
Melatonin Protects Bovine Spermatozoa by Reinforcing Their Antioxidant Defenses
 ,
,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Semen Collection, Sample Preparation and Experimental Design
2.3. Motility Assessment
2.4. Viability Assessment
2.5. Preparation of Whole Cell Lysates
2.6. Determination of Intracellular Glutathione (GSH)
2.7. Determination of the Intracellular Superoxide Anion Content
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
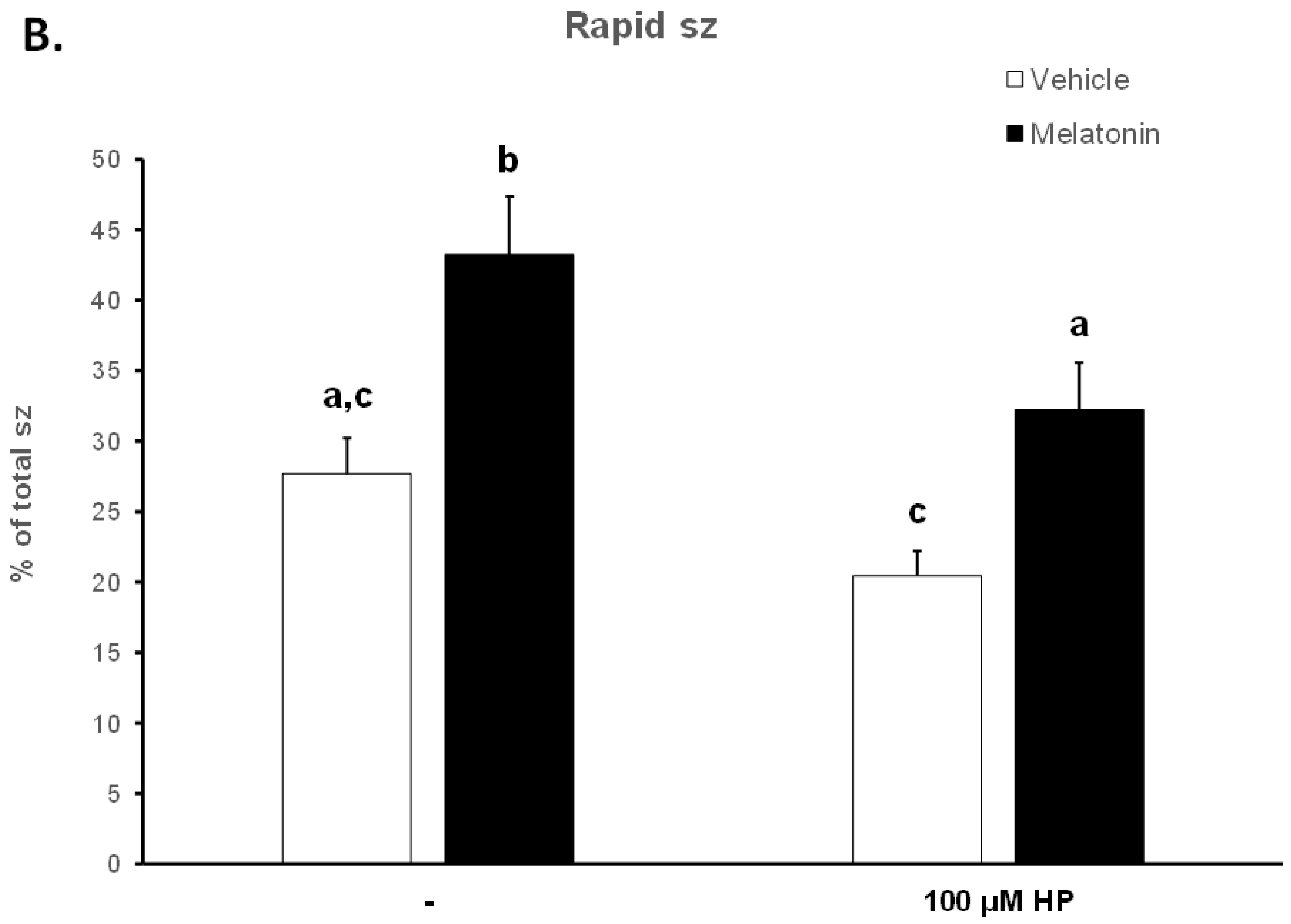
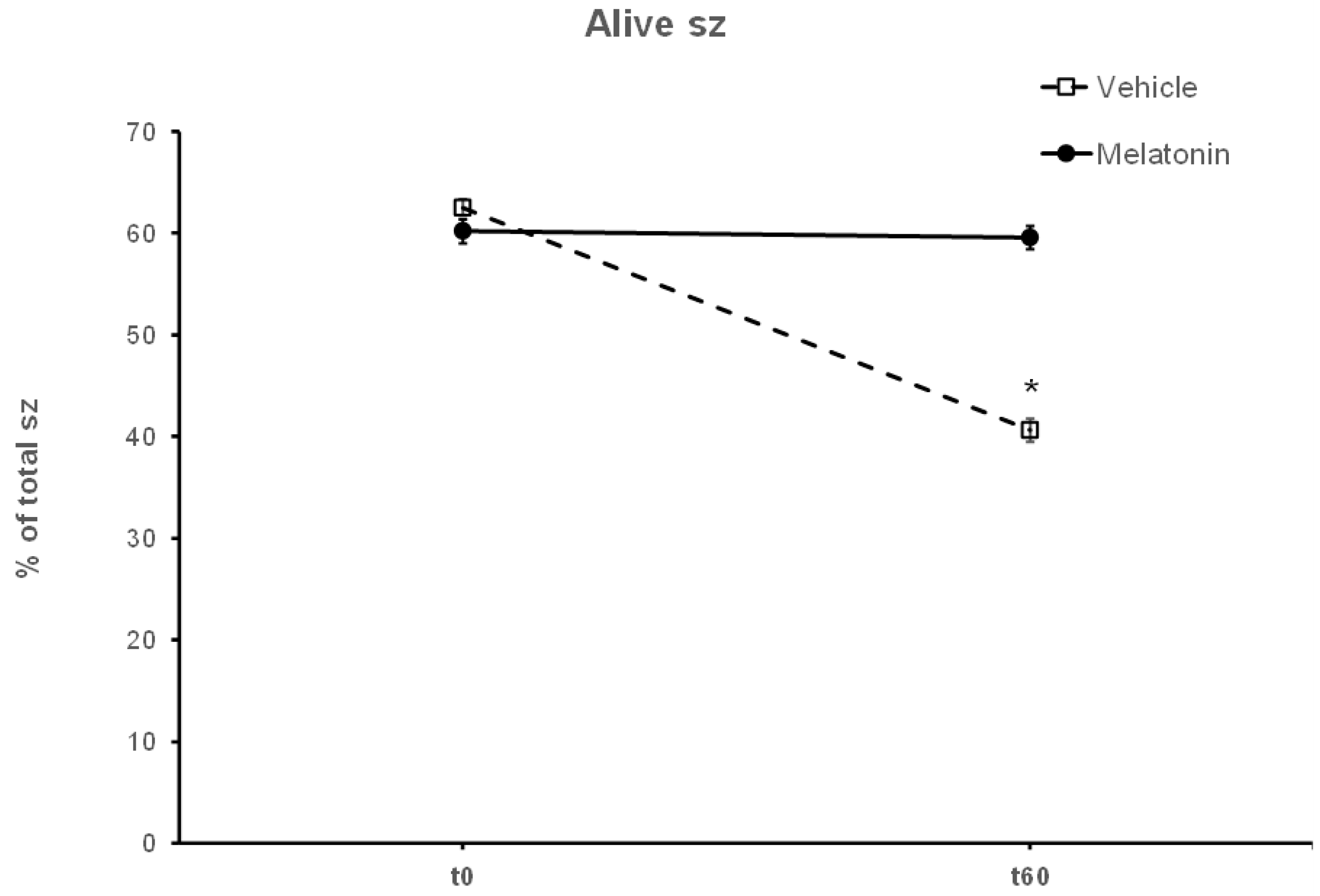
3.1. Melatonin Inhibits the Loss of Motility over Time during the Handling of Spermatozoa
3.2. In the Presence of Melatonin, Spermatozoa Retain Their Motility during Insult by Hydrogen Peroxide
3.3. Melatonin Protects Viability of Spermatozoa against Oxidative Stress
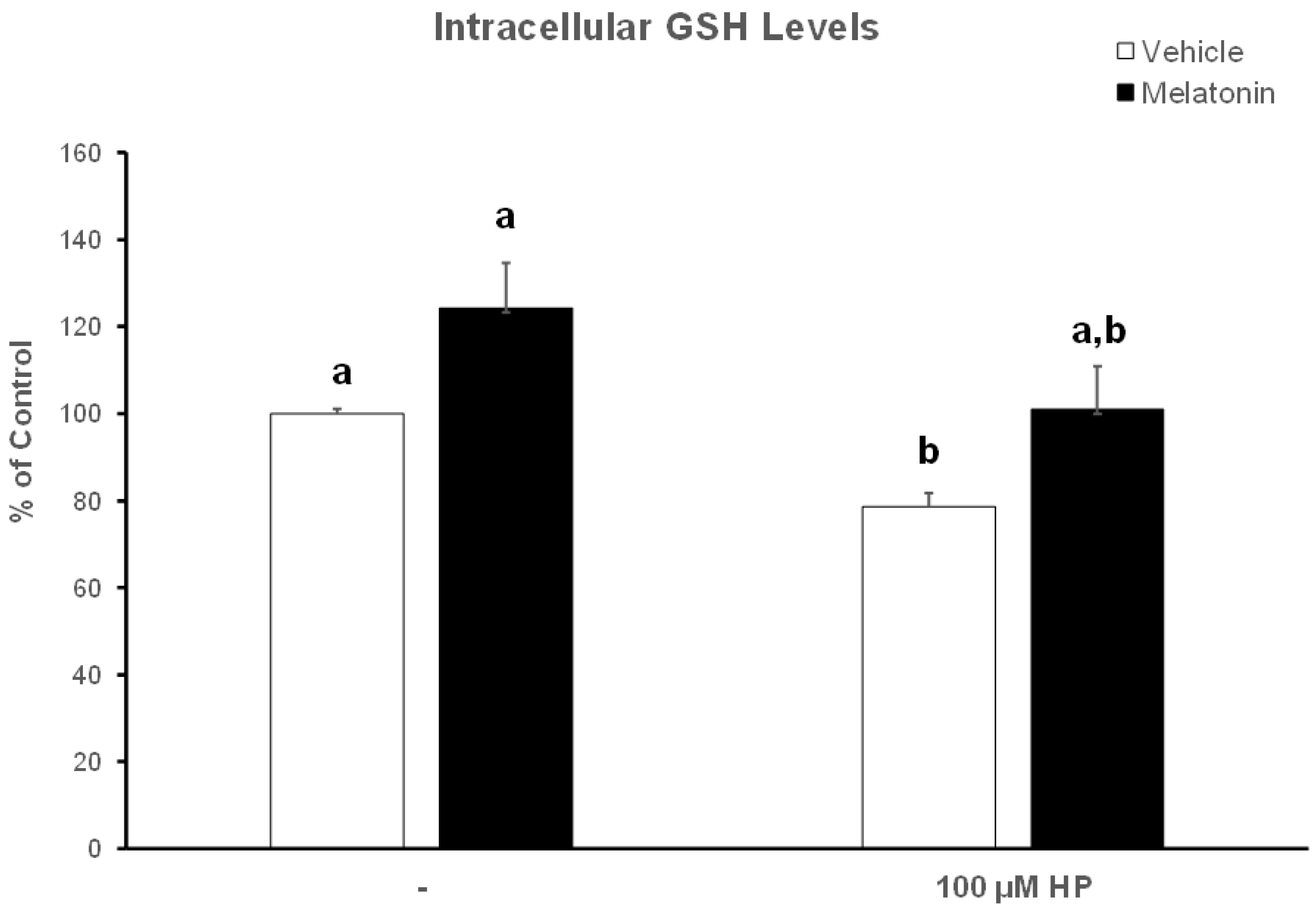
3.4. In the Presence of Melatonin, Spermatozoa Retain Their GSH Content
3.5. Melatonin Does Not Affect the Intracellular Levels of Superoxide Ion
3.6. Melatonin Enables Spermatozoa to Retain Low Levels of the Inducible Nitric Oxide Synthase (iNOS)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martinez, M.; Majzoub, A. Best Laboratory Practices and Therapeutic Interventions to Reduce Sperm DNA Damage. Andrologia 2021, 53, e13736. [Google Scholar] [CrossRef]
- Ugur, M.R.; Saber Abdelrahman, A.; Evans, H.C.; Gilmore, A.A.; Hitit, M.; Arifiantini, R.I.; Purwantara, B.; Kaya, A.; Memili, E. Advances in Cryopreservation of Bull Sperm. Front. Vet. Sci. 2019, 6, 268. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.W.; Zhang, H.; Ikemoto, I.; Anderson, D.J.; Loughlin, K.R. Reactive Oxygen Species Generation by Seminal Cells during Cryopreservation. Urology 1997, 49, 921–925. [Google Scholar] [CrossRef]
- Said, T.M.; Gaglani, A.; Agarwal, A. Implication of Apoptosis in Sperm Cryoinjury. Reprod. Biomed. Online 2010, 21, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Kogan, T.; Dahan, D.G.; Laor, R.; Argov-Argaman, N.; Zeron, Y.; Komsky-Elbaz, A.; Kalo, D.; Roth, Z. Association between Fatty Acid Composition, Cryotolerance and Fertility Competence of Progressively Motile Bovine Spermatozoa. Animals 2021, 11, 2948. [Google Scholar] [CrossRef] [PubMed]
- Lenzi, A.; Picardo, M.; Gandini, L.; Dondero, F. Lipids of the Sperm Plasma Membrane: From Polyunsaturated Fatty Acids Considered as Markers of Sperm Function to Possible Scavenger Therapy. Hum. Reprod. Update 1996, 2, 246–256. [Google Scholar] [CrossRef]
- Baldi, E.; Tamburrino, L.; Muratori, M.; Degl’Innocenti, S.; Marchiani, S. Adverse Effects of in vitro Manipulation of Spermatozoa. Anim. Reprod. Sci. 2020, 220, 106314. [Google Scholar] [CrossRef]
- Chaudière, J. Chapter 2 – Some Chemical and Biochemical Constraints of Oxidative Stress in Living Cells*. New Comprehensive Biochemistry 1994, 28, 25–66. [Google Scholar]
- Tsantarliotou, M.P.; Sapanidou, V.G. The Importance of Antioxidants in Sperm Quality and in vitro Embryo Production. J. Vet. Androl. 2018, 3, 1–12. [Google Scholar]
- Sapanidou, V.; Tsantarliotou, M.P.; Lavrentiadou, S.N. A Review of the Use of Antioxidants in Bovine Sperm Preparation Protocols. Anim. Reprod. Sci. 2023, 251, 107215. [Google Scholar] [CrossRef]
- Sapanidou, V.; Lavrentiadou, S.N.; Errico, M.; Panagiotidis, I.; Fletouris, D.; Efraimidis, I.; Zervos, I.; Taitzoglou, I.; Gasparrini, B.; Tsantarliotou, M. The Addition of Crocin in the Freezing Medium Extender Improves Post-Thaw Semen Quality. Reprod. Domest. Anim. 2022, 57, 269–276. [Google Scholar] [CrossRef]
- Pintus, E.; Ros-Santaella, J.L. Impact of Oxidative Stress on Male Reproduction in Domestic and Wild Animals. Antioxidants 2021, 10, 1154. [Google Scholar] [CrossRef]
- Sapanidou, V.; Taitzoglou, I.; Tsakmakidis, I.; Kourtzelis, I.; Fletouris, D.; Theodoridis, A.; Lavrentiadou, S.; Tsantarliotou, M. Protective Effect of Crocetin on Bovine Spermatozoa against Oxidative Stress during in Vitro Fertilization. Andrology 2016, 4, 1138–1149. [Google Scholar] [CrossRef]
- Bathgate, R. Antioxidant Mechanisms and Their Benefit on Post-Thaw Boar Sperm Quality. Reprod. Domest. Anim. 2011, 46, 23–25. [Google Scholar] [CrossRef]
- Karunakaran, M.; Devanathan, T.; Kulasekar, K.; Sridevi, P.; Jawahar, T.P.; Loganatahsamy, K.; Dhali, A.; Sellappan, S. Effect of Heparin Binding Protein and Hydrogen Peroxide on Lipid Peroxidation Status of Bovine Sperm Cells. Indian J. Anim. Sci. 2012, 82, 976–978. [Google Scholar]
- Ari Pujianto, D.; Oktarina, M.; Ayu Sharma Sharaswati, I. Hydrogen Peroxide Has Adverse Effects on Human Sperm Quality Parameters, Induces Apoptosis, and Reduces Survival. J. Hum. Reprod. Sci. 2021, 14, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Hacışevki, A.; Baba, B. An Overview of Melatonin as an Antioxidant Molecule: A Biochemical Approach. In Melatonin—Molecular Biology, Clinical and Pharmaceutical Approaches; Drăgoi, C.M., Nicolae, A.C., Eds.; IntechOpen: London, UK, 2018; pp. 59–85. ISBN 978-1-78984-504-4. [Google Scholar]
- Tsantarliotou, M.P.; Kokolis, N.A.; Smokovitis, A. Melatonin Administration Increased Plasminogen Activator Activity in Ram Spermatozoa. Theriogenology 2008, 69, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Lialiaris, T.S. Melatonin in Prevention of Mutagenesis, Oxidation and Other Damage to Cells. In Proceedings of the Melatonin in the Promotion of Health; CRC Press: Boca Raton, FL, USA, 2012; pp. 81–88. [Google Scholar]
- Casao, A.; Cebrián, I.; Asumpção, M.E.; Pérez-Pé, R.; Abecia, J.A.; Forcada, F.; Cebrián-Pérez, J.A.; Muiño-Blanco, T. Seasonal Variations of Melatonin in Ram Seminal Plasma Are Correlated to Those of Testosterone and Antioxidant Enzymes. Reprod. Biol. Endocrinol. 2010, 8, 59. [Google Scholar] [CrossRef]
- Monllor, F.; Espino, J.; Marchena, A.M.; Ortiz, Á.; Lozano, G.; García, J.F.; Pariente, J.A.; Rodríguez, A.B.; Bejarano, I. Melatonin Diminishes Oxidative Damage in Sperm Cells, Improving Assisted Reproductive Techniques. Turk. J. Biol. 2017, 41, 881–889. [Google Scholar] [CrossRef]
- Espino, J.; Ortiz, Á.; Bejarano, I.; Lozano, G.M.; Monllor, F.; García, J.F.; Rodríguez, A.B.; Pariente, J.A. Melatonin Protects Human Spermatozoa from Apoptosis via Melatonin Receptor- and Extracellular Signal-Regulated Kinase-Mediated Pathways. Fertil. Steril. 2011, 95, 2290–2296. [Google Scholar] [CrossRef]
- Domínguez-Rebolledo, A.E.; Fernández-Santos, M.R.; Bisbal, A.; Ros-Santaella, J.L.; Ramón, M.; Carmona, M.; Martínez-Pastor, F.; Garde, J.J. Improving the Effect of Incubation and Oxidative Stress on Thawed Spermatozoa from Red Deer by Using Different Antioxidant Treatments. Reprod. Fertil. Dev. 2010, 22, 856–870. [Google Scholar] [CrossRef] [PubMed]
- Casao, A.; Abecia, J.A.; Cebrián-Pérez, J.A.; Muiño-Blanco, T.; Vázquez, M.I.; Forcada, F. The Effects of Melatonin on in vitro Oocyte Competence and Embryo Development in Sheep|Efecto de La Melatonina En La Competencia Del Oocito y El Desarrollo Embrionario Ovino in vitro. Span. J. Agric. Res. 2010, 8, 35–41. [Google Scholar] [CrossRef]
- Jang, H.Y.; Kim, Y.H.; Kim, B.W.; Park, I.C.; Cheong, H.T.; Kim, J.T.; Park, C.K.; Kong, H.S.; Lee, H.K.; Yang, B.K. Ameliorative Effects of Melatonin against Hydrogen Peroxide-Induced Oxidative Stress on Boar Sperm Characteristics and Subsequent in vitro Embryo Development. Reprod. Domest. Anim. 2010, 45, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-Y.; Hao, H.-S.; Zhao, Y.-H.; Zhang, P.-P.; Wang, H.-Y.; Pang, Y.-W.; Du, W.-H.; Zhao, S.-J.; Liu, Y.; Huang, J.-M.; et al. Melatonin Improves the Fertilization Capacity of Sex-Sorted Bull Sperm by Inhibiting Apoptosis and Increasing Fertilization Capacitation via MT1. Int. J. Mol. Sci. 2019, 20, 3921. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.-W.; Sun, Y.-Q.; Jiang, X.-L.; Huang, Z.-Q.; Zhao, S.-J.; Du, W.-H.; Hao, H.-S.; Zhao, X.-M.; Zhu, H.-B. Protective Effects of Melatonin on Bovine Sperm Characteristics and Subsequent in vitro Embryo Development. Mol. Reprod. Dev. 2016, 83, 993–1002. [Google Scholar] [CrossRef]
- Carlos Gutié Rrez-Añez Id, J.; Henning, H.; Lucas-Hahn, A.; Baulain, U.; Aldag, P.; Sieg, B.; Hensel, V.; Herrmann, D.; Niemann, H. Melatonin Improves Rate of Monospermic Fertilization and Early Embryo Development in a Bovine IVF System. PLoS ONE 2021, 16, e0256701. [Google Scholar] [CrossRef]
- Fernández-Alegre, E.; Álvarez-Fernández, I.; Domínguez, J.C.; Casao, A.; Martínez-Pastor, F. Melatonin Non-Linearly Modulates Bull Spermatozoa Motility and Physiology in Capacitating and Non-Capacitating Conditions. Int. J. Mol. Sci. 2020, 21, 2701. [Google Scholar] [CrossRef]
- Casao, A.; Gallego, M.; Abecia, J.A.; Forcada, F.; Pérez-Pé, R.; Muiño-Blanco, T.; Cebrián-Pérez, J.Á. Identification and Immunolocalisation of Melatonin MT(1) and MT(2) Receptors in Rasa Aragonesa Ram Spermatozoa. Reprod. Fertil. Dev. 2012, 24, 953–961. [Google Scholar] [CrossRef]
- Gonzalez-Arto, M.; Luna, C.; Pérez-Pé, R.; Muiño-Blanco, T.; Cebrián-Pérez, J.A.; Casao, A. New Evidence of Melatonin Receptor Contribution to Ram Sperm Functionality. Reprod. Fertil. Dev. 2016, 28, 924–935. [Google Scholar] [CrossRef]
- Succu, S.; Berlinguer, F.; Pasciu, V.; Satta, V.; Leoni, G.G.; Naitana, S. Melatonin Protects Ram Spermatozoa from Cryopreservation Injuries in a Dose-Dependent Manner. J. Pineal Res. 2011, 50, 310–318. [Google Scholar] [CrossRef]
- Fujinoki, M. Melatonin-Enhanced Hyperactivation of Hamster Sperm. Reproduction 2008, 136, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Gwayi, N.; Bernard, R.T.F. The Effects of Melatonin on Sperm Motility in vitro in Wistar Rats. Andrologia 2002, 34, 391–396. [Google Scholar] [CrossRef]
- Ashrafi, I.; Kohram, H.; Ardabili, F.F. Antioxidative Effects of Melatonin on Kinetics, Microscopic and Oxidative Parameters of Cryopreserved Bull Spermatozoa. Anim. Reprod. Sci. 2013, 139, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Cheuquemán, C.; Arias, M.E.; Risopatrón, J.; Felmer, R.; Álvarez, J.; Mogas, T.; Sánchez, R. Supplementation of IVF Medium with Melatonin: Effect on Sperm Functionality and in vitro Produced Bovine Embryos. Andrologia 2015, 47, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Gimeno-Martos, S.; Casao, A.; Yeste, M.; Cebrián-Pérez, J.A.; Muiño-Blanco, T.; Pérez-Pé, R. Melatonin Reduces CAMP-Stimulated Capacitation of Ram Spermatozoa. Reprod. Fertil. Dev. 2019, 31, 420–431. [Google Scholar] [CrossRef]
- Tomás-Zapico, C.; Coto-Montes, A. Melatonin as Antioxidant under Pathological Processes. Recent Pat. Endocr. Metab. Immune Drug Discov. 2007, 1, 63–82. [Google Scholar] [CrossRef]
- Gilad, E.; Wong, H.R.; Zingarelli, B.; Virág, L.; O’connor, M.; Salzman, A.L.; Szabó, C. Melatonin Inhibits Expression of the Inducible Isoform of Nitric Oxide Synthase in Murine Macrophages: Role of Inhibition of NFκB Activation. FASEB J. 1998, 12, 685–693. [Google Scholar] [CrossRef]
- Luo, Y.; Zhu, Y.; Basang, W.; Wang, X.; Li, C.; Zhou, X. Roles of Nitric Oxide in the Regulation of Reproduction: A Review. Front. Endocrinol. 2021, 12, 752410. [Google Scholar] [CrossRef]
- Catt, J.W.; Henman, M. Toxic Effects of Oxygen on Human Embryo Development. Hum. Reprod. 2000, 15 (Suppl. S2), 199–206. [Google Scholar] [CrossRef]
- Gadea, J.; García-Vazquez, F.; Matás, C.; Gardón, J.C.; Cánovas, S.; Gumbao, D. Cooling and Freezing of Boar Spermatozoa: Supplementation of the Freezing Media with Reduced Glutathione Preserves Sperm Function. J. Androl. 2005, 26, 396–404. [Google Scholar] [CrossRef]
- Gadea, J.; Gumbao, D.; Cánovas, S.; García-Vázquez, F.A.; Alberto Grulló, L.; Carlos Gardó, J. Supplementation of the Dilution Medium after Thawing with Reduced Glutathione Improves Function and the in vitro Fertilizing Ability of Frozen-Thawed Bull Spermatozoa. Int. J. Androl. 2008, 31, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Sapanidou, V.; Taitzoglou, I.; Tsakmakidis, I.; Kourtzelis, I.; Fletouris, D.; Theodoridis, A.; Zervos, I.; Tsantarliotou, M. Antioxidant Effect of Crocin on Bovine Sperm Quality and in vitro Fertilization. Theriogenology 2015, 84, 1273–1282. [Google Scholar] [CrossRef] [PubMed]
- Vajta, G.; Holm, P.; Greve, T.; Callesen, H. Factors Affecting Survival Rates of in vitro Produced Bovine Embryos after Vitrification and Direct In-Straw Rehydration. Anim. Reprod. Sci. 1996, 45, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Mayer, D.T.; Squiers, C.D.; Bogart, R.; Oloufa, M.M. The Technique for Characterizing Mammalian Spermatozoa as Dead or Living by Differential Staining. J. Anim. Sci. 1951, 10, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, A.; Petrotos, K.; Stagos, D.; Gerasopoulos, K.; Maimaris, A.; Makris, H.; Kafantaris, I.; Makri, S.; Kerasioti, E.; Halabalaki, M.; et al. Enhancement of Antioxidant Mechanisms and Reduction of Oxidative Stress in Chickens after the Administration of Drinking Water Enriched with Polyphenolic Powder from Olive Mill Waste Waters. Oxid. Med. Cell. Longev. 2017, 2017, 8273160. [Google Scholar] [CrossRef]
- Becerra, M.C.; Eraso, A.J.; Albesa, I. Comparison of Oxidative Stress Induced by Ciprofloxacin and Pyoverdin in Bacteria and in Leukocytes to Evaluate Toxicity. Luminescence 2003, 18, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Halevas, E.; Matsia, S.; Hatzidimitriou, A.; Geromichalou, E.; Papadopoulos, T.A.; Katsipis, G.; Pantazaki, A.; Litsardakis, G.; Salifoglou, A. A Unique Ternary Ce(III)-Quercetin-Phenanthroline Assembly with Antioxidant and Anti-Inflammatory Properties. J. Inorg. Biochem. 2022, 235, 111947. [Google Scholar] [CrossRef] [PubMed]
- Katsipis, G.; Tzekaki, E.E.; Tsolaki, M.; Pantazaki, A.A. Salivary GFAP as a Potential Biomarker for Diagnosis of Mild Cognitive Impairment and Alzheimer’s Disease and Its Correlation with Neuroinflammation and Apoptosis. J. Neuroimmunol. 2021, 361, 577744. [Google Scholar] [CrossRef]
- Gualtieri, R.; Kalthur, G.; Barbato, V.; Longobardi, S.; Di Rella, F.; Adiga, S.K.; Talevi, R.; Moretti, E. Sperm Oxidative Stress during In Vitro Manipulation and Its Effects on Sperm Function and Embryo Development. Antioxidants 2021, 10, 1025. [Google Scholar] [CrossRef]
- Toor, J.S.; Sikka, S.C. Chapter 1.6—Human Spermatozoa and Interactions With Oxidative Stress. In Oxidants, Antioxidants and Impact of the Oxidative Status in Male Reproduction; Henkel, R., Samanta, L., Agarwal, A., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 45–53. ISBN 978-0-12-812501-4. [Google Scholar]
- O’Flaherty, C.; de Lamirande, E.; Gagnon, C. Positive Role of Reactive Oxygen Species in Mammalian Sperm Capacitation: Triggering and Modulation of Phosphorylation Events. Free. Radic. Biol. Med. 2006, 41, 528–540. [Google Scholar] [CrossRef]
- de Lamirande, E.; O’Flaherty, C. Sperm Activation: Role of Reactive Oxygen Species and Kinases. Biochim. Biophys. Acta Proteins Proteom. 2008, 1784, 106–115. [Google Scholar] [CrossRef]
- O’Flaherty, C. Redox Regulation of Mammalian Sperm Capacitation. Asian J. Androl. 2015, 17, 583–590. [Google Scholar] [CrossRef]
- Agarwal, A.; Said, T.M.; Bedaiwy, M.A.; Banerjee, J.; Alvarez, J.G. Oxidative Stress in an Assisted Reproductive Techniques Setting. Fertil. Steril. 2006, 86, 503–512. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, C.; Scarlata, E. Oxidative Stress and Reproductive Function: The Protection of Mammalian Spermatozoa against Oxidative Stress. Reproduction 2022, 164, F67–F78. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Maldonado Rosas, I.; Anagnostopoulou, C.; Cannarella, R.; Boitrelle, F.; Munoz, L.V.; Finelli, R.; Durairajanayagam, D.; Henkel, R.; Saleh, R. Oxidative Stress and Assisted Reproduction: A Comprehensive Review of Its Pathophysiological Role and Strategies for Optimizing Embryo Culture Environment. Antioxidants 2022, 11, 477. [Google Scholar] [CrossRef] [PubMed]
- Lonergan, P.; Fair, T. Maturation of Oocytes in vitro. Annu. Rev. Anim. Biosci. 2016, 4, 255–268. [Google Scholar] [CrossRef]
- Rizos, D.; Clemente, M.; Bermejo-Alvarez, P.; De La Fuente, J.; Lonergan, P.; Gutiérrez-Adán, A. Consequences of in vitro Culture Conditions on Embryo Development and Quality. Reprod. Domest. Anim. 2008, 43, 44–50. [Google Scholar] [CrossRef]
- Martins da Silva, S.J. DNA Fragmentation, Antioxidants and ART. In Assisted Reproduction Techniques; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2021; pp. 606–611. ISBN 9781119622215. [Google Scholar]
- Agarwal, A.; Durairajanayagam, D.; Du Plessis, S.S. Utility of Antioxidants during Assisted Reproductive Techniques: An Evidence Based Review. Reprod. Biol. Endocrinol. 2014, 12, 112. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Zeng, Z.; Chen, X.; Fan, B.; Meng, Z. Melatonin Supplementation to the Freezing Medium Enhances Post-Thaw Sperm Quality and Fertility of Giant Grouper (Epinephelus lanceolatus). Aquac Rep 2023, 31, 101662. [Google Scholar] [CrossRef]
- Brahmajosyula, M.; Morimoto, Y. Exogenous GSH Supplementation to Raw Semen Alters Sperm Kinematic Parameters in Infertile Patients. Reprod. Sci. 2023, 30, 2853–2865. [Google Scholar] [CrossRef]
- Nguyen, N.D.; Le, M.T.; Tran, N.Q.T.; Nguyen, Q.H.V.; Cao, T.N. Micronutrient Supplements as Antioxidants in Improving Sperm Quality and Reducing DNA Fragmentation. Basic Clin. Androl. 2023, 33, 23. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.-X.; Cabrera, J.; D’Arpa, D.; Sainz, R.M.; Mayo, J.; Ramos, S. The Oxidant/Antioxidant Network: Role of Melatonin. Biol. Signals Recept. 1999, 8, 56–63. [Google Scholar] [CrossRef]
- Ahmad, S.B.; Ali, A.; Bilal, M.; Rashid, S.M.; Wani, A.B.; Bhat, R.R.; Rehman, M.U. Melatonin and Health: Insights of Melatonin Action, Biological Functions, and Associated Disorders. Mol. Neurobiol. 2023, 43, 2437–2458. [Google Scholar] [CrossRef]
- Feng, X.; Zhang, Y.; Li, N.; Zhang, Y.; Zheng, Q.; Sun, M.; Tang, J.; Xu, Z. Melatonin in Reproductive Medicine: A Promising Therapeutic Target? Curr. Med. Chem. 2023, 30, 3090–3118. [Google Scholar] [CrossRef]
- Dehdari Ebrahimi, N.; Sadeghi, A.; Ala, M.; Ebrahimi, F.; Pakbaz, S.; Azarpira, N. Protective Effects of Melatonin against Oxidative Stress Induced by Metabolic Disorders in the Male Reproductive System: A Systematic Review and Meta-Analysis of Rodent Models. Front. Endocrinol. 2023, 14, 1202560. [Google Scholar] [CrossRef] [PubMed]
- Dehdari Ebrahimi, N.; Shojaei-Zarghani, S.; Taherifard, E.; Dastghaib, S.; Parsa, S.; Mohammadi, N.; Sabet Sarvestani, F.; Moayedfard, Z.; Hosseini, N.; Safarpour, H.; et al. Protective Effects of Melatonin against Physical Injuries to Testicular Tissue: A Systematic Review and Meta-Analysis of Animal Models. Front. Endocrinol. 2023, 14, 1123999. [Google Scholar] [CrossRef]
- Silva, B.R.; Silva, J.R.V. Mechanisms of Action of Non-Enzymatic Antioxidants to Control Oxidative Stress during in vitro Follicle Growth, Oocyte Maturation, and Embryo Development. Anim. Reprod. Sci. 2023, 249, 107186. [Google Scholar] [CrossRef] [PubMed]
- Role, W.; Silva, A.M.S.; Kim, Y.-B.; Li, Z.; Zhang, K.; Zhou, Y.; Zhao, J.; Wang, J.; Lu, W. Citation: Molecules Role of Melatonin in Bovine Reproductive Biotechnology. Molecules 2023, 28, 4940. [Google Scholar] [CrossRef]
- Griveau, J.E.; Renard, P.; Lannou, D.L. An in vitro Promoting Role for Hydrogen Peroxide in Human Sperm Capacitation. Int. J. Androl. 1994, 17, 300–307. [Google Scholar] [CrossRef]
- Rivlin, J.; Mendel, J.; Rubinstein, S.; Etkovitz, N.; Breitbart, H. Role of Hydrogen Peroxide in Sperm Capacitation and Acrosome Reaction1. Biol. Reprod. 2004, 70, 518–522. [Google Scholar] [CrossRef]
- ChaithraShree, A.R.; Ingole, S.D.; Dighe, V.D.; Nagvekar, A.S.; Bharucha, S.V.; Dagli, N.R.; Kekan, P.M.; Kharde, S.D. Effect of Melatonin on Bovine Sperm Characteristics and Ultrastructure Changes Following Cryopreservation. Veter. Med. Sci. 2020, 6, 177–186. [Google Scholar] [CrossRef]
- Li, C.; Zhou, X. Melatonin and Male Reproduction. Clin. Chim. Acta 2015, 446, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, A.; Espino, J.; Bejarano, I.; Lozano, G.M.; Monllor, F.; García, J.F.; Pariente, J.A.; Rodríguez, A.B. High Endogenous Melatonin Concentrations Enhance Sperm Quality and Short-Term in vitro Exposure to Melatonin Improves Aspects of Sperm Motility. J. Pineal Res. 2010, 50, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Cebrián-Pérez, J.A.; Casao, A.; González-Arto, M.; dos Santos Hamilton, T.R.; Pérez-Pé, R.; Muiño-Blanco, T. Melatonin in Sperm Biology: Breaking Paradigms. Reprod. Domest. Anim. 2014, 49, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Szabó, C.; Ischiropoulos, H.; Radi, R. Peroxynitrite: Biochemistry, Pathophysiology and Development of Therapeutics. Nat. Rev. Drug Discov. 2007, 6, 662–680. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, A.; Reiter, R.J.; Topal, T.; Manchester, L.C.; Oter, S.; Tan, D.-X. Melatonin: An Established Antioxidant Worthy of Use in Clinical Trials. Mol. Med. 2009, 15, 43–50. [Google Scholar] [CrossRef]
- Du Plessis, S.S.; Hagenaar, K.; Lampiao, F. The in vitro Effects of Melatonin on Human Sperm Function and Its Scavenging Activities on NO and ROS. Andrologia 2010, 42, 112–116. [Google Scholar] [CrossRef]
- Alderton, W.K.; Cooper, C.E.; Knowles, R.G. Nitric Oxide Synthases: Structure, Function and Inhibition. Biochem. J. 2001, 357, 593–615. [Google Scholar] [CrossRef]
- Pandi-Perumal, S.R.; Trakht, I.; Srinivasan, V.; Spence, D.W.; Maestroni, G.J.M.; Zisapel, N.; Cardinali, D.P. Physiological Effects of Melatonin: Role of Melatonin Receptors and Signal Transduction Pathways. Prog. Neurobiol. 2008, 85, 335–353. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | PM | VCL | VSL | VAP | ALH |
|---|---|---|---|---|---|
| Control-t0 | 21.16 ± 1.74 | 60.89 ± 3.17 a | 19.43 ± 1.19 a,b | 34.52 ± 1.87 a,b | 3.26 ± 0.16 a |
| Melatonin-t0 | 23.77 ± 1.55 | 59.58 ± 1.22 a | 20.93 ± 0.5 a,b | 35.83 ± 0.46 a,b | 3.09 ± 0.08 a |
| Control-t60 | 23.58 ± 2.23 | 49.82 ± 1.68 b | 18.55 ± 0.96 a,b | 30.47 ± 1.09 a,c | 2.72 ± 0.2 b,c |
| HP | 20.25 ± 4.62 | 43.72 ± 5.6 b | 17.83 ± 1.89 a | 23.85 ± 4.17 c | 2.93 ± 0.05 c |
| Melatonin-t60 | 24.3 ± 4.38 | 62.15 ± 1.84 a | 21.55 ± 1.95 b | 36.67 ± 1.49 b | 3.38 ± 0.18 a |
| Melatonin-HP | 19.12 ± 2.81 | 54.38 ± 4.31 a,b | 18.4 ± 1.08 a,b | 32 ± 1.93 a,b | 3.18 ± 0.19 a,b,c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lavrentiadou, S.N.; Sapanidou, V.; Tzekaki, E.E.; Margaritis, I.; Tsantarliotou, M.P. Melatonin Protects Bovine Spermatozoa by Reinforcing Their Antioxidant Defenses. Animals 2023, 13, 3219. https://doi.org/10.3390/ani13203219
Lavrentiadou SN, Sapanidou V, Tzekaki EE, Margaritis I, Tsantarliotou MP. Melatonin Protects Bovine Spermatozoa by Reinforcing Their Antioxidant Defenses. Animals. 2023; 13(20):3219. https://doi.org/10.3390/ani13203219
Chicago/Turabian StyleLavrentiadou, Sophia N., Vasiliki Sapanidou, Elena E. Tzekaki, Ioannis Margaritis, and Maria P. Tsantarliotou. 2023. "Melatonin Protects Bovine Spermatozoa by Reinforcing Their Antioxidant Defenses" Animals 13, no. 20: 3219. https://doi.org/10.3390/ani13203219
APA StyleLavrentiadou, S. N., Sapanidou, V., Tzekaki, E. E., Margaritis, I., & Tsantarliotou, M. P. (2023). Melatonin Protects Bovine Spermatozoa by Reinforcing Their Antioxidant Defenses. Animals, 13(20), 3219. https://doi.org/10.3390/ani13203219