Simple Summary
The emergence of antibiotics resistance is a warning sign to limit antibiotics usage as growth promoters in the poultry industry. Probiotics served as superior candidates for replacing antibiotics in the poultry sector. However, the beneficial functions of probiotics did not reach their targeted outcomes owing to the harsh environment in the poultry gut. In this context, evolution of biotechnological aids offers new avenues for increasing bioavailability and beneficial efficacy of in-feed additives including probiotics. Therefore, encapsulation of Bacillus amyloliquefaciens (B. amyloliquefaciens) into nanocarriers boosted its growth-promoting purposes and consequently modulated the functions of digestive enzymes and kept the microbiota homeostasis towards the beneficial ones. The strengthening capability of B. amyloliquefaciens-loaded nanoparticles for broilers’ gut barrier limited Campylobacter jejuni (C. jejuni) colonization and shedding. This superior outcome would in turn interrupt the transmission cycle of C. jejuni through the food chain and consequently protect against its adverse consequences in humans.
Abstract
Probiotics as novel antibiotics’ substitutes are verified to provide barriers for hindering the colonization of enteric bacterial pathogens with nutritional benefits. For enhancement of the probiotics’ effectiveness, their integration within nanomaterials is a paramount tool to support the progress of new compounds with functional features. Therefore, we addressed the impact of effective delivery of probiotics (Bacillus amyloliquefaciens) loaded nanoparticles (BNPs) on performance and Campylobacter jejuni (C. jejuni) shedding and colonization in poultry. Two hundred Ross broiler chickens were divided into four groups fed various BNP levels: BNPs I, BNPs II, BNPs III, and BNPs-free diets for 35 days. Nanoparticles delivery of probiotics within broiler diets improved growth performance as reflected by higher body weight gain and superior feed conversion ratio, especially in BNPs II- and BNPs III-fed groups. In parallel, the mRNA expression levels of digestive enzymes encoding genes (AMY2a, PNLIP, CELA1, and CCK) achieved their peaks in BNPs III-fed group (1.69, 1.49, 1.33, and 1.29-fold change, respectively) versus the control one. Notably, with increasing the levels of BNPs, the abundance of beneficial microbiota, such as Bifidobacterium and Lactobacillus species, was favored over harmful ones, including Clostridium species and Enterobacteriaceae. Birds fed higher levels of BNPs displayed significant improvement in the expression of barrier functions-linked genes including DEFB1, FABP-2, and MUC-2 alongside substantial reduction in cecal colonization and fecal shedding of C. jejuni. From the aforementioned positive effects of BNPs, we concluded their potential roles as growth promoters and effective preventive aids for C. jejuni infection in poultry.
1. Introduction
Feed additives as growth promoters have a robust impact on the production cost of broilers for covering alterations in profits due to instability in feed costs [1,2]. The extensive use of antibiotics as feed additives for enhancing poultry growth performance and treatment of bacterial infections leads to the emergence of multidrug resistant (MDR) bacterial strains [3,4,5,6,7,8]. Up until now, research is in progress exploring feed additives that can substitute antibiotics as prophylactics and growth promoters in poultry production. Probiotics became prospective antibiotic alternative additives in broilers owing to their impact on growth performance and the well-being of broilers compared to antibiotics [9,10,11]. The main favorable effects of probiotics correlate largely with improved feed bioavailability and digestibility, boosting the immune system, saving health, and providing superior carcass composition [12,13]. They have immunomodulatory roles in enhancing immunity against microbes and preventing exaggerated inflammatory responses, and they serve as biological barriers to protect the epithelial cells from being breached by pathogenic bacteria and to maintain epithelial integrity [14]. As well, probiotic bacteria are able to impede the growth of pathogenic microflora in the gastrointestinal tract of birds owing to their roles in nutrient depletion, blockage of pathogens’ target receptors on epithelial cells, and the creation of natural antibacterial products known as bacteriocins [15,16,17,18]. In addition, the early institution of probiotics in the gut can provide a barrier against foodborne pathogen colonization [19]. Bacillus-based probiotics have unique properties comprising growth promotion, immunomodulation, competitive exclusion [20,21], and production of a variety of extracellular substances and antimicrobial peptides against a wide range of pathogens [22,23]. Bacillus amyloliquefaciens (B. amyloliquefaciens) is a strong Bacillus species that produces numerous extracellular enzymes such as cellulase, metalloproteases, proteases, and α-amylases that augment digestibility and nutrient absorption in addition to overall gut immune functions [24,25]. Moreover, B. amyloliquefaciens produces bacteriocins [26] with a consequence of gut resistance to infection, and it reduces noxious gases emission in chickens [27]. Additionally, B. amyloliquefaciens CECT 5940 possesses a wide range of antimicrobial activity that can ultimately improve broilers’ health [28].
Disruption in the elegant interaction between gut microbiota, intestinal epithelium barrier, and host immunity plays a crucial role in the development of acute bacterial enteritis including campylobacteriosis [29,30], which is caused by Campylobacter jejuni (C. jejuni) and is considered one of the most serious foodborne pathogens causing zoonotic bacterial gastroenteritis in humans. Poultry are a major reservoir for C. jejuni as they provide it with an optimal temperature (42 °C) in their gastrointestinal tract, which is required for its colonization and proliferation [19,31]. Campylobacter jejuni inhabits avian cecum with approximately 106–108 colony forming unit (CFU)/g without clinical illness [32]. Pondering this situation, exploring ongoing promising strategies to reduce C. jejuni colonization in poultry and therefore in humans is needed. Administration of beneficial live probiotics can rescue the intestinal microbiota ecosystem balance, inhibit exaggerated immune responses against innocuous antigens, and hinder pathogenic bacterial colonization by competitive exclusion [33]. Recently, B. amyloliquefaciens CECT 5940 could probably improve growth rate, nutrient intake, gut function, and negative impacts of necrotic enteritis challenge [34]. However, its usage as an alternative to antibiotics is still limited because a harsh gut environment of gastric acids and bile salts may kill it and decrease its bioavailability with a consequent demand for high doses and long-term use of probiotics to exert their curative functions [35,36,37,38]. Therefore, to overcome these limitations and use the advantages of available advancement in technology, recent studies have investigated different formulation methods for effective oral delivery of probiotics to protect them from harsh gut environments and sustain their therapeutic effects [35,39]. Nanoparticle formulations provide a new gateway for achieving a safe delivery system for the ingredients used in feed by increasing their potency and concentration at their target sites [36,40,41]. Moreover, using natural biodegradable biopolymers for encapsulation of these ingredients is widely applied [42]. Based on this technology, co-encapsulation of probiotics strains for their engineering to improve their therapeutic efficiency has been reported [43]. However, the effect of using B. amyloliquefaciens-loaded nanoparticles (BNPs) as alternatives to antibiotics for growth promotion and competing against campylobacteriosis needs to be defined. Therefore, we investigated for the first time in this study the effect of feeding poultry with BNPs on growth performance, barrier function, and immunity of broiler chickens in addition to its efficiency on cecal colonization of C. jejuni and its shedding in poultry excreta.
2. Materials and Methods
2.1. Ethical Statement
All experimental procedures, bird rearing, and management were conducted upon approval from animal resources at the Faculty of Veterinary Medicine, Zagazig University, Egypt according to the rules of the Institutional Animal Care and Use Committee (ZU-IACUC/2/F/337/2022 approval number).
2.2. Bacillus amyloliquefaciens-Loaded Nanoparticles (BNPs)
The strain of B. amyloliquefaciens probiotic CECT 5940 obtained from Evonik Nutrition and Care GmbH was propagated in Luria–Bertani (Oxoid, Hampshire, UK) broth and incubated at 37 °C. Then, B. amyloliquefaciens was stored in the bacterial cryopreservation fluid at −80 °C for further experiments. The BNPs were prepared through incorporation of the bacteria into chitosan (0.4%, w/v) (Sigma-Aldrich, St. Louis, MO, USA) nanoparticles as previously described [44]. The prepared BNPs were stored by freezing at −20 °C in a cryoprotectant agent and were then dried and condensed for 18 h at −40 °C using LyoBeta 25™ freeze-dryer (Telstar, Terrassa, Spain). Finally, the dried cells were kept at 4 °C to be protected from light. The nano size and shape of the formulated BNPs were confirmed using transmission electron microscopy and Fourier-transform infrared spectroscopy characterization (Figure 1).
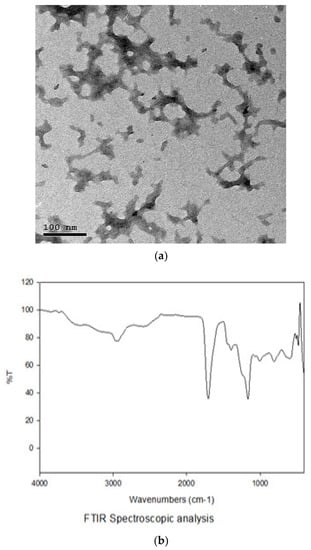
Figure 1.
Characterization of Bacillus amyloliquefaciens-encapsulated nanoparticles using transmission electron microscopy (a) and Fourier-transform infrared spectroscopy (b).
2.3. Birds and Experimental Design Considerations
Two hundred Ross 308 male broiler chickens (42.21 g average initial weight) at 1 day old were provided from a commercial hatchery (Dakahlia Poultry Company, Dakahlia, Egypt). The birds were housed in divided floor pens for separation between groups at the Animal Care Unit at the Faculty of Veterinary Medicine, Zagazig University. The birds were divided randomly into 4 groups; each group had 5 replicates (10 birds each). The first group defined as control was fed the conventional diet and the three other groups were offered the conventional diet supplemented with B. amyloliquefaciens-loaded nanoparticles (BNPs) with three different doses [BNPs I (2.5 × 105 CFU/g of feed), BNPs II (5 × 105 CFU/g of feed) and BNPs III (7.5 × 105 CFU/g of feed)]. Both control and tested groups were fed the conventional or BNPs-containing diets for 35 days during the experimental period. All chicks in control and tested groups were co-housed at the same rearing conditions: temperature (33 ± 1 °C), which was gradually decreased every week until it reached 24 ± 2 °C at the end of the experimental period and humidity (around 60%) was constant during the whole experimental period. The diet was formulated according to nutrition specification of a Ross broiler handbook [45] as presented in Table 1. Chemical analyses of all feed ingredients were performed using the standard method as endorsed by the Association of Official Analytical Chemists, AOAC77 [46]. All animals were provided with ad libitum access to water and food during the whole experimental period.

Table 1.
The ingredients of basal diets and nutrient composition.
2.4. Growth Performance and Mortality Indicators
Individual birds were weighed and feed residues were determined to calculate feed intake for evaluation of different growth parameters of broiler chickens within the growth phases (1–35 days) including body weight gain for each phase [final body weight (g/bird) − initial body weight (g/bird)] and feed conversion ratio (FCR) [feed intake (g/bird)/weight gain (g/bird)] [18,47]. Moreover, mortality rate was recorded during the period from 1 to 35 days.
2.5. Sampling Procedure
At the end of the experimental period, the birds (5 per replicate) were weighed and euthanized by cervical dislocation. The intestinal contents and cecal and fecal samples were then aseptically placed in sterile Eppendorf tubes and kept at −80 °C for further quantification of intestinal microbial populations and evaluation of C. jejuni colonization and shedding via real-time (quantitative) polymerase chain reaction (qPCR) strategies. Moreover, pancreatic and jejunal samples (around 1 cm each) were excised, flushed three times with phosphate buffered saline, and subjected later to genes expression analysis utilizing reverse transcription-quantitative polymerase chain reaction (RT-qPCR) assays.
2.6. Reverse Transcription Quantitative Polymerase Chain Reaction (RT-qPCR)
The expression of genes associated with digestive enzymes [alpha 2A amylase (AMY2A), pancreatic lipase (PNLIP), cholecystokinin (CCK), and chymotrypsin-like elastase family member 1 (CELA1)], barrier functions and antimicrobial defense [beta-defensin 1 (DEFB1), fatty acid-binding protein-2 (FABP-2) and mucin-2 (MUC-2)], and proinflammatory cytokines [interleukin-1beta (IL-1β) and tumor necrosis factor-alpha (TNF-α)] was done. Total RNA was extracted via RNeasy Mini kits (Qiagen, Cat. No. 74104) following the kits’ instructions. RNA purity and concentration were determined using NanoDrop ND-8000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). The cDNA was then synthesized using RevertAidTM H Minus kits (Fermentas Life Science, Pittsburgh, PA, USA) according to manufacturer’s instructions. The RT-qPCR assays were done using SsoAdvanced™ Universal SYBR Green® Supermix (Bio-Rad, 1725274) according to the manufacturer’s instructions and analyzed using Stratagene™ MX3005P qPCR thermocycler (Agilent Technologies, Santa Clara, CA, USA). The sequences of primers of target genes are illustrated in Table 2. Target genes expression was normalized using glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as a housekeeping gene. Relative fold changes in the target genes’ expression were determined using 2−ΔΔCT method [48].
Table 2.
Target genes and primers used for qPCR assays.
2.7. Campylobacter jejuni Challenge Model
A pandrug-resistant (PDR) and multi-virulent field C. jejuni strain was used in this experimental trial. It was previously recovered from cloacal swabs of recently slaughtered broiler chickens based on a previous research by one of the co-authors [51]. The strain was inoculated into Bolton broth (Oxoid, UK) for 48 h at 42 °C in microaerophilic conditions (10% CO2, 85% N2 and 5% O2) using an anaerobic jar (Sigma-Aldrich) with CampyGen sachets (Oxoid). The inoculated broth was then streaked onto modified charcoal cefoperazone deoxycholate agar (Oxoid, UK) plates that were incubated under the previous microaerophilic conditions. The strain was then evinced to be resistant to 24 antimicrobials (amoxicillin, ampicillin, sulbactam-ampicillin, amoxicillin-clavulanic acid, cephalothin, cefoxitin, cefoperazone, cefepime, imipenem, aztreonam, nalidixic acid, ciprofloxacin, trimethoprim-sulfamethoxazole, doxycycline, erythromycin, azithromycin, clarithromycin, tobramycin, gentamicin, amikacin, linezolid, chloramphenicol, colistin, and clindamycin) of 10 different antimicrobial categories being identified as PDR. The stain was also proved to harbor three substantial virulence factors, which have vital roles in its pathogenesis (wlaN, virB11, and flaA) as previously detailed) [52]. The challenge inoculum was adjusted to provide a viable concentration of 108 CFU/mL [4]. At 30 days of age, the birds in all groups including control were orally infected through the crop gavage with 108 CFU/mL of C. jejuni (1 mL per bird). The experimental infection was then affirmed following the appearance of clinical signs via re-isolation and identification of the infecting C. jejuni strain in addition to re-investigating its antimicrobial resistance pattern and virulence genes profiling.
2.8. Real-Time Quantitative Polymerase Chain Reaction (qPCR) for Evaluating C. jejuni Shedding and Colonization
Quantitative PCR assays were carried out to quantify the microbial populations including Bifidobacterium, Enterobacteriaceae, Lactobacillus, and Clostridium species in the avian intestinal contents at the end of the experimental period and to determine C. jejuni colonization in chickens’ cecal samples and its shedding in their fecal samples 3 and 7 days post-infection (33 and 35 d of age). Genome DNA was extracted with QIAamp DNA fast DNA stool kit (Qiagen, Hilden, Germany) adopting the guidance of the manufacturer. Purity and concentration of extracted DNA were determined using NanoDrop (Thermo Fisher Scientific). The sequences of the primers used for quantification of intestinal microbial populations and C. jejuni are illustrated in Table 2. The number of copies of DNA was determined, in triplicate, using Stratagene MX3005P® RT-PCR instrument considering the created standard calibration curves prepared from serially diluted pure bacterial cultures and then the bacterial quantities were expressed as log10 of the CFU/g of the sample.
2.9. Statistical Analysis
The results were evaluated via one-way ANOVA test using SPSS Inc. program version 20 (IBM Corp., Armonk, New York, NY, USA). Differences among the results were expressed as the standard error of the mean (SEM) and variations between means were assessed at a probability level of 5% using Tukey’s test. All graphs were made using GraphPad Prism program Version 8 (San Diego, CA, USA).
3. Results
3.1. Efficacy of B. amyloliquefaciens-Loaded Nanoparticles on Growth Performance and Mortality Percentage of Broiler Chickens
The effect of adding BNPs in different doses to broilers’ diets within the whole experimental period (35 days) is shown in Table 1. Our data revealed that broiler chickens fed BNPs with higher doses (BNPs II and BNPs III) for 35 days exhibited significant (p ˂ 0.05) increases in their body weight gain (2499 and 2569 g/bird, respectively) compared to those fed BNPs I (2430 g/bird) and control diets free of BNPs (2297 g/bird). While there was a reduction in food intake in the groups of birds fed BNPs compared to those in the control group, their efficiency for food utilization was improved (p ˂ 0.05) specifically with higher BNPs doses as indicated by lower FCR in these groups compared to that in the control one. Notably, inclusion of BNPs with higher doses significantly (p ˂ 0.05) decreased mortality rates compared with the challenged group (Table 3).
Table 3.
Effect of feeding male Ross broilers a diet formulated with three different levels of Bacillus amyloliquefaciens-encapsulated nanoparticles (BNPs I, II, III) on growth performance parameters.
3.2. Efficacy of B. amyloliquefaciens-Loaded Nanoparticles on Expression of Digestive Genes Controlling Digestion
The analysis of genes expression associated with digestive enzymes including AMY2a, PNLIP, CELA1, and CCK is illustrated in Figure 2. The results indicated that the expression levels of AMY2a and CCK genes attended their peaks (p ˂ 0.05) following supplementation of BNPs III. A concentration-dependent manner upregulation of PNLIP gene (p ˂ 0.05) was detected following BNPs’ dilatory inclusion. Significant (p ˂ 0.05) upregulation of relative expression of CELA1 gene was noticed in broilers fed diets supplemented with higher levels of BNPs compared to control ones fed diets free of BNPs.
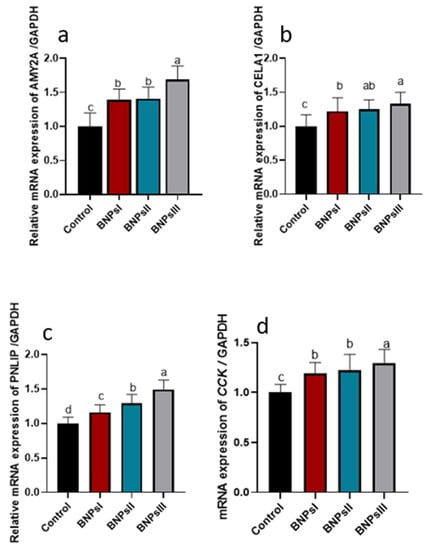
Figure 2.
Effect of feeding Ross broilers (n = 5/group) different levels of Bacillus amyloliquefaciens-encapsulated nanoparticles (BNPs I, II, and III) on the expression of digestive enzymes-related genes compared to birds fed basal diet. Graphs represented the relative mRNA expression of alpha 2A amylase (AMY2A, (a)), chymotrypsin-like elastase family member 1 (CELA1, (b)), pancreatic lipase (PNLIP, (c)), and cholecystokinin (CCK, (d)). The data represented the relative mRNA expression normalized to GAPDH and the significance is p < 0.05. a–d Mean values within columns showing numerous letters differ statistically.
3.3. Efficacy of B. amyloliquefaciens-Loaded Nanoparticles on Broiler Chickens’ Intestinal Microbial Populations
The abundance of intestinal microbial populations following BNPs supplementation is illustrated in Figure 3. As expected, there were increases in the abundance of Lactobacillus and Bifidobacterium and reductions in Clostridium and Enterobacteriaceae as indicated by qPCR in groups of birds fed BNPs compared to control ones. Although the differences among the three groups of birds fed different doses of BNPs did not reach statistical significance in reduction of Enterobacteriaceae and Clostridium species abundance unlike the control group, PNPIII showed the highest tendency in their reduction. Conversely, a high tendency in increasing Bifidobacterium populations was noticed in BNPs-fed groups compared to the control one. Likewise, BNPs III-fed birds had significant (p ˂ 0.05) highest counts for both Lactobacillus and Bifidobacterium species.
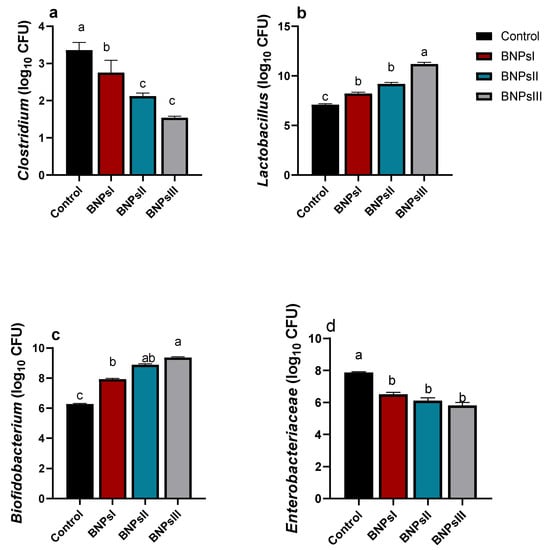
Figure 3.
Effect of feeding Ross broilers (n = 5/group) different levels of Bacillus amyloliquefaciens-encapsulated nanoparticles (BNPs I, II, and III) on the intestinal microbial populations compared to birds fed basal diet. Graphs represented the counts of Clostridium (a), Lactobacillus (b), Bifidobacterium (c), and Enterobacteriaceae (d) expressed as log10 of the CFU/g of the sample and the significance is p < 0.05. a–d Mean values within columns showing numerous letters differ statistically.
3.4. Efficacy of B. amyloliquefaciens-Loaded Nanoparticles on Intestinal Barrier and Cytokines-Associated Genes
Analysis of the relative mRNA expression of genes related to intestinal barrier and antimicrobial defense including DEFB1, FABP-2 and MUC-2, and proinflammatory cytokines comprising IL-β and TNF-α is depicted in Figure 4. We observed an overwhelming increase in the expression of MUC-2 gene after supplementing BNPs in broiler chickens’ diets in a dose-reliant way. In addition, a proportional (p ˂ 0.05) increase in the expression of FABP-2 gene was demonstrated in broiler chickens with increasing the dose of dietary BNPs when compared with the control group. The expression of DEFB1 gene achieved its highest peak in BNPs III-fed birds. Notably, prominent downregulation (p < 0.05) was noticed regarding IL-1β and TNF-α genes, especially after feeding higher BNPs levels when compared to the control group.
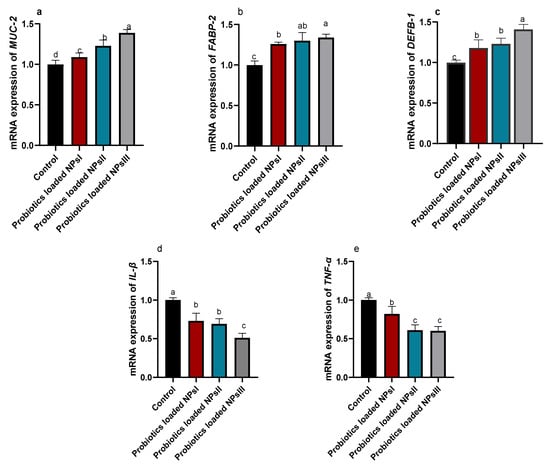
Figure 4.
Effect of feeding Ross broilers (n = 5/group) different levels of Bacillus amyloliquefaciens-encapsulated nanoparticles (BNPs I, II, and III) on the relative mRNA expression of mucin-2 (MUC-2, (a)), fatty acid-binding protein-2 (FABP-2, (b)), beta-defensin 1 (DEFB-1, (c)), interleukin-1beta (IL-1β, (d)), and tumor necrosis factor-alpha (TNF-α, (e)). The data represented the relative mRNA expression normalized to GAPDH and the significance is p < 0.05. a–d Mean values within columns showing numerous letters differ statistically.
3.5. Efficacy of B. amyloliquefaciens-Loaded Nanoparticles on Shedding and Colonization of C. jejuni
Investigating C. jejuni shedding and colonization in fecal and cecal samples 3 and 7 days post-infection using qPCR assay is shown in Figure 5. After 3 and 7 days of C. jejuni infection, there was a tendency of fecal and cecal C. jejuni counts toward reduction within groups of birds fed BNPs with different doses (BNPs I, BNPs II, and BNPs III) compared to those in the control group fed diets free of BNPs. Notably, a lower trend of C. jejuni counts was detected in fecal samples than cecal ones 3 and 7 days post-infection. At both intervals, inclusion of BNPs III in birds’ diets remarkably (p ˂ 0.05) reduced C. jejuni populations considering both sample types unlike the control group. Seven days post-infection, C. jejuni counts were significantly (p ˂ 0.05) reduced in fecal and cecal samples of birds fed BNPs III compared to those in the control group (2.93 and 4.22 CFU/g vs. 6.79 and 8.16 CFU/g), respectively.
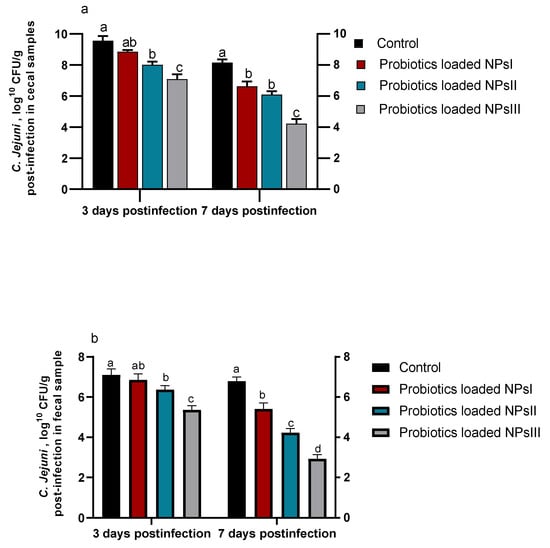
Figure 5.
Effect of feeding Ross broilers (n = 5/group) different levels of Bacillus amyloliquefaciens-encapsulated nanoparticles (BNPs I, II, and III) on colonization and shedding of Campylobacter jejuni (C. jejuni) in cecal (a) and fecal (b) samples at 3 and 7 days post-infection compared to birds fed basal diet and the significance is p < 0.05. a–d Mean values within columns showing numerous letters differ statistically.
4. Discussion
Higher productivity in the poultry industry has been complemented by the balance between nutrition, intestinal health, and animal welfare. Additionally, there are many impacts threatening this productivity such as the emergence of a large assortment of pathogens and bacterial resistance [53]. The concept of using in-feed live probiotics strains in the poultry industry not only exerts a beneficial effect on overall growth performance parameters [54], but also signifies their use in reducing and eliminating the colonization of pathogenic bacteria, such as C. jejuni [19]. Nevertheless, gut colonization and effectiveness of supplemented probiotics depend on various aspects involving the specificity of the strains relative to the host, digestive enzymes, bile acids, availability, nutritional status of the host, intestinal pH, stress, and form of substrate (prebiotics) [55]. Thus, offering probiotic living cells with a physical barrier for escaping from unfavorable environmental conditions is a crucial consequence, which currently has obtained substantial interest [56]. In this study, encapsulation of B. amyloliquefaciens in chitosan nanoparticles had beneficial effects on broiler chicken growth performance and reduced the colonization and shedding of C. jejuni.
C. jejuni represents the most prevalent species accountable for 84% of cases diagnosed recently in Europe [57]. Poultry is the main carrier of the bacteria and is responsible for the most outbreaks of campylobacteriosis through consumption of contaminated poultry meat [43]. The bacteria can colonize and persist in the chicken gut during their lifetime without causing diseases [31]. The emergence of MDR, extensively drug-resistant, and PDR strains [58,59], especially C. jejuni, which is resistant to the main drugs used in the treatment of severe post-acute sequelae of campylobacteriosis (fluoroquinolone and macrolides) evoked a warning sign about hazardous uses of antibiotics for enhancing poultry growth performance and treatment of bacterial infections [60]. The use of probiotics such as Lactobacillus, Bacillus, and Bifidobacterium has been shown to be a promising alternative solution for reduction of pathogen burden in the avian gut and consequently breaks the transmission cycle to humans through the food chain. However, the effects of inclusion of BNPs in poultry diets to decrease the [60] colonization and shedding of C. jejuni need to be defined.
In this study, we have shown that supplementation of B. amyloliquefaciens coated within chitosan nanoparticles, especially at high doses in broilers’ diets for 35 days, promoted their growth performance and increased their body weight gain. Moreover, feed utilization efficiency of broilers was improved as indicated by significant (p < 0.05) reduction in FCR and increase in BWG, especially in BNPs III-fed group. These results are consistent with previous data regarding the improvement in growth performance of broilers fed B. licheniformis spores supplement either alone or combined with mannan oligosaccharide feed additives [61]. Correspondingly, probiotics can modify the intestinal ecosystem by supplying digestion enzymes, reducing pH, and increasing the activity of enzymes in the gastrointestinal tract [62,63]. Similarly, [64] reported a significant improvement in FCR and growth performance of broilers fed dried brewer grain (10%) subjected to probiotics fermentation. Different formulations used for probiotics delivery would affect their absorption; therefore, in this study, chitosan nanoencapsulated probiotics improved their resistance against gastric and bile acids, which consequently increased their bioavailability, absorption, and sustained effectiveness during the overall period of experiments (35 days). The growth performance of birds was strongly correlated with the digestibility and utilization efficiency of feed [16]. Moreover, it has been known that dietary composition of broilers’ rations could modulate the expression of digestive enzymes and nutrient transport-related genes [57,65]. Therefore, our study revealed that a group of birds fed BNPs III for 35 days exhibited significant increase in relative mRNA expression of pancreatic digestive enzyme-linked genes, such as AMY2A, PNLIP, CELA1, and CCK. Aligned with our data, [64] stated that feeding microbially fermented dried brewer grains not only improved growth performance and carcass dressing, but also stimulated expression of pancreatic digestive enzymes encoding genes as AMY2A, PNLIP, CELA1, and CCK.
Microbiota and their metabolites prompt gut enteroendocrine cells to secrete gut hormones, which consequently influence metabolism [66]. Our results indicated the ability of BNPs to stimulate gut hormones, such as CCK, which plays important roles in gastric motility, appetite, bile acids, and pancreatic enzymes release [66,67]. Although less is known about how probiotics regulate CCK secretion, a recent research showed that feeding broilers for 42 days on microbially fermented olive pomace via two stages of solid fermentation using Bacillus subtilis followed by Lactobacillus casei upregulated the genes encoding pancreatic enzymes: CCK, AMY2A, PNLIP, and CELA1 [18]. Furthermore, dietary inclusion of microbially fermented soybean meal enhanced pancreatic enzymes’ activities in broilers [68]. Moreover, dietary supplementation of B. amyloliquefaciens had significant efficacy in enhancing chymotrypsin, amylase, and lipase activities compared to groups fed antibiotics alone [69]. Similarly, it has been reported that B. amyloliquefaciens (2 × 105 CFU/g diet) pretreatment either alone or combined with mannan oligosaccharides improved metabolic activity of intestinal cell energy-related genes through upregulation of mRNA genes expression of enzymes implicated in protein digestion. The mechanism by which probiotics regulate digestive enzymes secretion needs to be deciphered. However, this may be attributed to the role of probiotics in restoring intestinal normal architecture and achievement of larger healthy surface area for nutrient assimilation [70]. Moreover, the role of probiotics in increasing the relative mRNA expression of pancreatic digestive enzymes could be related to their vital role in the activity of enteroendocrine cells to express the pancreatic enzymes encoding genes. Additionally, increasing the expression of genes encoding pancreatic lipase associated enzymes (PNLIP and CCK) could be secondary to the production of probiotics’ metabolites including short chain fatty acids [71].
Our data indicated increases in the abundance of Bifidobacterium and Lactobacillus species, which was accompanied by reduction in Enterobacteriaceae and Clostridium species populations. In parallel, probiotics not only exert their beneficial effects on host resistance through improving its immunity, epithelial function, and competitive exclusion of pathogens burden within the avian gut, but also through their effects on rescuing the normal microbial populations of the intestine via favoring the growth of beneficial commensals and inhibiting that of the opportunistic ones [72]. Moreover, Bao et al. [73]. reported an increase in the abundance of Lactobacillus species combined with reduction in Enterobacteriaceae and Clostridiales counts after inclusion of B. amyloliquefaciens in poultry diet compared to poultry fed a basal diet [73]. The increase of Lactobacillus and Bifidobacterium within the cecal contents of broilers may be attributed to the ability of BNPs to produce extracellular enzymes as phytase, amylase, xylanase, and β-glucanase, which could improve food digestibility and utilization [74]. In addition, accumulation of short chain fatty acids produced within microbial metabolic processes confers a suitable pH that would support the growth of beneficial bacteria and prevent that of harmful ones [73,74]. Both Lactobacillus and Bifidobacterium provide substrates for augmenting the growth of butyrate-producing bacteria, which stimulates epithelial regeneration and improves host energy metabolism [61].
Improvement of epithelial barrier functions as indicated by upregulation of MUC2 and IgA genes expression was observed following administration of Bacillus coagulans to chickens [75]. Consistent with this finding, we found that improvement of growth performance of birds was correlated with upregulation of epithelial barrier functions-related genes including MUC2, DEFB1, and FABP2, specifically, in birds supplemented with BNPs III in their diet for 35 days. Avian defensin represents the innate antimicrobial peptide with a wide range of spectrum against bacteria, fungi, and protozoa [76,77]. Moreover, its ability to kill bacteria is through alteration in cell membrane permeability and immunomodulatory functions involving chemoattraction for leukocytes to the injury site and elimination of bacterial lipopolysaccharides and lipoteichoic acids triggering proinflammatory response [76]. The upregulation of avian defensin following BNPs supplementation suggests its role as an antimicrobial peptide against MDR bacteria as proved previously [78]. Mucus is the first physical barrier protecting epithelial cells from microbial translocation and inflammatory response [79]. Likewise, intestinal barrier is strengthened by a glycosylated mucin-rich layer produced by goblet cells [80]. Supporting this view, probiotics have been proven to strengthen the integrity of the intestinal barrier by elevating the number of goblet cells that support the mucus layer [81]. A previous study described that many Bacillus species were evidenced to upregulate the expression of intestinal mucin [82] as demonstrated in our study. FABP-2 gene expression has been indicated as a biomarker for intestinal barrier in broilers’ gut and its higher expression signifies its superior role in intestinal function [83]. Similarly, our data showed upregulation of FABP-2 relative mRNA expression, which suggested that BNPs enhanced epithelial integrity and functions. In the present study, downregulation of intestinal gene expression levels of proinflammatory cytokines (TNF-α and IL-1β) were observed in broilers fed higher levels of BNPs. This observation indicated their suppressive role on proinflammatory cytokines, which in turn counteracted the inflammation [80,84,85].
Several studies evaluated the in vitro and in vivo anti-campylobacter activities of different probiotics strains [15,19]. However, the effect of dietary inclusion of probiotics-loaded nanocarriers on the improvement of birds’ resistance against C. jejuni and their interaction with host epithelial cells need to be explicitly defined. Our data further demonstrated that the aforementioned beneficial effects of BNPs, especially with higher doses, led to significant reduction in cecal colonization and fecal shedding of C. jejuni. These data suggested the ability of BNPs to competitively exclude C. jejuni from broilers’ intestine and prevent its colonization. From what has been declared above, our data explained several possible mechanisms by which BNPs could mediate the exclusion and inhibition of C. jejuni colonization and translocation within the avian gut. These mechanisms are mediated firstly through the improvement of nutrient digestibility and assimilation and secondly via their effects on improving intestinal morphology and functions through upregulation of FABP2 and MUC2 genes, which increased mucus secretion and prevented bacterial adhesion. Lastly, they exert antimicrobial impacts either through secretion of antimicrobial peptides, such as β-defensin, that breaks down the bacterial cell wall and consequently activates the innate immune response to control inflammatory process and prevent bacterial translocation.
5. Conclusions
Our data suggested that encapsulation of B. amyloliquefaciens as a probiotic bacterium by chitosan nanoparticles enhanced its bioavailability and maintained its beneficial effects in the broiler chicken’s gut. The augmented growth promoter properties of BNPs were detected in our study and supported by higher expression of digestive enzymes-related genes. Friendly gut bacterial species including Bifidobacterium and Lactobacillus species outnumbered the unfriendly ones following inclusion of dietary BNPs. Supplementation of BNPs could also enhanced gut barrier functions and consequently decrease C. jejuni colonization with a superior outcome of minimizing its fecal shedding.
Author Contributions
Conceptualization, H.I., D.I., S.E.S., A.W., R.M.E.-T. and M.I.A.E.-H.; methodology, D.I., A.W., R.M.E.-T., W.R.E.-G., B.A.A., B.A.-H.Y.A., S.M.A.-R. and M.I.A.E.-H.; software, D.I., A.W., R.M.E.-T., W.R.E.-G., B.A.A., S.E.S. and M.I.A.E.-H.; validation, H.I., D.I., R.M.E.-T., W.R.E.-G., B.A.A. and M.I.A.E.-H.; formal analysis, D.I. and M.I.A.E.-H.; investigation, H.I., D.I., A.W., R.M.E.-T., W.R.E.-G., S.E.S., B.A.A., B.A.-H.Y.A., S.M.A.-R. and M.I.A.E.-H.; resources, D.I., A.W., R.M.E.-T., W.R.E.-G., B.A.A., S.M.A.-R. and M.I.A.E.-H.; data curation, H.I., D.I., A.W., R.M.E.-T., W.R.E.-G., B.A.A., S.E.S., B.A.-H.Y.A., S.M.A.-R. and M.I.A.E.-H.; writing—original draft preparation, H.I., S.E.S., D.I., A.W. and R.M.E.-T.; writing—review and editing, D.I. and M.I.A.E.-H.; visualization, H.I., D.I., A.W., R.M.E.-T., W.R.E.-G., B.A.A., B.A.-H.Y.A., S.M.A.-R. and M.I.A.E.-H.; supervision, H.I., D.I., A.W., B.A.-H.Y.A., S.M.A.-R. and M.I.A.E.-H.; project administration, H.I., D.I., A.W., R.M.E.-T., W.R.E.-G., B.A.A., B.A.-H.Y.A., S.M.A.-R. and M.I.A.E.-H.; funding acquisition, D.I., A.W., R.M.E.-T., W.R.E.-G., S.E.S., B.A.A. and M.I.A.E.-H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Deputyship for Research and Innovation, Ministry of Education in Saudi Arabia through Project number (INST007).
Institutional Review Board Statement
All experimental procedures, bird rearing, and management were conducted upon approval from animal resources at the Faculty of Veterinary Medicine, Zagazig University, Egypt according to the rules of the Institutional Animal Care and Use Committee (ZU-IACUC/2/F/337/2022 approval number).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data presented in this research are offered upon request from the corresponding author.
Acknowledgments
The authors extend their appreciation to the Deputyship for Research and Innovation, Ministry of Education in Saudi Arabia, for funding this research work (Project number INST007).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ibrahim, D.; Ismail, T.A.; Khalifa, E.; El-Kader, A.; Shaimaa, A.; Mohamed, D.I.; Mohamed, D.T.; Shahin, S.E.; El-Hamid, A.; Marwa, I. Supplementing Garlic Nanohydrogel Optimized Growth, Gastrointestinal Integrity and Economics and Ameliorated Necrotic Enteritis in Broiler Chickens Using a Clostridium perfringens Challenge Model. Animals 2021, 11, 2027. [Google Scholar] [CrossRef] [PubMed]
- Kishawy, A.T.; Al-Khalaifah, H.S.; Nada, H.S.; Roushdy, E.M.; Zaglool, A.W.; Ahmed Ismail, T.; Ibrahim, S.M.; Ibrahim, D. Black Pepper or Radish Seed Oils in a New Combination of Essential Oils Modulated Broiler Chickens’ Performance and Expression of Digestive Enzymes, Lipogenesis, Immunity, and Autophagy-Related Genes. Vet. Sci. 2022, 9, 43. [Google Scholar] [CrossRef]
- Poudel, S.; Li, T.; Chen, S.; Zhang, X.; Cheng, W.-H.; Sukumaran, A.T.; Kiess, A.S.; Zhang, L. Prevalence, Antimicrobial Resistance, and Molecular Characterization of Campylobacter Isolated from Broilers and Broiler Meat Raised without Antibiotics. Microbiol. Spectr. 2022, 10, e00251-22. [Google Scholar] [CrossRef]
- Aljazzar, A.; Abd El-Hamid, M.I.; El-Malt, R.M.; El-Gharreb, W.R.; Abdel-Raheem, S.M.; Ibrahim, A.M.; Abdelaziz, A.M.; Ibrahim, D. Prevalence and Antimicrobial Susceptibility of Campylobacter Species with Particular Focus on the Growth Promoting, Immunostimulant and Anti-Campylobacter jejuni Activities of Eugenol and Trans-Cinnamaldehyde Mixture in Broiler Chickens. Animals 2022, 12, 905. [Google Scholar] [CrossRef]
- El-Hamid, A.; Marwa, I.; Sewid, A.H.; Samir, M.; Hegazy, W.A.; Bahnass, M.M.; Mosbah, R.A.; Ghaith, D.M.; Khalifa, E.; Ramadan, H. Clonal diversity and epidemiological characteristics of ST239-MRSA strains. Front. Cell. Infect. Microbiol. 2022, 12, 241. [Google Scholar] [CrossRef]
- Ahmed, H.A.; Tahoun, A.B.; Abou Elez, R.M.; Abd El-Hamid, M.I.; Abd Ellatif, S.S. Prevalence of Yersinia enterocolitica in milk and dairy products and the effects of storage temperatures on survival and virulence gene expression. Int. Dairy J. 2019, 94, 16–21. [Google Scholar] [CrossRef]
- Abd El-Hamid, M.I.; Awad, N.F.; Hashem, Y.M.; Abdel-Rahman, M.A.; Abdelaziz, A.M.; Mohammed, I.A.; Abo-Shama, U.H. In vitro evaluation of various antimicrobials against field Mycoplasma gallisepticum and Mycoplasma synoviae isolates in Egypt. Poult. Sci. 2019, 98, 6281–6288. [Google Scholar] [CrossRef]
- Khater, S.I.; Lotfy, M.M.; Alandiyjany, M.N.; Alqahtani, L.S.; Zaglool, A.W.; Althobaiti, F.; Ismail, T.A.; Soliman, M.M.; Saad, S.; Ibrahim, D. Therapeutic Potential of Quercetin Loaded Nanoparticles: Novel Insights in Alleviating Colitis in an Experimental DSS Induced Colitis Model. Biomedicines 2022, 10, 1654. [Google Scholar] [CrossRef]
- Abd El-Hamid, M.I.; El-Sayed, M.; Ali, A.R.; Abdallah, H.; Arnaout, M.I.; El-mowalid, G.A. Marjoram extract down-regulates the expression of Pasteurella multocida adhesion, colonization and toxin genes: A potential mechanism for its antimicrobial activity. Comp. Immunol. Microbiol. Infect. Dis. 2019, 62, 101–108. [Google Scholar] [CrossRef]
- Elfaky, M.A.; Abdel-Hamid, M.I.; Khalifa, E.; Alshareef, W.A.; Mosbah, R.A.; Elazab, S.T.; Ghoneim, M.M.; Al-Sanea, M.M.; Bendary, M.M. Innovative next-generation therapies in combating multi-drug-resistant and multi-virulent Escherichia coli isolates: Insights from in vitro, in vivo, and molecular docking studies. Appl. Microbiol. Biotechnol. 2022, 106, 1691–1703. [Google Scholar] [CrossRef] [PubMed]
- Awad, N.F.; Hashem, Y.M.; Elshater, N.S.; Khalifa, E.; Hamed, R.I.; Nossieur, H.H.; Abd-Allah, E.M.; Elazab, S.T.; Nassan, M.A.; Abd El-Hamid, M.I. Therapeutic potentials of aivlosin and/or zinc oxide nanoparticles against Mycoplasma gallisepticum and/or Ornithobacterium rhinotracheale with a special reference to the effect of zinc oxide nanoparticles on aivlosin tissue residues: An in vivo approach. Poult. Sci. 2022, 101, 101884. [Google Scholar] [CrossRef]
- Dhama, K.; Verma, V.; Sawant, P.; Tiwari, R.; Vaid, R.; Chauhan, R. Applications of probiotics in poultry: Enhancing immunity and beneficial effects on production performances and health: A review. J. Immunol. Immunopathol. 2011, 13, 1–19. [Google Scholar]
- Farnell, M.; Donoghue, A.; De Los Santos, F.S.; Blore, P.; Hargis, B.; Tellez, G.; Donoghue, D. Upregulation of oxidative burst and degranulation in chicken heterophils stimulated with probiotic bacteria. Poult. Sci. 2006, 85, 1900–1906. [Google Scholar] [CrossRef]
- Suryadi, U.; Prasetyo, A. Probiotics based on Local Microorganism as a subtitute of Antibiotic Growth Promotor (AGP) on Broiler productivity. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Chiyoda City, Tokyo, Japan, 2018; p. 012019. [Google Scholar]
- Śmiałek, M.; Kowalczyk, J.; Koncicki, A. The use of probiotics in the reduction of Campylobacter spp. Prevalence in poultry. Animals 2021, 11, 1355. [Google Scholar] [CrossRef] [PubMed]
- Al-Khalaifah, H.S.; Shahin, S.E.; Omar, A.E.; Mohammed, H.A.; Mahmoud, H.I.; Ibrahim, D. Effects of graded levels of microbial fermented or enzymatically treated dried brewer’s grains on growth, digestive and nutrient transporter genes expression and cost effectiveness in broiler chickens. BMC Vet. Res. 2020, 16, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, D.; Abdelfattah-Hassan, A.; Arisha, A.H.; Abd El-Aziz, R.M.; Sherief, W.R.; Adli, S.H.; El Sayed, R.; Metwally, A.E. Impact of feeding anaerobically fermented feed supplemented with acidifiers on its quality and growth performance, intestinal villi and enteric pathogens of mulard ducks. Livest. Sci. 2020, 242, 104299. [Google Scholar] [CrossRef]
- Ibrahim, D.; Moustafa, A.; Shahin, S.; Sherief, W.; Farag, M.; Nassan, M. Impact of fermented or enzymatically fermented dried olive pomace on growth, expression of digestive enzymes and glucose transporters genes, oxidative stability of frozen meat and economic efficiency of broiler chickens. Front. Vet. Sci. 2021, 8, 442. [Google Scholar] [CrossRef]
- Deng, W.; Dittoe, D.K.; Pavilidis, H.O.; Chaney, W.E.; Yang, Y.; Ricke, S.C. Current perspectives and potential of probiotics to limit foodborne Campylobacter in poultry. Front. Microbiol. 2020, 11, 583429. [Google Scholar] [CrossRef] [PubMed]
- Cutting, S.M. Bacillus probiotics. Food Microbiol. 2011, 28, 214–220. [Google Scholar] [CrossRef]
- Murshed, M.; Abudabos, A. Effects of the dietary inclusion of a probiotic, a prebiotic or their combinations on the growth performance of broiler chickens. Braz. J. Poult. Sci. 2015, 17, 99–103. [Google Scholar] [CrossRef]
- Korenblum, E.; von Der Weid, I.; Santos, A.; Rosado, L.; Sebastián, G.; Coutinho, C.; Magalhaes, F.; De Paiva, M.; Seldin, L. Production of antimicrobial substances by Bacillus subtilis LFE-1, B. firmus H2O-1 and B. licheniformis T6-5 isolated from an oil reservoir in Brazil. J. Appl. Microbiol. 2005, 98, 667–675. [Google Scholar] [CrossRef]
- Omar, A.E.; Al-Khalaifah, H.S.; Ismail, T.A.; El-Aziz, A.; Reda, M.; El-Mandrawy, S.A.; Shalaby, S.I.; Ibrahim, D. Performance, serum biochemical and immunological parameters, and digestive enzyme and intestinal barrier-related gene expression of broiler chickens fed fermented fava bean by-products as a substitute for conventional feed. Front. Vet. Sci. 2021, 8, 740. [Google Scholar] [CrossRef]
- Lee, Y.-J.; Kim, B.-K.; Lee, B.-H.; Jo, K.-I.; Lee, N.-K.; Chung, C.-H.; Lee, Y.-C.; Lee, J.-W. Purification and characterization of cellulase produced by Bacillus amyoliquefaciens DL-3 utilizing rice hull. Bioresour. Technol. 2008, 99, 378–386. [Google Scholar] [CrossRef]
- Alkushi, A.; Abdelfattah-Hassan, A.; Eldoumani, H.; Elazab, S.T.; Mohamed, S.A.; Metwally, A.S.; El-Shetry, E.S.; Saleh, A.A.; ElSawy, N.A.; Ibrahim, D. Probiotics-loaded nanoparticles attenuated colon inflammation, oxidative stress, and apoptosis in colitis. Sci. Rep. 2022, 12, 5116. [Google Scholar] [CrossRef] [PubMed]
- Ulyanova, V.; Mahmud, R.S.; Dudkina, E.; Vershinina, V.; Domann, E.; Ilinskaya, O. Phylogenetic distribution of extracellular guanyl-preferring ribonucleases renews taxonomic status of two Bacillus strains. J. Gen. Appl. Microbiol. 2016, 62, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.Y.; Wu, Z.L.; Wang, G.Z.; Liu, W.C. The effect of Bacillus amyloliquefaciens on productive performance of laying hens. Ital. J. Anim. Sci. 2018, 17, 436–441. [Google Scholar] [CrossRef]
- Lei, X.; Piao, X.; Ru, Y.; Zhang, H.; Péron, A.; Zhang, H. Effect of Bacillus amyloliquefaciens-based direct-fed microbial on performance, nutrient utilization, intestinal morphology and cecal microflora in broiler chickens. Asian-Australas. J. Anim. Sci. 2015, 28, 239. [Google Scholar] [CrossRef]
- Kalischuk, L.D.; Buret, A.G. A role for Campylobacter jejuni-induced enteritis in inflammatory bowel disease? Am. J. Physiol.-Gastrointest. Liver Physiol. 2010, 298, G1–G9. [Google Scholar] [CrossRef]
- Ibrahim, D.; Sewid, A.H.; Arisha, A.H.; Abd El-Fattah, A.H.; Abdelaziz, A.M.; Al-Jabr, O.A.; Kishawy, A.T. Influence of Glycyrrhiza glabra Extract on Growth, Gene Expression of Gut Integrity, and Campylobacter jejuni Colonization in Broiler Chickens. Front. Vet. Sci. 2020, 7, 612063. [Google Scholar] [CrossRef] [PubMed]
- Hermans, D.; Pasmans, F.; Heyndrickx, M.; Van Immerseel, F.; Martel, A.; Van Deun, K.; Haesebrouck, F. A tolerogenic mucosal immune response leads to persistent Campylobacter jejuni colonization in the chicken gut. Crit. Rev. Microbiol. 2012, 38, 17–29. [Google Scholar] [CrossRef]
- Abd El-Hamid, M.I.; Abd El-Aziz, N.K.; Samir, M.; El-Naenaeey, E.-s.Y.; Abo Remela, E.M.; Mosbah, R.A.; Bendary, M.M. Genetic diversity of Campylobacter jejuni isolated from avian and human sources in Egypt. Front. Microbiol. 2019, 10, 2353. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.R.; Aggarwal, P.; Costa, R.G.; Cole, A.M.; Trinchieri, G. Targeting the gut microbiota for cancer therapy. Nat. Rev. Cancer 2022, 22, 703–722. [Google Scholar] [CrossRef] [PubMed]
- Gharib-Naseri, K.; Dorigam, J.C.; Doranalli, K.; Morgan, N.; Swick, R.A.; Choct, M.; Wu, S.-B. Bacillus amyloliquefaciens CECT 5940 improves performance and gut function in broilers fed different levels of protein and/or under necrotic enteritis challenge. Anim. Nutr. 2021, 7, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Jampilek, J.; Kos, J.; Kralova, K. Potential of nanomaterial applications in dietary supplements and foods for special medical purposes. Nanomaterials 2019, 9, 296. [Google Scholar] [CrossRef]
- Zhou, J.; Li, M.; Chen, Q.; Li, X.; Chen, L.; Dong, Z.; Zhu, W.; Yang, Y.; Liu, Z.; Chen, Q. Programmable probiotics modulate inflammation and gut microbiota for inflammatory bowel disease treatment after effective oral delivery. Nat. Commun. 2022, 13, 3432. [Google Scholar] [CrossRef]
- Hegazy, W.A.; Salem, I.M.; Alotaibi, H.F.; Khafagy, E.-S.; Ibrahim, D. Terazosin Interferes with Quorum Sensing and Type Three Secretion System and Diminishes the Bacterial Espionage to Mitigate the Salmonella Typhimurium Pathogenesis. Antibiotics 2022, 11, 465. [Google Scholar] [CrossRef]
- Alandiyjany, M.N.; Abdelaziz, A.S.; Abdelfattah-Hassan, A.; Hegazy, W.A.; Hassan, A.A.; Elazab, S.T.; Mohamed, E.A.; El-Shetry, E.S.; Saleh, A.A.; ElSawy, N.A. Novel In Vivo Assessment of Antimicrobial Efficacy of Ciprofloxacin Loaded Mesoporous Silica Nanoparticles against Salmonella typhimurium Infection. Pharmaceuticals 2022, 15, 357. [Google Scholar] [CrossRef]
- Anselmo, A.C.; McHugh, K.J.; Webster, J.; Langer, R.; Jaklenec, A. Biomaterials: Layer-by-Layer Encapsulation of Probiotics for Delivery to the Microbiome (Adv. Mater. 43/2016). Adv. Mater. 2016, 28, 9442. [Google Scholar] [CrossRef]
- Ibrahim, D.; Arisha, A.H.; Khater, S.I.; Gad, W.M.; Hassan, Z.; Abou-Khadra, S.H.; Mohamed, D.I.; Ahmed Ismail, T.; Gad, S.A.; Eid, S.A. Impact of Omega-3 Fatty Acids Nano-Formulation on Growth, Antioxidant Potential, Fillet Quality, Immunity, Autophagy-Related Genes and Aeromonas hydrophila Resistance in Nile Tilapia (Oreochromis niloticus). Antioxidants 2022, 11, 1523. [Google Scholar] [CrossRef]
- Ibrahim, D.; Shahin, S.E.; Alqahtani, L.S.; Hassan, Z.; Althobaiti, F.; Albogami, S.; Soliman, M.M.; El-Malt, R.M.; Al-Harthi, H.F.; Alqadri, N. Exploring the Interactive Effects of Thymol and Thymoquinone: Moving towards an Enhanced Performance, Gross Margin, Immunity and Aeromonas sobria Resistance of Nile Tilapia (Oreochromis niloticus). Animals 2022, 12, 3034. [Google Scholar] [CrossRef]
- Lin, S.; Mukherjee, S.; Li, J.; Hou, W.; Pan, C.; Liu, J. Mucosal immunity–mediated modulation of the gut microbiome by oral delivery of probiotics into Peyer’s patches. Sci. Adv. 2021, 7, eabf0677. [Google Scholar] [CrossRef]
- Rawson, T.; Colles, F.M.; Terry, J.C.D.; Bonsall, M.B. Mechanisms of biodiversity between Campylobacter sequence types in a flock of broiler–breeder chickens. Ecol. Evol. 2022, 12, e8651. [Google Scholar] [CrossRef]
- Alkushi, A.G.; Elazab, S.T.; Abdelfattah-Hassan, A.; Mahfouz, H.; Salem, G.A.; Sheraiba, N.I.; Mohamed, E.A.; Attia, M.S.; El-Shetry, E.S.; Saleh, A.A. Multi-Strain-Probiotic-Loaded Nanoparticles Reduced Colon Inflammation and Orchestrated the Expressions of Tight Junction, NLRP3 Inflammasome and Caspase-1 Genes in DSS-Induced Colitis Model. Pharmaceutics 2022, 14, 1183. [Google Scholar] [CrossRef] [PubMed]
- Aviagen, W. Ross 308: Broiler’s management and nutrition specification. 2018. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International, Association of Official Analytical Chemists; AOAC: Washington, DC, USA, 2012. [Google Scholar]
- Ibrahim, D.; El Sayed, R.; Abdelfattah-Hassan, A.; Morshedy, A. Creatine or guanidinoacetic acid? Which is more effective at enhancing growth, tissue creatine stores, quality of meat, and genes controlling growth/myogenesis in Mulard ducks. J. Appl. Anim. Res. 2019, 47, 159–166. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kheravii, S.K.; Ionescu, C.; Blanchard, A.; Barekatain, R.; Bajagai, Y.S.; Wu, S.-B. A microencapsulated mixture of eugenol and garlic tincture supplementation mitigates the effect of necrotic enteritis on intestinal integrity and increases goblet cells in broilers. Microorganisms 2021, 9, 1451. [Google Scholar] [CrossRef] [PubMed]
- Mirhosseini, S.Z.; Seidavi, A.; Shivazad, M.; Chamani, M.; Sadeghi, A.A.; Pourseify, R. Detection of Clostridium sp. and its relation to different ages and gastrointestinal segments as measured by molecular analysis of 16S rRNA genes. Braz. Arch. Biol. Technol. 2010, 53, 69–76. [Google Scholar] [CrossRef]
- Ammar, A.M.; El-Naenaeey, E.-S.Y.; El-Malt, R.; El-Gedawy, A.A.; Khalifa, E.; Elnahriry, S.S.; El-Hamid, A.; Marwa, I. Prevalence, antimicrobial susceptibility, virulence and genotyping of Campylobacter jejuni with a special reference to the anti-virulence potential of Eugenol and beta-resorcylic acid on some multi-drug resistant isolates in Egypt. Animals 2021, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Ammar, A.M.; El-Naenaeey, E.-S.Y.; Abd El-Hamid, M.I.; El-Gedawy, A.A.; Elmalt, R.M. Campylobacter as a Major Foodborne Pathogen: A Review of Its Characteristics, Pathogenesis, Antimicrobial Resistance and Control. J. Microbiol. Biotechnol. Food Sci. 2021, 10, 609–619. [Google Scholar] [CrossRef]
- Lutful Kabir, S. The role of probiotics in the poultry industry. Int. J. Mol. Sci. 2009, 10, 3531–3546. [Google Scholar] [CrossRef]
- Bai, S.; Wu, A.; Ding, X.; Lei, Y.; Bai, J.; Zhang, K.; Chio, J. Effects of probiotic-supplemented diets on growth performance and intestinal immune characteristics of broiler chickens. Poult. Sci. 2013, 92, 663–670. [Google Scholar] [CrossRef]
- Bomba, A.; Nemcova, R.; Gancarcikova, S.; Herich, R.; Guba, P.; Mudronova, D. Improvement of the probiotic effect of micro-organisms by their combination with maltodextrins, fructo-oligosaccharides and polyunsaturated fatty acids. Br. J. Nutr. 2002, 88, S95–S99. [Google Scholar] [CrossRef] [PubMed]
- Fávaro-Trindade, C.S.; Heinemann, R.J.B.; Pedroso, D.d.L. Developments in probiotic encapsulation. CABI Rev. 2011, 2021, 1–8. [Google Scholar] [CrossRef]
- Soro, A.B.; Whyte, P.; Bolton, D.J.; Tiwari, B.K. Strategies and novel technologies to control Campylobacter in the poultry chain: A review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1353–1377. [Google Scholar] [CrossRef]
- Bendary, M.M.; Abd El-Hamid, M.I.; El-Tarabili, R.M.; Hefny, A.A.; Algendy, R.M.; Elzohairy, N.A.; Ghoneim, M.M.; Al-Sanea, M.M.; Nahari, M.H.; Moustafa, W.H. Clostridium perfringens Associated with Foodborne Infections of Animal Origins: Insights into Prevalence, Antimicrobial Resistance, Toxin Genes Profiles, and Toxinotypes. Biology 2022, 11, 551. [Google Scholar] [CrossRef] [PubMed]
- Ammar, A.M.; Abd El-Hamid, M.I.; Mohamed, Y.H.; Mohamed, H.M.; Al-Khalifah, D.H.; Hozzein, W.N.; Selim, S.; El-Neshwy, W.M.; El-Malt, R.M. Prevalence and antimicrobial susceptibility of bovine Mycoplasma species in Egypt. Biology 2022, 11, 1083. [Google Scholar] [CrossRef]
- Ammar, A.M.; El-Hamid, A.; Marwa, I.; El-Malt, R.; Azab, D.S.; Albogami, S.; Al-Sanea, M.M.; Soliman, W.E.; Ghoneim, M.M.; Bendary, M.M. Molecular detection of fluoroquinolone resistance among multidrug-, extensively drug-, and pan-drug-resistant Campylobacter species in Egypt. Antibiotics 2021, 10, 1342. [Google Scholar] [CrossRef]
- Emami, N.K.; Calik, A.; White, M.B.; Kimminau, E.A.; Dalloul, R.A. Effect of probiotics and multi-component feed additives on microbiota, gut barrier and immune responses in broiler chickens during subclinical necrotic enteritis. Front. Vet. Sci. 2020, 7, 572142. [Google Scholar] [CrossRef]
- Kabir, S.L. The dynamics of probiotics in enhancing poultry meat production and quality. Department of Microbiology and Hygiene, Faculty of Veterinary science, Bangladesh Agricultural University. Int. J. Poult. Sci. 2009, 3, 361–364. [Google Scholar]
- Rehman, A.; Arif, M.; Sajjad, N.; Al-Ghadi, M.; Alagawany, M.; Abd El-Hack, M.; Alhimaidi, A.; Elnesr, S.; Almutairi, B.; Amran, R. Dietary effect of probiotics and prebiotics on broiler performance, carcass, and immunity. Poult. Sci. 2020, 99, 6946–6953. [Google Scholar] [CrossRef]
- Khalili, A.; Tabeidian, S.A.; Toghyani, M.; Ghalamkari, G.; Bahrami, Y. Effect of different levels of brewer’s dried grains and enzyme on performance, protein digestibility, immune response and performance of broilers. Int. J. Acad. Res. 2011, 3, 1153–1157. [Google Scholar]
- Lin, P.; Shih, B.; Hsu, J. Effects of different sources of dietary non-starch polysaccharides on the growth performance, development of digestive tract and activities of pancreatic enzymes in goslings. Br. Poult. Sci. 2010, 51, 270–277. [Google Scholar] [CrossRef]
- Martin, A.M.; Sun, E.W.; Rogers, G.B.; Keating, D.J. The influence of the gut microbiome on host metabolism through the regulation of gut hormone release. Front. Physiol. 2019, 10, 428. [Google Scholar] [CrossRef] [PubMed]
- Kheravii, S.K.; Swick, R.A.; Choct, M.; Wu, S.-B. Upregulation of genes encoding digestive enzymes and nutrient transporters in the digestive system of broiler chickens by dietary supplementation of fiber and inclusion of coarse particle size corn. BMC Genom. 2018, 19, 208. [Google Scholar] [CrossRef]
- Soumeh, E.; Mohebodini, H.; Toghyani, M.; Shabani, A.; Ashayerizadeh, A.; Jazi, V. Synergistic effects of fermented soybean meal and mannan-oligosaccharide on growth performance, digestive functions, and hepatic gene expression in broiler chickens. Poult. Sci. 2019, 98, 6797–6807. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Cao, X.; Wu, Y.; Mei, X.; Xu, H.; Wang, Y.; Zhang, X.; Gong, L.; Li, W. Effects of probiotic Bacillus as an alternative of antibiotics on digestive enzymes activity and intestinal integrity of piglets. Front. Microbiol. 2018, 9, 2427. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.; Yu, Y.-H.; Hsiao, F.S.-H.; Dybus, A.; Ali, I.; Hsu, H.-C.; Cheng, Y.-H. Probiotics as a Friendly Antibiotic Alternative: Assessment of Their Effects on the Health and Productive Performance of Poultry. Fermentation 2022, 8, 672. [Google Scholar] [CrossRef]
- Matur, E.; Ulker, C.; Arslan, M.; Elif, E.; Akyazi, I.; Evren, E. The effects of Enterococcus faecium NCIMB10415 on the development of pancreas and small intestine and on activity of pancreatic digestive enzymes in broiler chickens. Arch. Geflugelkd. 2007, 71, 162–168. [Google Scholar]
- Balta, I.; Butucel, E.; Stef, L.; Pet, I.; Gradisteanu-Pircalabioru, G.; Chifiriuc, C.; Gundogdu, O.; McCleery, D.; Corcionivoschi, N. Anti-Campylobacter probiotics: Latest mechanistic insights. Foodborne Pathog. Dis. 2022, 19, 693–703. [Google Scholar] [CrossRef]
- Bao, C.; Zhang, W.; Wang, J.; Liu, Y.; Cao, H.; Li, F.; Liu, S.; Shang, Z.; Cao, Y.; Dong, B. The Effects of Dietary Bacillus amyloliquefaciens TL106 Supplementation, as an Alternative to Antibiotics, on Growth Performance, Intestinal Immunity, Epithelial Barrier Integrity, and Intestinal Microbiota in Broilers. Animals 2022, 12, 3085. [Google Scholar] [CrossRef]
- Farhat-Khemakhem, A.; Blibech, M.; Boukhris, I.; Makni, M.; Chouayekh, H. Assessment of the potential of the multi-enzyme producer Bacillus amyloliquefaciens US573 as alternative feed additive. J. Sci. Food Agric. 2018, 98, 1208–1215. [Google Scholar] [CrossRef] [PubMed]
- Zhen, W.; Shao, Y.; Gong, X.; Wu, Y.; Geng, Y.; Wang, Z.; Guo, Y. Effect of dietary Bacillus coagulans supplementation on growth performance and immune responses of broiler chickens challenged by Salmonella enteritidis. Poult. Sci. 2018, 97, 2654–2666. [Google Scholar] [CrossRef]
- Sugiarto, H.; Yu, P.-L. Avian antimicrobial peptides: The defense role of β-defensins. Biochem. Biophys. Res. Commun. 2004, 323, 721–727. [Google Scholar] [CrossRef]
- Mohammed, E.S.; Igarashi, Y.; Isobe, N.; Yoshimura, Y. Effects of Probiotics on the Expression and Localization of Avian β-defensins in the Proventriculus of Broiler Chicks. J. Poult. Sci. 2015, 52, 57–67. [Google Scholar] [CrossRef]
- Rima, M.; Rima, M.; Fajloun, Z.; Sabatier, J.-M.; Bechinger, B.; Naas, T. Antimicrobial peptides: A potent alternative to antibiotics. Antibiotics 2021, 10, 1095. [Google Scholar] [CrossRef]
- Linden, S.; Sutton, P.; Karlsson, N.; Korolik, V.; McGuckin, M. Mucins in the mucosal barrier to infection. Mucosal Immunol. 2008, 1, 183–197. [Google Scholar] [CrossRef]
- Abd El-Hamid, M.I.; Ibrahim, D.; Hamed, R.I.; Nossieur, H.H.; Elbanna, M.H.; Baz, H.; Abd-Allah, E.M.; El Oksh, A.S.; Ibrahim, G.A.; Khalifa, E. Modulatory impacts of multi-strain probiotics on rabbits’ growth, nutrient transporters, tight junctions and immune system to fight against Listeria monocytogenes infection. Animals 2022, 12, 2082. [Google Scholar] [CrossRef]
- de LeBlanc, A.d.M.; Dogi, C.A.; Galdeano, C.M.; Carmuega, E.; Weill, R.; Perdigón, G. Effect of the administration of a fermented milk containing Lactobacillus casei DN-114001 on intestinal microbiota and gut associated immune cells of nursing mice and after weaning until immune maturity. BMC Immunol. 2008, 9, 27. [Google Scholar] [CrossRef] [PubMed]
- Aliakbarpour, H.; Chamani, M.; Rahimi, G.; Sadeghi, A.; Qujeq, D. The Bacillus subtilis and lactic acid bacteria probiotics influences intestinal mucin gene expression, histomorphology and growth performance in broilers. Asian-Australas. J. Anim. Sci. 2012, 25, 1285. [Google Scholar] [CrossRef]
- Chen, J.; Tellez, G.; Richards, J.D.; Escobar, J. Identification of potential biomarkers for gut barrier failure in broiler chickens. Front. Vet. Sci. 2015, 2, 14. [Google Scholar] [CrossRef]
- Hashem, Y.M.; Abd El-Hamid, M.I.; Awad, N.F.; Ibrahim, D.; Elshater, N.S.; El-Malt, R.M.; Hassan, W.H.; Abo-Shama, U.H.; Nassan, M.A.; El-Bahy, S.M. Insights into growth-promoting, anti-inflammatory, immunostimulant, and antibacterial activities of Toldin CRD as a novel phytobiotic in broiler chickens experimentally infected with Mycoplasma gallisepticum. Poult. Sci. 2022, 101, 102154. [Google Scholar] [CrossRef] [PubMed]
- Awad, N.F.; Abd El-Hamid, M.I.; Hashem, Y.; Erfan, A.; Abdelrahman, B.; Mahmoud, H.I. Impact of single and mixed infections with Escherichia coli and Mycoplasma gallisepticum on Newcastle disease virus vaccine performance in broiler chickens: An in vivo perspective. J. Appl. Microbiol. 2019, 127, 396–405. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).