. J. Wound Care 2008, 17, 333–341. [Google Scholar] [CrossRef]
- Lu, J.; Cokcetin, N.N.; Burke, C.M.; Turnbull, L.; Liu, M.; Carter, D.A.; Whitchurch, C.B.; Harry, E.J. Honey can inhibit and eliminate biofilms produced by Pseudomonas aeruginosa. Sci. Rep. 2019, 9, 18160. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Turnbull, L.; Burke, C.M.; Liu, M.; Carter, D.A.; Schlothauer, R.C.; Whitchurch, C.B.; Elizabeth, J.; Harry, E.J. Manuka-type honeys can eradicate biofilms produced by Staphylococcus aureus strains with different biofilm-forming abilities. Peer J. 2014, 2, e326. [Google Scholar] [CrossRef] [Green Version]
- Bobiş, O.; Dezmirean, D.S.; Moise, A.R. Honey and Diabetes: The Importance of Natural Simple Sugars in Diet for Preventing and Treating Different Type of Diabetes. Oxidative Med. Cell. Longev. 2018, 2018, 4757893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prucek, R.; Panáček, A.; Gajdová, Z.; Večeřová, R.; Kvítek, L.; Gallo, J.; Kolář, M. Specific detection of Staphylococcus aureus infection and marker for Alzheimer disease by surface enhanced Raman spectroscopy using silver and gold nanoparticle-coated magnetic polystyrene beads. Sci. Rep. 2021, 11, 6240. [Google Scholar] [CrossRef]
- Shoemark, D.K.; Allen, S.J. The Microbiome and Disease: Reviewing the Links between the Oral Microbiome, Aging, and Alzheimer’s Disease. J. Alzheimer’s Dis. 2015, 43, 725–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolan, V.C.; Harrison, J.; Cox, J.A.G. Dissecting the antimicrobial composition of honey. Antibiotics 2019, 8, 251. [Google Scholar] [CrossRef] [Green Version]
- Sanz, M.L.; Polemis, N.; Morales, V.; Corzo, N.; Drakoularakou, A.; Gibson, G.R.; Rastall, R.A. In vitro investigation into the potential prebiotic activity of honey oligosaccharides. J. Agric. Food Chem. 2005, 53, 2914–2921. [Google Scholar] [CrossRef] [PubMed]
- Gmeiner, M.; Kneifel, W.; Kulbe, K.D.; Wouters, R.; De Boever, P.; Nollet, L.; Verstraete, W. Influence of a synbiotic mixture consisting of Lactobacillus acidophilus 74-2 and a fructooligosaccharide preparation on the microbial ecology sustained in a simulation of the human intestinal microbial ecosystem (SHIME reactor). Appl. Microb. Biotech. 2000, 53, 219–223. [Google Scholar] [CrossRef]
- Carvalho de Melo, F.H.; Dantas Duarte Menezes, F.N.; de Sousa, J.M.B.; dos Santos Lima, M.; da Silva Campelo Borges, G.; de Souza, E.L.; Magnani, M. Prebiotic activity of monofloral honeys produced by stingless bees in the semi-arid region of Brazilian Northeastern toward Lactobacillus acidophilus LA-05 and Bifidobacterium lactis BB-12. Food Res. Int. 2020, 128, 108809. [Google Scholar] [CrossRef]
- Das, A.; Datta, S.; Mukherjee, S.; Bose, S.; Ghosh, S.; Dhar, P. Evaluation of antioxidative, antibacterial and probiotic growth stimulatory activities of Sesamum indicum honey containing phenolic compounds and lignans. LWT Food Sci. Technol. 2015, 61, 244–250. [Google Scholar] [CrossRef]
- Shamala, T.R.; Shri Jyothi, Y.; Saibaba, P. Stimulatory effect of honey on multiplication of lactic acid bacteria under in vitro and in vivo conditions. Lett. Appl. Microbiol. 2000, 30, 453–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Total Polyphenols μg/g (±SD) | α-Glucosidase Inhibition IC50 mg/mL (±SD) | |
|---|---|---|
| Strawberry tree | 552.29 (±22.18) | 32.7 (±2.4) |
| Tree of heaven | 220.62 (±8.19) | 25.4 (±2.1) |
| Sulla | 182.4 (±11.21) | 20.2 (±2.8) |
| Cardoon | 183.95 (±6.3) | 34.03 (±3.1) |
| Ivy | 257.07 (±8.73) | 1.29 (±4.5) |
| Fir | 386.01(±15.15) | 26.8 (±2.3) |
| Rhododendron | 110.46 (±15.21) | 28.7 (±2.7) |
| A. baumannii | E. coli | L. monocytogenes | P. aeruginosa | S. aureus | |
|---|---|---|---|---|---|
| Cardoon | 30 μL/mL (±3.0) | 35 μL/mL (±3.0) | 20 μL/mL (±3.0) | 25 μL/mL (±5.0) | 30 μL/mL (±2.0) |
| Fir | 25 μL/mL (±2.0) | 35 μL/mL (±2.0) | 20 μL/mL (±2.0) | 35 μL/mL (±3.0) | 20 μL/mL (±2.0) |
| Ivy | 25 μL/mL (±2.0) | 25 μL/mL (±2.0) | 25 μL/mL (±2.0) | 30 μL/mL (±3.0) | 35 μL/mL (±3.0) |
| Rhododendron | 20 μL/mL (±2.0) | 25 μL/mL (±2.0) | 30 μL/mL (±2.0) | 35 μL/mL (±3.0) | >50 μL/mL |
| Strawberry three | 35 μL/mL (±5.0) | 25 μL/mL (±5.0) | 35 μL/mL (±5.0) | 40 μL/mL (±4.0) | 35 μL/mL (±3.0) |
| Sulla | 20 μL/mL (±5.0) | 20 μL/mL (±5.0) | 20 μL/mL (±5.0) | 35 μL/mL (±3.0) | 45 μL/mL (±3.0) |
| Tree of Heaven | 35 μL/mL (±2.0) | 30 μL/mL (±2.0) | 25 μL/mL (±2.0) | 35 μL/mL (±3.0) | 35 μL/mL (±3.0) |
| MTT | CV | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| AB | EC | LM | PS | SA | AB | EC | LM | PS | SA | |
| ca | 21.55 | 59.59 | 0 | 80.37 | 60.66 | 58.16 | 0 | 92.88 | 93.12 | 83.86 |
| 5.71 μL/mL | (±1.03) | (±2.38) | (±0) | (±0.31) | (±0.95) | (±0.98) | (±0) | (±0.22) | (±0.25) | (±2.71) |
| ca | 26.04 | 71.49 | 0 | 81.53 | 62.25 | 82.00 | 15.41 | 93.27 | 93.41 | 88.33 |
| 11.42 μL/mL | (±1.18) | (±1.20) | (±0) | (±0.11) | (±0.30) | (±1.29) | (±1.31) | (±0.19) | (±0.13) | (±0.15) |
| fir | 0 | 49.74 | 0 | 66.10 (±1.35) | 42.87 | 71.42 | 44.00 | 91.54 | 88.00 | 81.90 |
| 5.71 μL/mL | (±0) | (±1.13) | (±0) | (±3.77) | (±3.18) | (±2.05) | (±0.15) | (±0.19) | (±1.46) | |
| fir | 9.52 | 61.85 | 0 | 67.67 | 70.14 | 76.79 | 64.96 | 92.03 | 89.80 | 80.07 |
| 11.42 μL/mL | (±1.24) | (±0.74) | (±0) | (±0.33) | (±1.00) | (±2.80) | (±4.91) | (±0.23) | (±0.10) | (±0.89) |
| ivy | 7.18 | 43.17 | 0 | 73.06 | 44.75 | 65.89 | 0 | 87.71 | 86.26 | 75.66 |
| 5.71 μL/mL | (±1.78) | (±1.42) | (±0) | (±0.56) | (±3.04) | (±1.45) | (±0) | (±0.20) | (±1.02) | (±1.23) |
| ivy | 9.81 | 48.61 | 0 | 71.03 | 40.12 | 72.26 | 54.71 | 89.93 | 87.28 | 78.43 |
| 11.42 μL/mL | (±2.43) | (±1.68) | (±0) | (±2.09) | (±0.72) | (±1.06) | (±1.73) | (±0.18) | (±0.37) | (±0.18) |
| rhod | 18.55 | 8.93 | 0 | 52.91 | 32.65 | 29.15 | 0 | 49.84 | 0 | 0 |
| 5.71 μL/mL | (±0.57) | (±0.37) | (±0) | (±1.33) | (±1.33) | (±1.90) | (±0) | (±2.19) | (±0) | (±0) |
| rhod | 39.37 (±3.21) | 13.98 | 0 | 60.44 | 38.05 | 69.68 | 0 | 65.70 | 14.60 | 0 |
| 11.42 μL/mL | (±0.78) | (±0) | (±0.56) | (±1.05) | (±2.68) | (±0) | (±1.55) | (±1.95) | (±0) | |
| sul | 55.46 | 23.21 | 0 | 57.59 | 38.13 | 16.47 | 41.59 | 56.79 | 0 | 0 |
| 5.71 μL/mL | (±0.93) | (±1.93) | (±0) | (±0.43) | (±0.21) | (±2.19) | (±2.08) | (±0.46) | (±0) | (±0) |
| sul | 66.30 | 44.75 | 0 | 62.61 | 39.03 | 32.12 | 72.92 | 72.29 | 34.99 | 1.54 |
| 11.42 μL/mL | (±0.85) | (±2.00) | (±0) | (±0.21) | (±0.29) | (±1.71) | (±1.70) | (±0.98) | (±7.79) | (±0.46) |
| sw | 6.23 | 55.08 | 0 | 63.12 | 22.98 | 0 | 0 | 57.68 | 9.86 | 0 |
| 5.71 μL/mL | (±2.29) | (±1.26) | (±0) | (±0.27) | (±1.00) | (±0) | (±0) | (±0.60) | (±1.92) | (±0) |
| sw | 16.05 | 69.72 | 0 | 64.32 | 24.89 | 72.19 | 21.51 | 61.00 | 10.29 | 20.80 |
| 11.42 μL/mL | (±1.10) | (±1.11) | (±0) | (±0.30) | (±0.23) | (±3.11) | (±1.68) | (±0.69) | (±1.02) | (±2.35) |
| th | 27.07 | 49.88 | 0 | 61.73 | 27.11 | 26.38 | 6.98 | 65.83 | 0 | 1.05 |
| 5.71 μL/mL | (±1.32) | (±0.84) | (±0) | (±0.74) | (±0.79) | (±1.41) | (±2.20) | (±0.03) | (±0) | (±0.62) |
| th | 31.73 | 73.87 | 16.71 | 75.23 | 61.64 | 49.31 | 38.92 | 66.08 | 49.14 | 26.13 |
| 11.42 μL/mL | (±0.87) | (±1.10) | (±3.13) | (±0.28) | (±1.70) | (±2.14) | (±3.77) | (±0.02) | (±0.32) | (±0.39) |
| MTT | CV | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| AB | EC | LM | PS | SA | AB | EC | LM | PS | SA | |
| ca | 2.83 | 8.88 | 50.93 | 0 | 50.22 | 0 | 54.95 | 50.97 | 21.30 | 36.68 |
| 5.71 μL/mL | (±0.18) | (±1.53) | (±0.80) | (±0) | (±1.52) | (±0) | (±1.35) | (±0.66) | (±1.89) | (±0.75) |
| ca | 44.97 | 43.48 | 53.73 | 15.73 | 55.21 | 19.95 | 52.20 | 68.05 | 37.30 | 55.14 |
| 11.42 μL/mL | (±1.25) | (±1.19) | (±1.52) | (±1.31) | (±2.71) | (±1.99) | (±5.45) | (±2.36) | (±0.80) | (±0.90) |
| fir | 3.76 | 0 | 44.37 | 0 | 40.29 | 31.50 | 30.52 | 58.13 | 46.59 | 51.29 |
| 5.71 μL/mL | (±1.04) | (±0) | (±2.40) | (±0) | (±1.69) | (±1.77) | (±1.70) | (±0.77) | (±0.83) | (±1.73) |
| fir | 40.53 | 49.16 | 60.98 | 41.37 | 56.81 | 33.69 | 43.06 | 68.30 | 54.85 | 58.97 |
| 11.42 μL/mL | (±2.43) | (±2.30) | (±3.10) | (±2.15) | (±1.03) | (±2.61) | (±1.54) | (±0.01) | (±1.79) | (±2.17) |
| ivy | 0 | 0 | 34.65 | 0 | 47.18 | 22.33 | 13.97 | 52.07 | 26.00 | 32.97 |
| 5.71 μL/mL | (±0) | (±0) | (±1.49) | (±0) | (±0.83) | (±4.58) | (±2.16) | (±1.20) | (±1.52) | (±1.22) |
| ivy | 0 | 0 | 55.86 | 0 | 49.81 | 32.80 | 40.99 | 70.34 | 41.22 | 51.76 |
| 11.42 μL/mL | (±0) | (±0) | (±0.86) | (±0) | (±0.83) | (±2.59) | (±0.62) | (±0.72) | (±2.81) | (±0.54) |
| rhod | 0 | 0 | 13.25 | 0 | 38.65 | 49.83 | 47.92 | 36.56 | 31.60 | 24.98 |
| 5.71 μL/mL | (±0) | (±0) | (±1.50) | (±0) | (±1.05) | (±1.25) | (±0.64) | (±1.35) | (±0.53) | (±0.66) |
| rhod | 7.16 | 0.11 | 28.30 | 0 | 47.71 | 51.67 | 66.63 | 44.30 | 36.67 | 27.54 |
| 11.42 μL/mL | (±1.51) | (±0.02) | (±1.18) | (±0) | (±1.00) | (±2.50) | (±0.19) | (±1.65) | (±0.75) | (±0.51) |
| sul | 0 | 7.47 | 40.49 | 0 | 37.15 | 44.35 | 55.41 | 50.85 | 47.42 | 44.77 |
| 5.71 μL/mL | (±0) | (±1.38) | (±0.70) | (±0) | (±0.14) | (±3.05) | (±0.88) | (±0.89) | (±0.85) | (±1.24) |
| sul | 13.48 | 22.19 | 42.37 | 0.57 | 45.38 | 51.77 | 66.24 | 71.57 | 52.74 | 49.19 |
| 11.42 μL/mL | (±2.27) | (±2.41) | (±0.39) | (±0.09) | (±1.10) | (±1.20) | (±1.50) | (±0.45) | (±0.70) | (±0.71) |
| sw | 0 | 0 | 40.66 | 0 | 32.87 | 0 | 55.78 | 35.71 | 0 | 40.31 |
| 5.71 μL/mL | (±0) | (±0) | (±1.35) | (±0) | (±2.31) | (±0) | (±2.88) | (±0.79) | (±0) | (±1.18) |
| sw | 2.18 | 7.76 | 42.52 | 0 | 39.82 | 8.42 | 56.64 | 47.47 | 18.91 | 43.81 |
| 11.42 μL/mL | (±0.94) | (±3.60) | (±0.56) | (±0) | (±1.17) | (±1.96) | (±1.00) | (±0.85) | (±1.54) | (±2.95) |
| th | 0 | 4.46 | 28.27 | 0 | 37.50 | 33.52 | 59.39 | 64.33 | 34.81 | 79.46 |
| 5.71 μL/mL | (±0) | (±1.07) | (±0.91) | (±0) | (±1.04) | (±1.51) | (±1.49) | (±0.40) | (±1.27) | (±0.61) |
| th | 23.37 | 25.52 | 46.78 | 0 | 44.42 | 34.87 | 73.66 | 69.49 | 61.56 | 84.27 |
| 11.42 μL/mL | (±1.23) | (±1.43) | (±0.96) | (±0) | (±0.81) | (±2.30) | (±1.96) | (±0.85) | (±1.73) | (±1.18) |
| MTT | CV | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| AB | EC | LM | PS | SA | AB | EC | LM | PS | SA | |
| ca | 8.44 | 0 | 0 | 36.10 | 24.93 | 1.27 | 0 | 33.71 | 10.96 | 35.62 |
| 5.71 μL/mL | (±0.96) | (±0) | (±0) | (±2.74) | (±2.13) | (±0.74) | (±0) | (±0.88) | (±0.88) | (±1.42) |
| ca | 13.97 | 0 | 0 | 11.66 | 41.06 | 8.66 | 0 | 53.84 | 19.94 | 40.33 |
| 11.42 μL/mL | (±1.75) | (±0) | (±0) | (±1.03) | (±1.76) | (±8.66) | (±0) | (±1.13) | (±0.28) | (±0.56) |
| fir | 0 | 0 | 0 | 49.00 | 26.05 | 21.31 | 0 | 37.08 | 32.46 | 33.34 |
| 5.71 μL/mL | (±0) | (±0) | (±0) | (±1.38) | (±1.53) | (±1.07) | (±0) | (±1.61) | (±1.03) | (±0.98) |
| fir | 85.19 | 0 | 42.40 | 70.76 | 44.29 | 23.95 | 0 | 52.43 | 34.41 | 38.21 |
| 11.42 μL/mL | (±1.39) | (±0) | (±3.83) | (±0.68) | (±2.70) | (±0.31) | (±0) | (±0.73) | (±0.50) | (±0.66) |
| ivy | 0 | 0 | 44.48 | 30.84 | 25.17 | 6.70 | 0 | 33.86 | 42.14 | 44.59 |
| 5.71 μL/mL | (±0) | (±0) | (±1.49) | (±1.50) | (±1.07) | (±2.12) | (±0) | (±0.70) | (±1.37) | (±0.68) |
| ivy | 6.66 | 0 | 53.80 | 38.05 | 43.89 | 22.30 | 0 | 50.98 | 44.54 | 64.96 |
| 11.42 μL/mL | (±0.70) | (±0) | (±2.01) | (±1.61) | (±2.62) | (±1.34) | (±0) | (±0.57) | (±1.72) | (±0.40) |
| rhod | 41.57 | 0 | 0 | 0 | 0 | 10.62 | 0 | 31.83 | 20.22 | 32.38 |
| 5.71 μL/mL | (±0.68) | (±0) | (±0) | (±0) | (±0) | (±1.63) | (±0) | (±2.89) | (±0.87) | (±0.38) |
| rhod | 45.27 | 0 | 23.16 | 37.66 | 1.10 | 16.58 | 0 | 44.83 | 28.13 | 34.77 |
| 11.42 μL/mL | (±1.15) | (±0) | (±2.33) | (±1.59) | (±0.09) | (±1.01) | (±0) | (±1.09) | (±0.54) | (±0.81) |
| sul | 5.90 | 0 | 27.17 | 0 | 0 | 11.43 | 0 | 36.56 | 41.77 | 45.13 |
| 5.71 μL/mL | (±0.97) | (±0) | (±1.64) | (±0) | (±0) | (±2.23) | (±0) | (±0.32) | (±1.50) | (±0.69) |
| sul | 31.28 | 0 | 36.45 | 19.50 | 46.23 | 23.04 | 0 | 37.47 | 44.98 | 48.15 |
| 11.42 μL/mL | (±1.65) | (±0) | (±2.49) | (±3.13) | (±1.17) | (±0.76) | (±0) | (±0.19) | (±0.70) | (±2.47) |
| sw | 0 | 0 | 0 | 21.86 | 30.83 | 2.65 | 0 | 33.88 | 23.45 | 32.87 |
| 5.71 μL/mL | (±0) | (±0) | (±0) | (±1.58) | (±2.21) | (±0.31) | (±0) | (±0.49) | (±1.85) | (±0.52) |
| sw | 9.02 | 0 | 0 | 28.81 | 55.47 | 11.87 | 0 | 54.10 | 51.08 | 37.24 |
| 11.42 μL/mL | (±1.72) | (±0) | (±0) | (±0.93) | (±3.53) | (±1.40) | (±0) | (±1.20) | (±1.10) | (±0.91) |
| th | 18.78 | 0 | 0 | 27.40 | 37.84 | 23.54 | 0 | 11.84 | 25.70 | 46.85 |
| 5.71 μL/mL | (±3.06) | (±0) | (±0) | (±3.94) | (±1.86) | (±0.42) | (±0) | (±1.22) | (±0.57) | (±0.93) |
| th | 57.95 | 0 | 34.28 | 66.98 | 43.78 | 27.51 | 0 | 30.41 | 26.89 | 76.64 |
| 11.42 μL/mL | (±0.71) | (±0) | (±4.91) | (±6.79) | (±0.64) | (±0.95) | (±0) | (±1.89) | (±0.45) | (±0.68) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fratianni, F.; Ombra, M.N.; d’Acierno, A.; Caputo, L.; Amato, G.; De Feo, V.; Coppola, R.; Nazzaro, F. Polyphenols Content and In Vitro ?-Glycosidase Activity of Different Italian Monofloral Honeys, and Their Effect on Selected Pathogenic and Probiotic Bacteria. Microorganisms 2021, 9, 1694. https://doi.org/10.3390/microorganisms9081694
Fratianni F, Ombra MN, d’Acierno A, Caputo L, Amato G, De Feo V, Coppola R, Nazzaro F. Polyphenols Content and In Vitro ?-Glycosidase Activity of Different Italian Monofloral Honeys, and Their Effect on Selected Pathogenic and Probiotic Bacteria. Microorganisms. 2021; 9(8):1694. https://doi.org/10.3390/microorganisms9081694
Chicago/Turabian StyleFratianni, Florinda, Maria Neve Ombra, Antonio d’Acierno, Lucia Caputo, Giuseppe Amato, Vincenzo De Feo, Raffaele Coppola, and Filomena Nazzaro. 2021. "Polyphenols Content and In Vitro ?-Glycosidase Activity of Different Italian Monofloral Honeys, and Their Effect on Selected Pathogenic and Probiotic Bacteria" Microorganisms 9, no. 8: 1694. https://doi.org/10.3390/microorganisms9081694
APA StyleFratianni, F., Ombra, M. N., d’Acierno, A., Caputo, L., Amato, G., De Feo, V., Coppola, R., & Nazzaro, F. (2021). Polyphenols Content and In Vitro ?-Glycosidase Activity of Different Italian Monofloral Honeys, and Their Effect on Selected Pathogenic and Probiotic Bacteria. Microorganisms, 9(8), 1694. https://doi.org/10.3390/microorganisms9081694