
Using an Automated Operant Conditioning Procedure to Test Colour Discrimination in Two Juvenile Piranhas, Pygocentrus nattereri: A Lesson on Failures and Pitfalls and How to Avoid Them


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Subjects
2.2. Preliminary Assessment of Colour Preference
2.2.1. Stimuli and Apparatus
2.2.2. Procedure
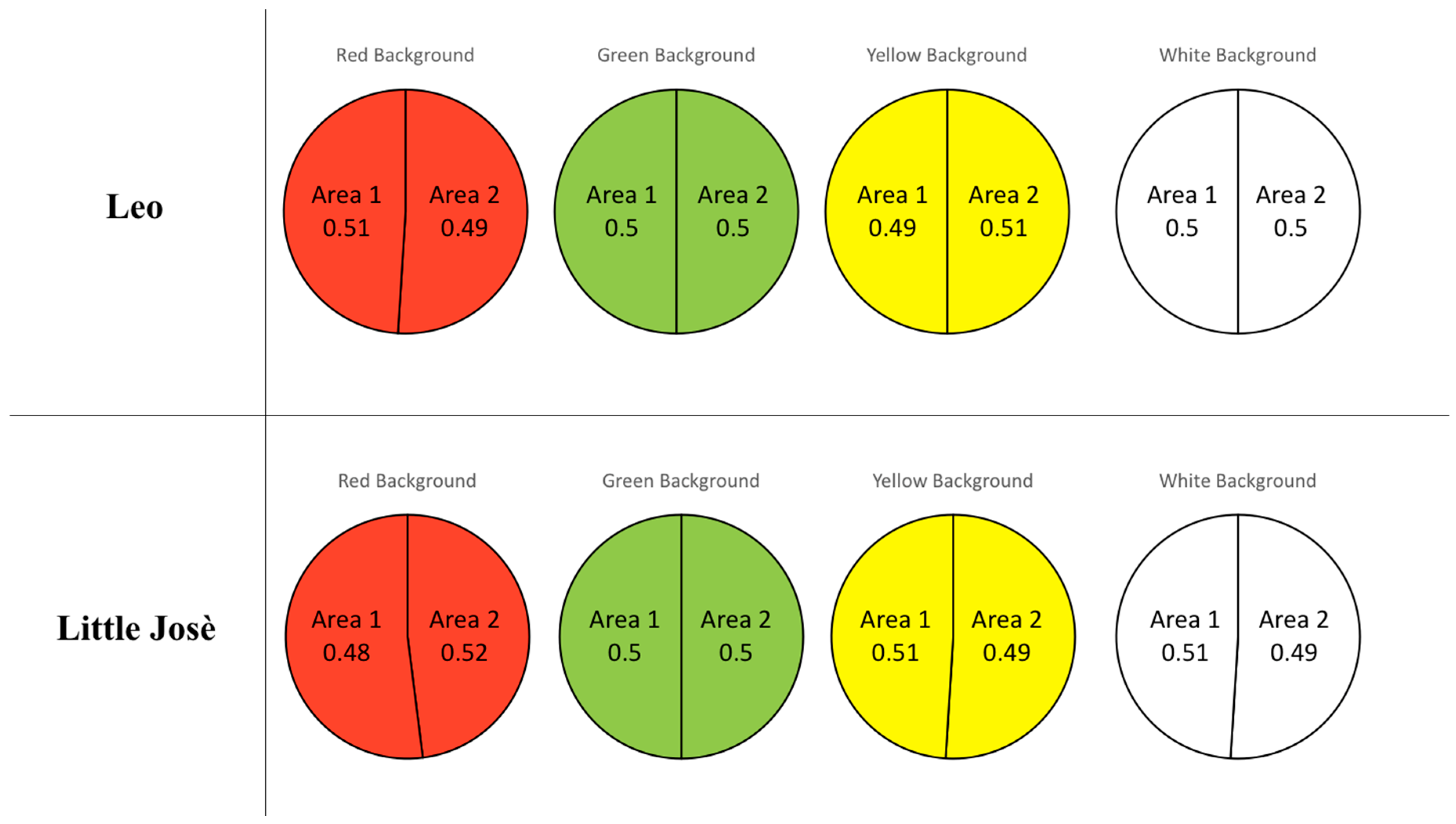
2.2.3. Data Analysis
2.3. Experiment 1 and 2: Response Learning with Coloured Stimuli
2.3.1. Stimuli and Apparatus
2.3.2. Procedure
2.4. Experiment 3: Response Learning (Achromatic Stimuli) + Place Learning Task
2.5. Data Analysis for Experiment 1, 2, and 3.
3. Results and Discussion
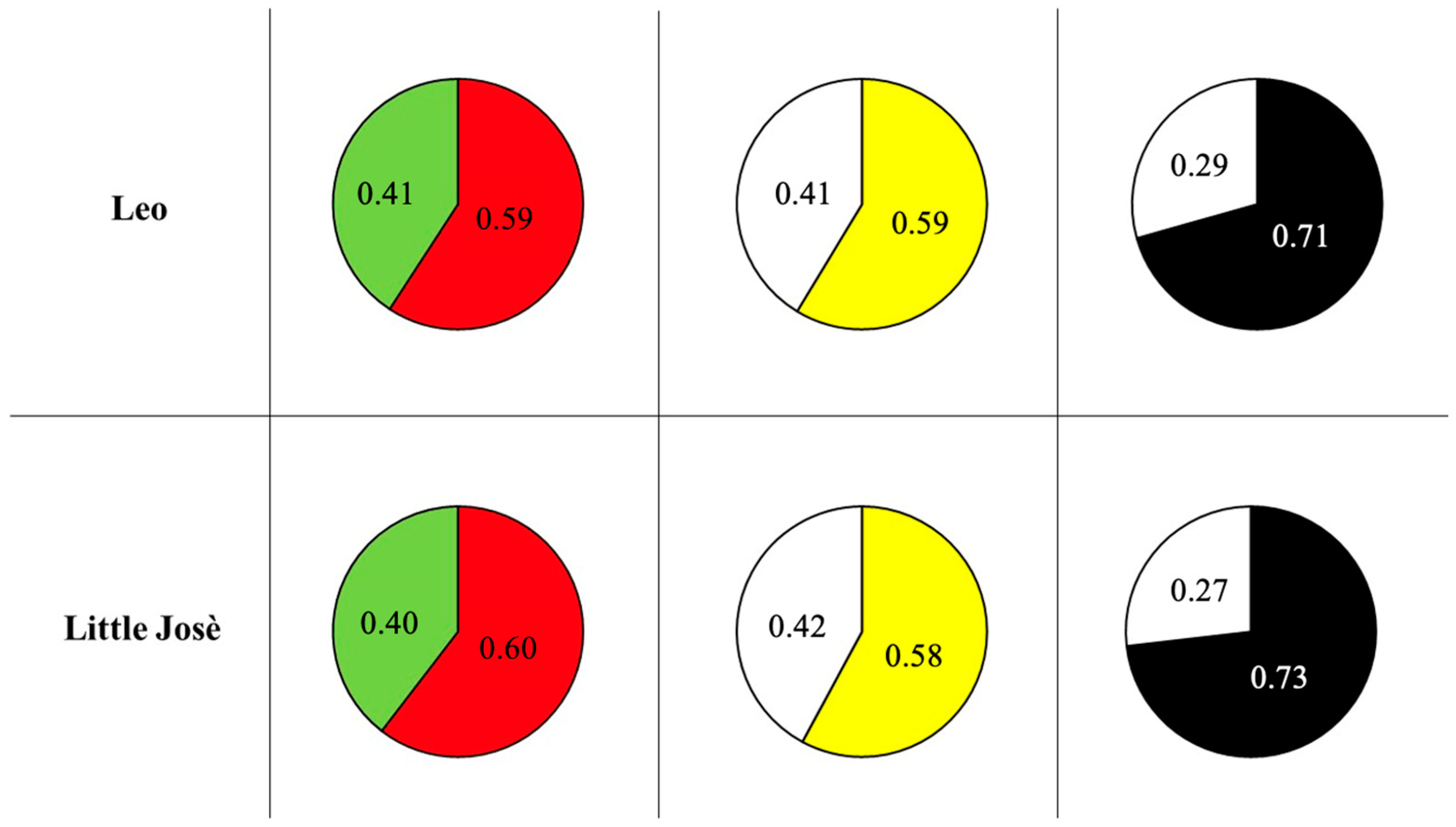
3.1. Preliminary Assessment of Colour Preference
3.2. Response Learning with Coloured Stimuli
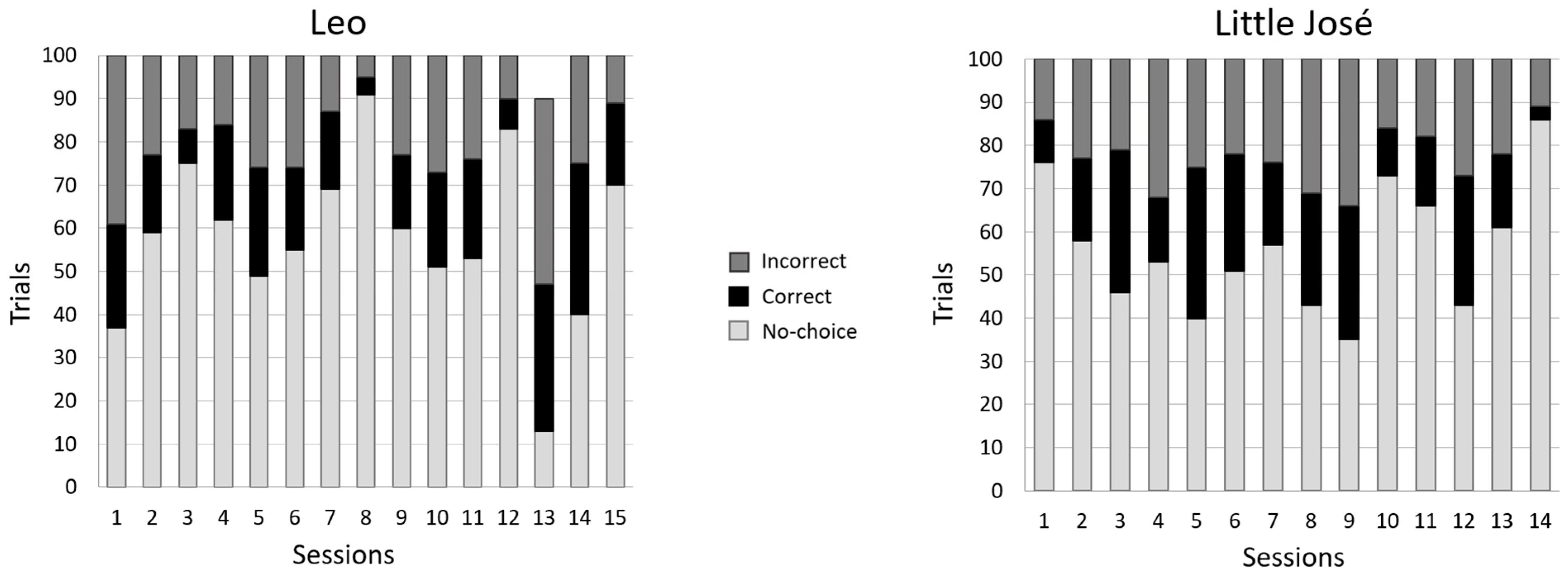
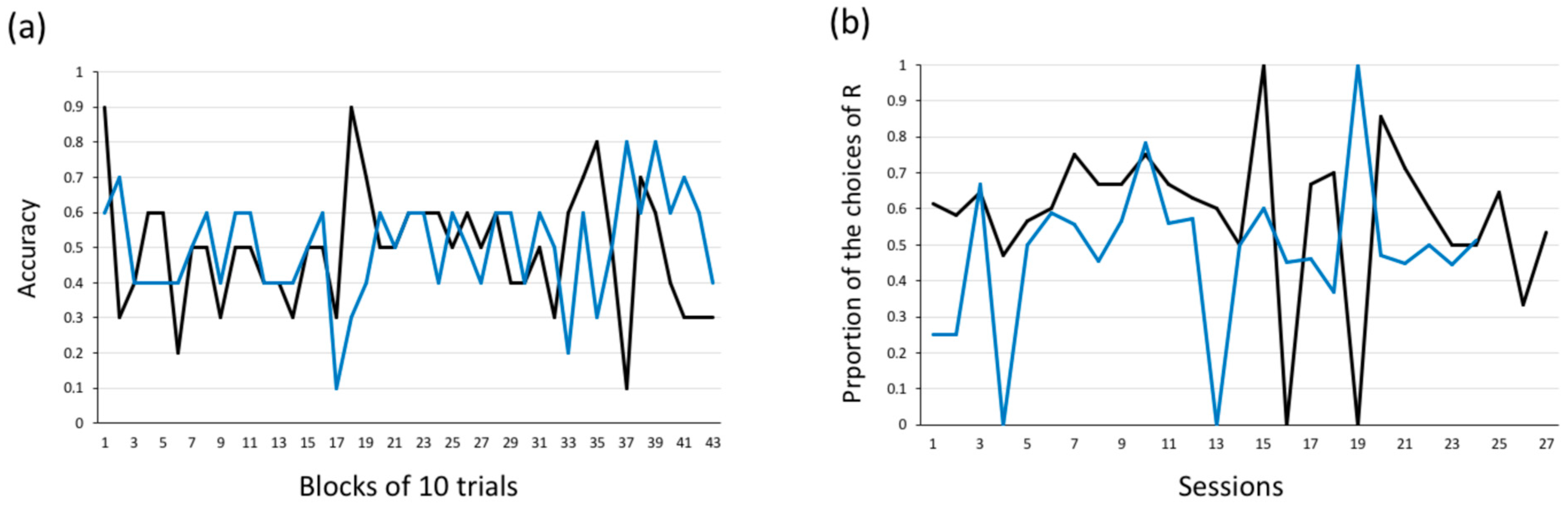
3.2.1. Experiment 1 (Red Vs. Green)
3.2.2. Experiment 2 (Yellow vs. White)
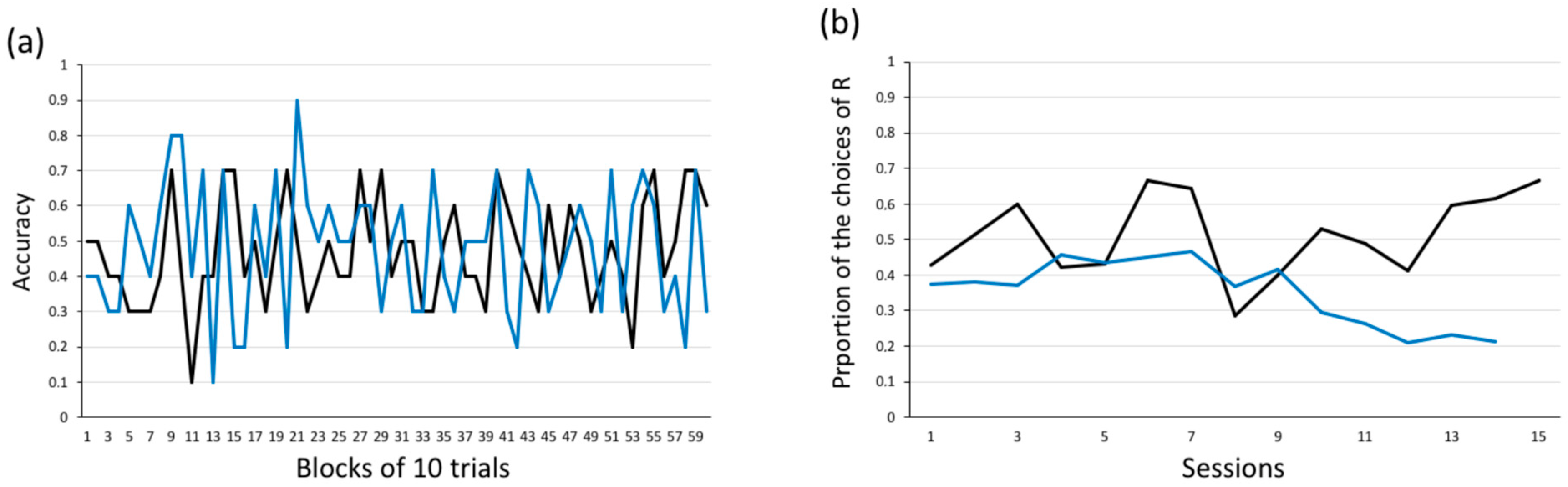
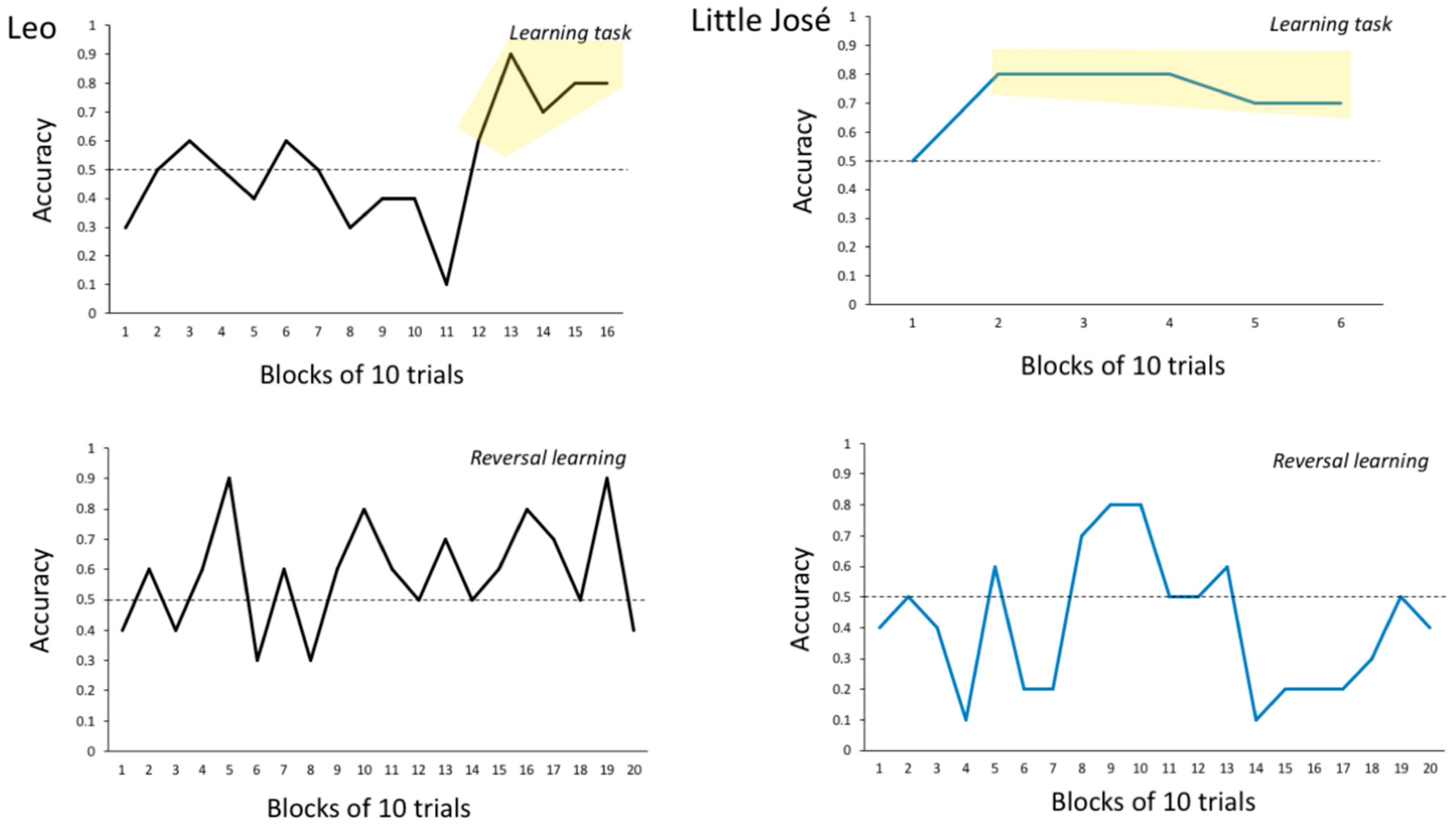
3.2.3. Experiment 3: Response Learning (Achromatic Stimuli) + Place Learning Task
4. Conclusions and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, C.; Laland, K.; Krause, J. Fish Cognition and Behavior; John Wiley & Sons: Hoboken, NJ, USA, 2011. [Google Scholar]
- Bshary, R.; Wickler, W.; Fricke, H. Fish cognition: A primate’s eye view. Anim. Cogn. 2002, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Salena, M.G.; Turko, A.J.; Singh, A.; Pathak, A.; Hughes, E.; Brown, C.; Balshine, S. Understanding fish cognition: A review and appraisal of current practices. Anim. Cogn. 2021, 24, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Buechel, S.D.; Boussard, A.; Kotrschal, A.; van der Bijl, W.; Kolm, N. Brain size affects performance in a reversal-learning test. Proc. R. Soc. B Biol. Sci. 2018, 285, 20172031. [Google Scholar] [CrossRef] [PubMed]
- Daniel, M.M.; Schluessel, V. Serial reversal learning in freshwater stingrays (Potamotrygon motoro). Anim. Cogn. 2020, 23, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Miletto Petrazzini, M.E.; Agrillo, C.; Izard, V.; Bisazza, A. Do humans (Homo sapiens) and fish (Pterophyllum scalare) make similar numerosity judgments? J. Comp. Psychol. 2016, 130, 380. [Google Scholar] [CrossRef]
- Miletto Petrazzini, M.E.; Pecunioso, A.; Dadda, M.; Agrillo, C. Searching for the critical p of Macphail’s null hypothesis: The contribution of numerical abilities of fish. Front. Psychol. 2020, 11, 55. [Google Scholar] [CrossRef]
- Bisazza, A.; Butterworth, B.; Piffer, L.; Bahrami, B.; Miletto Petrazzini, M.E.; Agrillo, C. Collective enhancement of numerical acuity by meritocratic leadership in fish. Sci. Rep. 2014, 4, 4560. [Google Scholar] [CrossRef]
- Gómez-Laplaza, L.M.; Gerlai, R. Spontaneous discrimination of small quantities: Shoaling preferences in angelfish (Pterophyllum scalare). Anim. Cogn. 2011, 14, 565–574. [Google Scholar] [CrossRef]
- Potrich, D.; Zanon, M.; Vallortigara, G. Archerfish number discrimination. eLife 2022, 11, e74057. [Google Scholar] [CrossRef]
- Lee, S.A.; Vallortigara, G.; Flore, M.; Spelke, E.S.; Sovrano, V.A. Navigation by environmental geometry: The use of zebrafish as a model. J. Exp. Biol. 2013, 216, 3693–3699. [Google Scholar] [CrossRef]
- Braithwaite, V.A.; De Perera, T.B. Short-range orientation in fish: How fish map space. Mar. Freshw. Behav. Physiol. 2006, 39, 37–47. [Google Scholar] [CrossRef]
- Kohda, M.; Bshary, R.; Kubo, N.; Awata, S.; Sowersby, W.; Kawasaka, K.; Kobayashi, T.; Sogawa, S. Cleaner fish recognize self in a mirror via self-face recognition like humans. Proc. Nat. Acad. Sci. USA 2023, 120, e2208420120. [Google Scholar] [CrossRef] [PubMed]
- Mair, A.; Dadda, M.; Kitaoka, A.; Agrillo, C. Illu-Shoal choice: An exploration of different means for enrichment of captive zebrafish. Animals 2023, 13, 2640. [Google Scholar] [CrossRef] [PubMed]
- Meshalkina, D.A.; Kizlyk, M.N.; Kysil, E.V.; Collier, A.D.; Echevarria, D.J.; Abreu, M.S.; Barcellos, L.J.G.; Song, C.; Kalueff, A.V. Understanding zebrafish cognition. Behav. Proc. 2017, 141, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.; Laland, K.N. Social learning of a novel avoidance task in the guppy: Conformity and social release. Anim. Behav. 2002, 64, 41–47. [Google Scholar] [CrossRef]
- Lucon-Xiccato, T.; Bisazza, A.; Bertolucci, C. Guppies show sex and individual differences in the ability to inhibit behaviour. Anim. Cogn. 2020, 23, 535–543. [Google Scholar] [CrossRef]
- Gómez-Laplaza, L.M.; Gerlai, R. Can angelfish (Pterophyllum scalare) count? Discrimination between different shoal sizes follows Weber’s law. Anim. Cogn. 2011, 14, 1–9. [Google Scholar] [CrossRef]
- Gómez-Laplaza, L.M.; Gerlai, R. Angelfish (Pterophyllum scalare) discriminate between small quantities: A role of memory. J. Comp. Psychol. 2015, 129, 78. [Google Scholar] [CrossRef]
- Wegman, J.J.; Morrison, E.; Wilcox, K.T.; DeLong, C.M. Visual perception of photographs of rotated 3d objects in goldfish (Carassius auratus). Animals 2022, 12, 1797. [Google Scholar] [CrossRef]
- Otero Coronel, S.; Martorell, N.; Beron de Astrada, M.; Medan, V. Stimulus contrast information modulates sensorimotor decision making in goldfish. Front. Neural Circuits 2020, 14, 23. [Google Scholar] [CrossRef]
- Sibeaux, A.; Karlsson, C.; Newport, C.; Burt de Perera, T. Distance estimation in the goldfish (Carassius auratus). Proc. R. Soc. Lond. Ser. B 2022, 289, 20221220. [Google Scholar] [CrossRef] [PubMed]
- Triki, Z.; Bshary, R. Sex differences in the cognitive abilities of a sex-changing fish species Labroides dimidiatus. R. Soc. Open Sci. 2021, 8, 210239. [Google Scholar] [CrossRef] [PubMed]
- Aellen, M.; Burkart, J.M.; Bshary, R. No evidence for general intelligence in a fish. Ethology 2022, 128, 424–436. [Google Scholar] [CrossRef]
- Raick, X.; Huby, A.; Kurchevski, G.; Godinho, A.L.; Parmentier, É. Yellow-eyed piranhas produce louder sounds than red-eyed piranhas in an invasive population of Serrasalmus marginatus. J. Fish Biol. 2020, 97, 1676–1680. [Google Scholar] [CrossRef] [PubMed]
- Uetanabaro, M.; Wang, T.; Abe, A.S. Breeding behaviour of the red-bellied piranha, Pygocentrus nattereri, in nature. Environ. Biol. Fishes 1993, 38, 369–371. [Google Scholar] [CrossRef]
- Velasco-Hogan, A.; Meyers, M.A. Bite force mechanics and allometry of piranha (Serrasalmidae). J. Mech. Behav. Biomed. Mater. 2021, 115, 104296. [Google Scholar] [CrossRef]
- Skinner, B.F. Selection by consequences. Science 1981, 213, 501–504. [Google Scholar] [CrossRef]
- Cantlon, J.F.; Brannon, E.M. Basic math in monkeys and college students. PLoS Biol. 2007, 5, e328. [Google Scholar] [CrossRef]
- Perdue, B.M.; Beran, M.J.; Washburn, D.A. A computerized testing system for primates: Cognition, welfare, and the Rumbaughx. Behav. Proc. 2018, 156, 37–50. [Google Scholar] [CrossRef]
- Herbranson, W.T. Pigeons, humans, and the Monty Hall dilemma. Curr. Dir. Psychol. Sci. 2012, 21, 297–301. [Google Scholar] [CrossRef]
- Nakamura, N.; Watanabe, S.; Fujita, K. Pigeons perceive the Ebbinghaus-Titchener circles as an assimilation illusion. J. Exp. Psychol. Anim. Behav. Proc. 2008, 34, 375. [Google Scholar] [CrossRef] [PubMed]
- Gatto, E.; Testolin, A.; Bisazza, A.; Zorzi, M.; Lucon-Xiccato, T. Poor numerical performance of guppies tested in a Skinner box. Sci. Rep. 2020, 10, 16724. [Google Scholar] [CrossRef] [PubMed]
- Gatto, E.; Lucon-Xiccato, T.; Bisazza, A.; Manabe, K.; Dadda, M. The devil is in the detail: Zebrafish learn to discriminate visual stimuli only if salient. Behav. Proc. 2020, 179, 104215. [Google Scholar] [CrossRef] [PubMed]
- Vonk, J.; Torgerson-White, L.; McGuire, M.; Thueme, M.; Thomas, J.; Beran, M.J. Quantity estimation and comparison in western lowland gorillas (Gorilla gorilla gorilla). Anim. Cogn. 2014, 17, 755–765. [Google Scholar] [CrossRef]
- Vonk, J.; Beran, M.J. Bears ‘count’ too: Quantity estimation and comparison in black bears, Ursus americanus. Anim. Behav. 2012, 84, 231–238. [Google Scholar] [CrossRef]
- Perdue, B.M.; Talbot, C.F.; Stone, A.M.; Beran, M.J. Putting the elephant back in the herd: Elephant relative quantity judgments match those of other species. Anim. Cogn. 2012, 15, 955–961. [Google Scholar] [CrossRef]
- Pepperberg, I.M. Grey parrot numerical competence: A review. Anim.Cogn. 2006, 9, 377–391. [Google Scholar] [CrossRef]
- Bisazza, A.; Tagliapietra, C.; Bertolucci, C.; Foà, A.; Agrillo, C. Non-visual numerical discrimination in a blind cavefish (Phreatichthys andruzzii). J. Exp. Biol. 2014, 217, 1902–1909. [Google Scholar] [CrossRef]
- Fuss, T.; Schluessel, V. The Ebbinghaus illusion in the gray bamboo shark (Chiloscyllium griseum) in comparison to the teleost damselfish (Chromis chromis). Zoology 2017, 123, 16–29. [Google Scholar] [CrossRef]
- Brown, J.L.; Shively, F.D.; LaMotte, R.H.; Sechzer, J.A. Color discrimination in the cat. J. Comp. Physiol. Psychol. 1973, 84, 534. [Google Scholar] [CrossRef]
- Miletto Petrazzini, M.E.; Pecunioso, A.; Dadda, M.; Agrillo, C. The impact of brain lateralization and anxiety-like behaviour in an extensive operant conditioning task in zebrafish (Danio rerio). Symmetry 2019, 11, 1395. [Google Scholar] [CrossRef]
- Savage, A.; Dronzek, L.A.; Snowdon, C.T. Color discrimination by the cotton-top tamarin (Saguinus oedipus oedipus) and its relation to fruit coloration. Folia Primatol. 1987, 49, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Goodman, J. Place vs. response learning: History, controversy, and neurobiology. Front. Behav. Neurosci. 2021, 14, 598570. [Google Scholar] [CrossRef] [PubMed]
- Lucon-Xiccato, T.; Bisazza, A. Discrimination reversal learning reveals greater female behavioural flexibility in guppies. Biol. Lett. 2014, 10, 20140206. [Google Scholar] [CrossRef]
- Agrillo, C.; Miletto Petrazzini, M.E.; Tagliapietra, C.; Bisazza, A. Inter-specific differences in numerical abilities among teleost fish. Front. Psychol. 2012, 3, 483. [Google Scholar] [CrossRef] [PubMed]
- Agrillo, C.; Bisazza, A. Understanding the origin of number sense: A review of fish studies. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20160511. [Google Scholar] [CrossRef]
- Earl, B. Humans, fish, spiders and bees inherited working memory and attention from their last common ancestor. Front. Psychol. 2023, 13, 937712. [Google Scholar] [CrossRef]
- Maximino, C.; Marques de Brito, T.; Dias, C.A.; Gouveia, A., Jr.; Morato, S. Scototaxis as anxiety-like behavior in fish. Nat. Protoc. 2010, 5, 209–216. [Google Scholar] [CrossRef]
- Miletto Petrazzini, M.E.; Gambaretto, L.; Dadda, M.; Brennan, C.; Agrillo, C. Are cerebral and behavioural lateralization related to anxiety-like traits in the animal model zebrafish (Danio rerio)? Laterality 2021, 26, 144–162. [Google Scholar] [CrossRef]
- Miletto Petrazzini, M.E.; Pecunioso, A.; Dadda, M.; Agrillo, C. Does brain lateralization affect the performance in binary choice tasks? A study in the animal model Danio rerio. Symmetry 2020, 12, 1294. [Google Scholar] [CrossRef]
- Agrillo, C.; Bisazza, A. Spontaneous versus trained numerical abilities. A comparison between the two main tools to study numerical competence in non-human animals. J. Neurosci. Methods 2014, 234, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Santacà, M.; Agrillo, C. Two halves are less than the whole: Evidence of a length bisection bias in fish (Poecilia reticulata). PLoS ONE 2020, 15, e0233157. [Google Scholar] [CrossRef] [PubMed]
- Bisazza, A.; Santacà, M. Zebrafish excel in number discrimination under an operant conditioning paradigm. Anim. Cogn. 2022, 25, 917–933. [Google Scholar] [CrossRef] [PubMed]
- Maximino, C.; Marques, T.; Dias, F.; Cortes, F.V.; Taccolini, I.B.; Pereira, P.M.; Colmanetti, R.; Gazolla, R.A.; Tavares, R.I.; Rodrigues, S.T.K.; et al. A comparative analysis of the preference for dark environments in five teleosts. Int. J. Comp. Psychol. 2007, 20, 351–367. [Google Scholar] [CrossRef]
- Jozet-Alves, C.; Viblanc, V.A.; Romagny, S.; Dacher, M.; Healy, S.D.; Dickel, L. Visual lateralization is task and age dependent in cuttlefish, Sepia officinalis. Anim. Behav. 2012, 83, 1313–1318. [Google Scholar] [CrossRef]
- O’Shea-Wheller, T.A. Honeybees show a context-dependent rightward bias. Biol. Lett. 2009, 15, 20180877. [Google Scholar] [CrossRef]
- Spiezio, C.; Pugassi, M.; Agrillo, C.; Regaiolli, B. Color preference and manual laterality in the emperor tamarin (Saguinus imperator). Am. J. Primatol. 2022, 84, e23375. [Google Scholar] [CrossRef]
- Agrillo, C.; Parrish, A.E.; Beran, M.J. Do primates see the solitaire illusion differently? A comparative assessment of humans (Homo sapiens), chimpanzees (Pan troglodytes), rhesus monkeys (Macaca mulatta), and capuchin monkeys (Cebus apella). J. Comp. Psychol. 2014, 128, 402–413. [Google Scholar] [CrossRef]
- Knudson, D.V.; Lindsey, C. Type I and Type II errors in correlations of various sample sizes. Comp. Psychol. 2014, 3, 03-CP. [Google Scholar] [CrossRef]
- Kanai, R.; Rees, G. The structural basis of inter-individual differences in human behaviour and cognition. Nat. Rev. Neurosci. 2011, 12, 231–242. [Google Scholar] [CrossRef]
- Lucon-Xiccato, T.; Manabe, K.; Bisazza, A. Guppies learn faster to discriminate between red and yellow than between two shapes. Ethology 2019, 125, 82–91. [Google Scholar] [CrossRef]
- Gatto, E.; Santacà, M.; Verza, I.; Dadda, M.; Bisazza, A. Automated operant conditioning devices for fish. Do they work? Animals 2021, 11, 1397. [Google Scholar] [CrossRef] [PubMed]
- Gatto, E.; Lucon-Xiccato, T.; Savaşçı, B.B.; Dadda, M.; Bisazza, A. Experimental setting affects the performance of guppies in a numerical discrimination task. Anim. Cogn. 2017, 20, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Escobar-Camacho, D.; Carleton, K.L.; Narain, D.W.; Pierotti, M.E. Visual pigment evolution in Characiformes: The dynamic interplay of teleost whole-genome duplication, surviving opsins and spectral tuning. Mol. Ecol. 2020, 29, 2234–2253. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, B.F.; Aeschliman, B.; Pierce, P. Studies in spatial learning. VIII. Place performance and the acquisition of place dispositions. J. Comp. Phys. Psychol. 1950, 43, 73–85. [Google Scholar] [CrossRef]
- Hicks, L.H. Effects of overtraining on acquisition and reversal of place and response learning. Psychol. Rep. 1964, 15, 459–462. [Google Scholar] [CrossRef]
- Packard, M.G.; McGaugh, J.L. Double dissociation of fornix and caudate nucleus lesions on acquisition of two water maze tasks: Further evidence for multiple memory systems. Behav. Neurosci. 1992, 106, 439–446. [Google Scholar] [CrossRef]
- Packard, M.G. Glutamate infused posttraining into the hippocampus or caudate-putamen differentially strengthens place and response learning. Proc. Natl. Acad. Sci. USA 1999, 96, 12881–12886. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agrillo, C.; Pecunioso, A. Using an Automated Operant Conditioning Procedure to Test Colour Discrimination in Two Juvenile Piranhas, Pygocentrus nattereri: A Lesson on Failures and Pitfalls and How to Avoid Them. Animals 2024, 14, 3187. https://doi.org/10.3390/ani14223187
Agrillo C, Pecunioso A. Using an Automated Operant Conditioning Procedure to Test Colour Discrimination in Two Juvenile Piranhas, Pygocentrus nattereri: A Lesson on Failures and Pitfalls and How to Avoid Them. Animals. 2024; 14(22):3187. https://doi.org/10.3390/ani14223187
Chicago/Turabian StyleAgrillo, Christian, and Alessandra Pecunioso. 2024. "Using an Automated Operant Conditioning Procedure to Test Colour Discrimination in Two Juvenile Piranhas, Pygocentrus nattereri: A Lesson on Failures and Pitfalls and How to Avoid Them" Animals 14, no. 22: 3187. https://doi.org/10.3390/ani14223187
APA StyleAgrillo, C., & Pecunioso, A. (2024). Using an Automated Operant Conditioning Procedure to Test Colour Discrimination in Two Juvenile Piranhas, Pygocentrus nattereri: A Lesson on Failures and Pitfalls and How to Avoid Them. Animals, 14(22), 3187. https://doi.org/10.3390/ani14223187