Changes in the Mitochondrial Dynamics and Functions Together with the mRNA/miRNA Network in the Heart Tissue Contribute to Hypoxia Adaptation in Tibetan Sheep

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Sample Collection
2.3. Measurement of Blood Physiological and Biochemical Indexes
2.4. Hematoxylin and Eosin Staining
2.5. RNA Extraction
2.6. Library Construction and Sequencing for mRNA and miRNA
2.7. mRNA Identification
2.8. miRNA Identification
2.9. Functional Annotation of DEmRNAs
2.10. Association Analysis of DEmRNAs-DEmiRNAs
2.11. qPCR Verification
2.12. Expression Analysis of Mitochondrial Dynamic-Related Genes
2.13. Analysis of the Expression of Mitochondrial Dynamic-Related Proteins
2.14. Ultrastructural Observation of the Mitochondria
2.15. Mitochondrial Functional Analysis
2.16. Statistical Analysis
3. Results
3.1. Analyses of Blood Physiological and Biochemical Indices
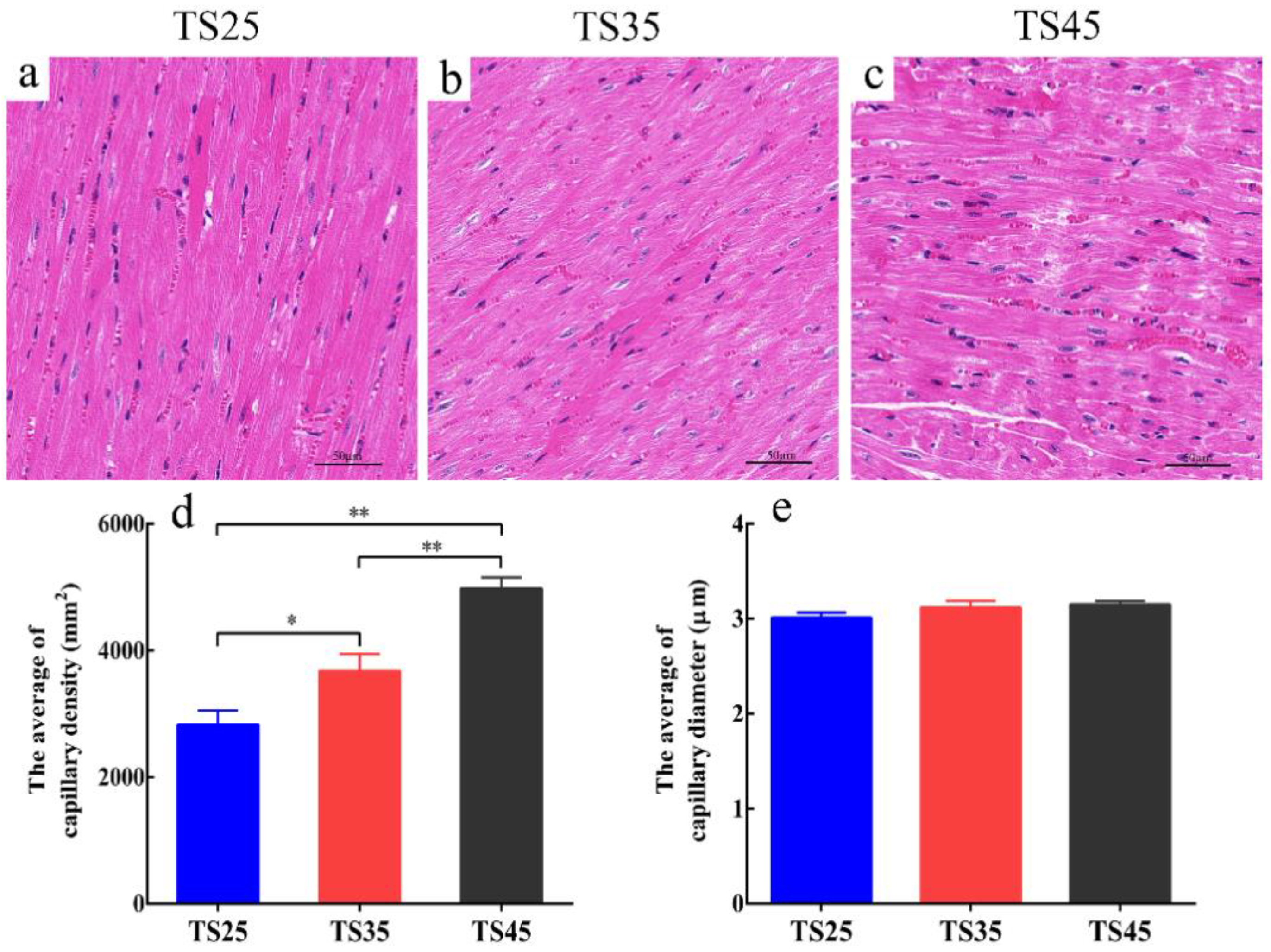
3.2. Analyses of the Structure of the Cardiac Tissues
3.3. Identification of the DEmRNAs in the Hearts of the Tibetan Sheep
3.4. Identification of DEmiRNAs in the Hearts of the Tibetan Sheep
3.5. Functional Analysis of the DEmRNAs
3.6. Hypoxia Related DEmRNA–DEmiRNA Association Analysis and Functional Analysis of the Target Genes
3.7. qPCR Analysis of the Mitochondrial Dynamic-Related Genes
3.8. Immunohistochemical Analysis of Mitochondrial Dynamic-Related Proteins
3.9. Ultrastructural Observation of the Mitochondria
3.10. Mitochondrial Function Analysis
4. Discussion
4.1. The Changes in Blood Indices Promote the Efficiency of Oxygen Transport
4.2. Adaptive Regulation of mRNAs and miRNAs under Hypoxia
4.3. Mitochondrial Fission Increased and Fusion Decreased under Hypoxia
4.4. TCA Cycle Is Down-Regulated and OXPHOS Is Up-Regulated under Hypoxia
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beall, C.M. Two routes to functional adaptation: Tibetan and Andean high-altitude natives. Proc. Natl. Acad. Sci. USA 2007, 104, 8655–8660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kluckova, K.; Tennant, D.A. Metabolic implications of hypoxia and pseudohypoxia in pheochromocytoma and paraganglioma. Cell Tissue Res. 2018, 372, 367–378. [Google Scholar] [CrossRef] [Green Version]
- He, Y.Y.; Munday, J.S.; Perrott, M.; Wang, G.; Liu, X. Association of Age with the Expression of Hypoxia-Inducible Factors HIF-1α, HIF-2α, HIF-3α and VEGF in Lung and Heart of Tibetan Sheep. Animals 2019, 9, 673. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.L.; Lu, X.L.; Cheng, Z.P.; Tian, M.F.; Qiangba, Y.Z.; Fu, Q.; Ren, Z.L. Comparative proteomic analysis of Tibetan pig spermatozoa at high and low altitudes. BMC Genom. 2019, 20, 569. [Google Scholar] [CrossRef]
- Xin, G.S.; Long, R.J.; Guo, X.S.; Irvine, J.; Ding, L.M.; Ding, L.L.; Shang, Z.H. Blood mineral status of grazing Tibetan sheep in the Northeast of the Qinghai—Tibetan Plateau. Livest. Sci. 2011, 136, 102–107. [Google Scholar] [CrossRef]
- Ni, W.J.; Leng, X.M. miRNA-Dependent Activation of mRNA Translation. MicroRNA 2016, 5, 83–86. [Google Scholar] [CrossRef]
- Chen, J.D.; Chen, J.X.; Fu, H.R.; Li, Y.; Wang, L.L.; Luo, S.K.; Lu, H.Y. Hypoxia exacerbates nonalcoholic fatty liver disease via the HIF-2α/PPARα pathway. Am. J. Physiol. Endocrinol. Metab. 2019, 317, E710–E722. [Google Scholar] [CrossRef]
- Xie, Y.B.; Shi, X.F.; Sheng, K.; Han, G.X.; Li, W.Q.; Zhao, Q.Q.; Jiang, B.L.; Feng, J.M.; Li, J.P.; Gu, Y.H. PI3K/Akt signaling transduction pathway, erythropoiesis and glycolysis in hypoxia (Review). Mol. Med. Rep. 2019, 19, 783–791. [Google Scholar] [CrossRef] [Green Version]
- Li, L.J.; Jiang, D.M. Hypoxia-responsive miRNA-21-5p inhibits Runx2 suppression by targeting SMAD7 in MC3T3-E1 cells. J. Cell. Biochem. 2019, 120, 16867–16875. [Google Scholar] [CrossRef] [Green Version]
- Ren, S.S.; Liu, J.Z.; Feng, Y.; Li, Z.Y.; He, L.; Li, L.L.; Cao, X.Z.; Wang, Z.H.; Zhang, Y.W. Knockdown of circDENND4C inhibits glycolysis, migration and invasion by up-regulating miR-200b/c in breast cancer under hypoxia. J. Exp. Clin. Cancer Res. 2019, 38, 388. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Chen, J.Q.; Cheng, Y.H.; Fu, Y.F.; Zhao, H.Z.; Tang, M.Y.; Zhao, H.; Lin, N.; Shi, X.H.; Lei, Y.; et al. Mesenchymal stem cell-derived exosomes protect beta cells against hypoxia-induced apoptosis via miR-21 by alleviating ER stress and inhibiting p38 MAPK phosphorylation. Stem. Cell. Res. Ther. 2020, 11, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, Y.L.; Hu, J.; Wang, J.Q.; Liu, X.; Li, S.B.; Luo, Y.Z. Effect of glycolysis and heat shock proteins on hypoxia adaptation of Tibetan sheep at different altitude. Gene 2021, 803, 145893. [Google Scholar] [CrossRef] [PubMed]
- Kramer, P.; Bressan, P. Our (Mother’s) Mitochondria and Our Mind. Perspect. Psychol. Sci. 2018, 13, 88–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotiadis, V.N.; Duchen, M.R.; Osellame, L.D. Mitochondrial quality control and communications with the nucleus are important in maintaining mitochondrial function and cell health. Biochim. Biophys. Acta 2014, 1840, 1254–1265. [Google Scholar] [CrossRef] [Green Version]
- Lopez, J.; Tait, S.W.G. Mitochondrial apoptosis: Killing cancer using the enemy within. Br. J. Cancer 2015, 112, 957–962. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; He, Y.Y.; Luo, Y.Z. Expression of OPA1 and Mic60 genes and their association with mitochondrial cristae morphology in Tibetan sheep. Cell Tissue Res. 2019, 376, 273–279. [Google Scholar] [CrossRef]
- Yu, R.; Jin, S.B.; Lendahl, U.; Nistér, M.; Zhao, J. Human Fis1 regulates mitochondrial dynamics through inhibition of the fusion machinery. EMBO J. 2019, 38, e99748. [Google Scholar] [CrossRef]
- Horbay, R.; Bilyy, R. Mitochondrial dynamics during cell cycling. Apoptosis 2016, 21, 1327–1335. [Google Scholar] [CrossRef]
- Archer, S.L. Mitochondrial dynamics—Mitochondrial fission and fusion in human diseases. N. Engl. J. Med. 2013, 369, 2236–2251. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.N.; Yuan, H.N.; Yang, T.L.; Li, Y.Q.; Gao, C.X.; Jiao, T.; Cai, Y.; Zhao, S.G. The Expression Regulatory Network in the Lung Tissue of Tibetan Pigs Provides Insight Into Hypoxia-Sensitive Pathways in High-Altitude Hypoxia. Front. Genet. 2021, 12, 691592. [Google Scholar] [CrossRef]
- Chen, S.F.; Zhou, Y.Q.; Chen, Y.R.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Ostell, J.; Pruitt, K.D.; Sayers, E.W. GenBank. Nucleic Acids Res. 2018, 46, D41–D47. [Google Scholar] [CrossRef] [Green Version]
- Griffiths-Jones, S.; Bateman, A.; Marshall, M.; Khanna, A.; Eddy, S.R. Rfam: An RNA family database. Nucleic Acids Res. 2003, 31, 439–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths-Jones, S.; Grocock, R.J.; van Dongen, S.; Bateman, A.; Enright, A.J. miRBase: MicroRNA sequences, targets and gene nomenclature. Nucleic Acids Res. 2006, 34, D140–D144. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.C.; Hsu, C.C.; Wang, J.S. High-Intensity Interval Training Improves Erythrocyte Osmotic Deformability. Med. Sci. Sports Exerc. 2019, 51, 1404–1412. [Google Scholar] [CrossRef] [PubMed]
- Swenson, E.R. Hypoxia and Its Acid-Base Consequences: From Mountains to Malignancy. Adv. Exp. Med. Biol. 2016, 903, 301–323. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Lei, S.Q.; Zhao, B.; Wu, Y.; Su, W.T.; Liu, M.; Meng, Q.T.; Zhou, B.; Leng, Y.; Xia, Z.Y. NLRP3 Inflammasome Activation-Mediated Pyroptosis Aggravates Myocardial Ischemia/Reperfusion Injury in Diabetic Rats. Oxidative Med. Cell. Longev. 2017, 2017, 9743280. [Google Scholar] [CrossRef]
- Darby, I.A.; Hewitson, T.D. Hypoxia in tissue repair and fibrosis. Cell Tissue Res. 2016, 365, 553–562. [Google Scholar] [CrossRef]
- Zheng, J.J.; Que, Q.Y.; Xu, H.T.; Luo, D.S.; Sun, Z.; Ni, J.S.; Que, H.F.; Ma, J.; Wu, D.; Shi, H. Hypoxia Activates SOX5/Wnt/β-Catenin Signaling by Suppressing MiR-338-3p in Gastric Cancer. Technol. Cancer Res. Treat. 2020, 19, 1533033820905825. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.S.; Yin, S.W.; Mao, J.Q.; Liang, F.F.; Zhao, C.; Li, P.; Zhou, G.Q.; Chen, S.Q.; Tang, Z.L. Integrated analysis of mRNA-seq and miRNA-seq in the liver of Pelteobagrus vachelli in response to hypoxia. Sci. Rep. 2016, 6, 22907. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.Q.; Zhu, J.J.; Wang, Y.; Li, Q.; Lin, S. Identification of differentially expressed genes through RNA sequencing in goats (Capra hircus) at different postnatal stages. PLoS ONE 2017, 12, e0182602. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, C.Z.; Wang, N.N.; Li, Z.P.; Heller, R.; Liu, R.; Zhao, Y.; Han, J.G.; Pan, X.Y.; Zheng, Z.Q.; et al. Genetic basis of ruminant headgear and rapid antler regeneration. Science 2019, 364, eaav6335. [Google Scholar] [CrossRef]
- Hermkens, D.M.A.; Stam, O.C.G.; de Wit, N.M.; Fontijn, R.D.; Jongejan, A.; Moerland, P.D.; Mackaaij, C.; Waas, I.S.E.; Daemen, M.; de Vries, H.E. Profiling the unique protective properties of intracranial arterial endothelial cells. Acta Neuropathol. Commun. 2019, 7, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonson, B.; Subramanya, V.; Chan, M.C.; Zhang, A.; Franchino, H.; Ottaviano, F.; Mishra, M.K.; Knight, A.C.; Hunt, D.; Ghiran, I.; et al. DDiT4L promotes autophagy and inhibits pathological cardiac hypertrophy in response to stress. Sci. Signal. 2017, 10, eaaf5967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.Q.; Shi, P.F.; Zhao, G.C.; Xu, J.; Peng, W.; Zhang, J.Y.; Zhang, G.H.; Wang, X.W.; Dong, Z.; Chen, F.; et al. Targeting cancer stem cell pathways for cancer therapy. Signal Transduct. Target. Ther. 2020, 5, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, K.L.; Lai, E.C. Adult-specific functions of animal microRNAs. Nat. Rev. Genet. 2013, 14, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Sotgia, F.; Lisanti, M.P. Mitochondrial biomarkers predict tumor progression and poor overall survival in gastric cancers: Companion diagnostics for personalized medicine. Oncotarget 2017, 8, 67117–67128. [Google Scholar] [CrossRef] [Green Version]
- Kaplon, J.; Zheng, L.; Meissl, K.; Chaneton, B.; Selivanov, V.A.; Mackay, G.; van der Burg, S.H.; Verdegaal, E.M.; Cascante, M.; Shlomi, T.; et al. A key role for mitochondrial gatekeeper pyruvate dehydrogenase in oncogene-induced senescence. Nature 2013, 498, 109–112. [Google Scholar] [CrossRef]
- Ludikhuize, M.C.; Meerlo, M.; Gallego, M.P.; Xanthakis, D.; Burgaya Julià, M.; Nguyen, N.T.B.; Brombacher, E.C.; Liv, N.; Maurice, M.M.; Paik, J.H.; et al. Mitochondria Define Intestinal Stem Cell Differentiation Downstream of a FOXO/Notch Axis. Cell Metab. 2020, 32, 889–900. [Google Scholar] [CrossRef]
- Li, X.J.; Jiang, Y.H.; Meisenhelder, J.L.; Yang, W.W.; Hawke, D.H.; Zheng, Y.H.; Xia, Y.; Aldape, K.; He, J.; Hunter, T.; et al. Mitochondria-Translocated PGK1 Functions as a Protein Kinase to Coordinate Glycolysis and the TCA Cycle in Tumorigenesis. Mol. Cell 2016, 61, 705–719. [Google Scholar] [CrossRef] [Green Version]
- Losón, O.C.; Song, Z.Y.; Chen, H.C.; Chan, D.C. Fis1, Mff, MiD49, and MiD51 mediate Drp1 recruitment in mitochondrial fission. Mol. Biol. Cell 2013, 24, 659–667. [Google Scholar] [CrossRef]
- Wu, W.X.; Li, W.; Chen, H.; Jiang, L.; Zhu, R.Z.; Feng, D. FUNDC1 is a novel mitochondrial-associated-membrane (MAM) protein required for hypoxia-induced mitochondrial fission and mitophagy. Autophagy 2016, 12, 1675–1676. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.H.; Qiu, B.Q.; Ma, M.; Zhang, R.; Hsu, S.J.; Liu, H.H.; Chen, J.; Gao, D.M.; Cui, J.F.; Ren, Z.G.; et al. Suppressing DRP1-mediated mitochondrial fission and mitophagy increases mitochondrial apoptosis of hepatocellular carcinoma cells in the setting of hypoxia. Oncogenesis 2020, 9, 67. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Liu, Y.F.; Tang, Y.; Wang, X.F.; Li, Z.C.; Li, R.; Ti, Z.Y.; Gao, W.D.; Bai, J.G.; Lv, Y. Increased mitochondrial fission is critical for hypoxia-induced pancreatic beta cell death. PLoS ONE 2018, 13, e0197266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina, A.J.A.; Wikstrom, J.D.; Stiles, L.; Las, G.; Mohamed, H.; Elorza, A.; Walzer, G.; Twig, G.; Katz, S.; Corkey, B.E.; et al. Mitochondrial networking protects beta-cells from nutrient-induced apoptosis. Diabetes 2009, 58, 2303–2315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Twig, G.; Hyde, B.; Shirihai, O.S. Mitochondrial fusion, fission and autophagy as a quality control axis: The bioenergetic view. Biochim. Biophys. Acta 2008, 1777, 1092–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.C.; Liu, J.; Kang, R.; Tang, D.L. Mitophagy Receptors in Tumor Biology. Front. Cell Dev. Biol. 2020, 8, 594203. [Google Scholar] [CrossRef]
- Little, A.C.; Kovalenko, I.; Goo, L.E.; Hong, H.S.; Kerk, S.A.; Yates, J.A.; Purohit, V.; Lombard, D.B.; Merajver, S.D.; Lyssiotis, C.A. High-content fluorescence imaging with the metabolic flux assay reveals insights into mitochondrial properties and functions. Commun. Biol. 2020, 3, 271. [Google Scholar] [CrossRef]
- Zhao, X.R.; Liu, L.D.; Li, R.; Wei, X.; Luan, W.Q.; Liu, P.S.; Zhao, J. Hypoxia-Inducible Factor 1-α (HIF-1α) Induces Apoptosis of Human Uterosacral Ligament Fibroblasts Through the Death Receptor and Mitochondrial Pathways. Med. Sci. Monit. 2018, 24, 8722–8733. [Google Scholar] [CrossRef]
- Chaban, Y.; Boekema, E.J.; Dudkina, N.V. Structures of mitochondrial oxidative phosphorylation supercomplexes and mechanisms for their stabilisation. Biochim. Biophys. Acta 2014, 1837, 418–426. [Google Scholar] [CrossRef] [Green Version]
- Fuhrmann, D.C.; Brüne, B. Mitochondrial composition and function under the control of hypoxia. Redox Biol. 2017, 12, 208–215. [Google Scholar] [CrossRef]
- Shimizu, M. NAD(+)/NADH homeostasis affects metabolic adaptation to hypoxia and secondary metabolite production in filamentous fungi. Biosci. Biotechnol. Biochem. 2018, 82, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.P.; Price, N.L.; Ling, A.J.; Moslehi, J.J.; Montgomery, M.K.; Rajman, L.; White, J.P.; Teodoro, J.S.; Wrann, C.D.; Hubbard, B.P.; et al. Declining NAD(+) induces a pseudohypoxic state disrupting nuclear-mitochondrial communication during aging. Cell 2013, 155, 1624–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, J.S.; D’Imprima, E.; Vonck, J. Mitochondrial Respiratory Chain Complexes. In Membrane Protein Complexes: Structure and Function; Springer: Singapore, 2018; Volume 87, pp. 167–227. [Google Scholar] [CrossRef]
- Mills, E.L.; Kelly, B.; Logan, A.; Costa, A.S.H.; Varma, M.; Bryant, C.E.; Tourlomousis, P.; Däbritz, J.H.M.; Gottlieb, E.; Latorre, I.; et al. Succinate Dehydrogenase Supports Metabolic Repurposing of Mitochondria to Drive Inflammatory Macrophages. Cell 2016, 167, 457–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierron, D.; Wildman, D.E.; Hüttemann, M.; Markondapatnaikuni, G.C.; Aras, S.; Grossman, L.I. Cytochrome c oxidase: Evolution of control via nuclear subunit addition. Biochim. Biophys. Acta 2012, 1817, 590–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | GenBank ID | Primer Sequence (5′→3′) | Product Size (bp) | Annealing Temperature/°C | Application |
|---|---|---|---|---|---|
| OPA1 | XM_012140446.1 | F: ATGAAATAGAACTCCGAATG R: GTCAACAAGCACCATCCT | 112 | 60 | qPCR |
| Mic60 | XM_012169573.1 | F: TTGAGATGGTCCTTGGTT R: TTGTTTCTGAGGTGGTGAG | 136 | 60 | |
| Drp1 | XM_015094867.2 | F: TCACCCGGAGACCTCTCATT R: TCCATGTAGCAGGGTCATTTTCT | 93 | 60 | |
| MFF | XM_027965256.1 | F: TCCAGCACGTGCATACTGAG R: CCGCCCCACTCACTAAATGT | 107 | 60 | |
| Fis1 | XM_027961118.1 | F: TGAAGTATGTGCGAGGGCTG R: CCATGCCCACTAGTCCATCTTT | 108 | 60 | |
| β-actin | NM_001009784.3 | F: GCTGTATTCCCCTCCATCGT R: GGATACCTCTCTTGCTCTGG | 97 | 60 | Reference gene |
| RPL19 | XM_012186026.3 | F: AATGCCAATGCCAACTC R: CCCTTTCGCTACCTATACC | 151 | 60 |
| Blood Physiological Indexes | Tibetan Sheep | ||
|---|---|---|---|
| TS25 | TS35 | TS45 | |
| Oxygen pressure, PO2 (mmHg) | 37.75 ± 2.02 a | 34.00 ± 1.78 a | 28.50 ± 1.04 b |
| Oxygen saturation, SO2 (%) | 72.00 ± 0.82 a | 65.75 ± 0.91 b | 59.75 ± 0.83 c |
| Hemoglobin, HGB (g/dL) | 11.60 ± 0.44 c | 13.05 ± 0.44 b | 15.80 ± 0.37 a |
| Hematocrit, HCT (%PCV) | 34.75 ± 0.85 c | 39.50 ± 0.87 b | 50.50 ± 0.87 a |
| Potential of hydrogen, PH | 7.37 ± 0.03 a | 7.33 ± 0.02 a | 7.31 ± 0.01 a |
| Carbon dioxide pressure, PCO2 (mmHg) | 55.70 ± 1.42 a | 40.75 ± 1.19 b | 34.92 ± 1.64 c |
| Concentration of bicarbonate, HCO3- (mmol/L) | 27.35 ± 0.79 a | 25.38 ± 0.83 a | 25.03 ± 0.81 a |
| Base excess, BE (mmol/L) | 3.00 ± 0.41 a | 3.25 ± 0.48 a | 3.75 ± 0.48 a |
| Blood Biochemical Indexes | Tibetan Sheep | ||
|---|---|---|---|
| TS25 | TS35 | TS45 | |
| Creatine Kinase, CK (U/L) | 256.60 ± 4.43 c | 300.34 ± 4.31 b | 499.62 ± 6.38 a |
| Creatine kinase isoenzymes, CK-MB (U/L) | 42.12 ± 1.86 c | 49.70 ± 1.90 b | 58.91 ± 2.12 a |
| Lactate dehydrogenase, LDH (U/L) | 616.57 ± 7.68 c | 833.31 ± 6.37 b | 906.08 ± 5.09 a |
| Lactate dehydrogenase isoenzymes, LDH1 (U/L) | 131.34 ± 4.25 b | 156.68 ± 4.48 a | 164.78 ± 5.23 a |
| Superoxide dismutase, SOD (U/mL) | 198.03 ± 4.77 b | 206.28 ± 4.59 b | 244.20 ± 4.65 a |
| Glutathione peroxidase, GPX (U/mL) | 54.76 ± 3.03 b | 64.83 ± 2.46a b | 69.06 ± 5.49 a |
| Low-density lipoprotein, LDL (mmol/L) | 0.52 ± 0.06 b | 0.72 ± 0.04 a | 0.81 ± 0.03 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, Y.; Li, S.; Zhao, F.; Wang, J.; Liu, X.; Hu, J.; Bao, G.; Luo, Y. Changes in the Mitochondrial Dynamics and Functions Together with the mRNA/miRNA Network in the Heart Tissue Contribute to Hypoxia Adaptation in Tibetan Sheep. Animals 2022, 12, 583. https://doi.org/10.3390/ani12050583
Wen Y, Li S, Zhao F, Wang J, Liu X, Hu J, Bao G, Luo Y. Changes in the Mitochondrial Dynamics and Functions Together with the mRNA/miRNA Network in the Heart Tissue Contribute to Hypoxia Adaptation in Tibetan Sheep. Animals. 2022; 12(5):583. https://doi.org/10.3390/ani12050583
Chicago/Turabian StyleWen, Yuliang, Shaobin Li, Fangfang Zhao, Jiqing Wang, Xiu Liu, Jiang Hu, Gaoliang Bao, and Yuzhu Luo. 2022. "Changes in the Mitochondrial Dynamics and Functions Together with the mRNA/miRNA Network in the Heart Tissue Contribute to Hypoxia Adaptation in Tibetan Sheep" Animals 12, no. 5: 583. https://doi.org/10.3390/ani12050583