
Effect of Anacardium occidentale Leaf Powder on Growth Performance, Diarrhea Incidence, Blood Biochemistry, and Intestinal Traits in Weaned Piglets

 , , and
, , and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Animal Management
2.2. Growth Performance and Diarrhea Incidence Measurements
2.3. Sample Collection
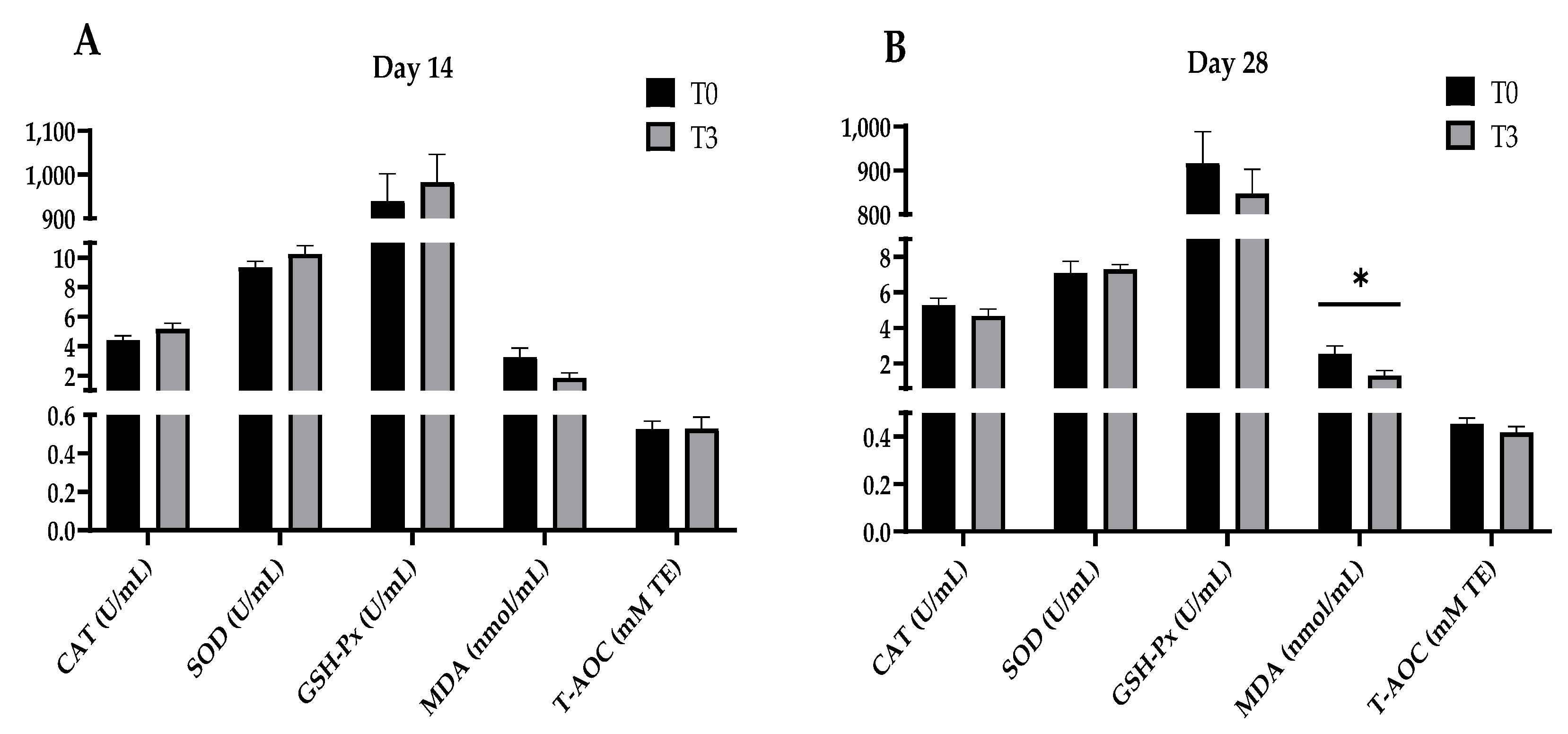
2.4. Antioxidant Indexes in Plasma
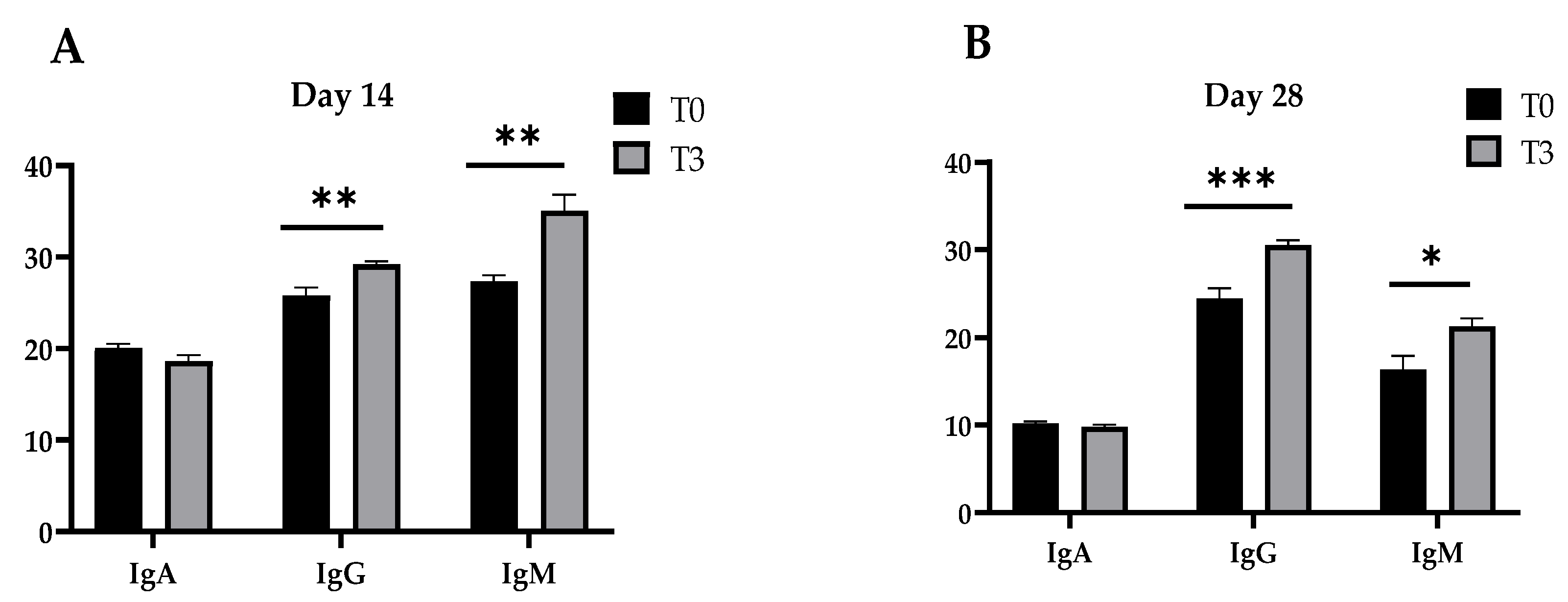
2.5. Immunoglobulin Measurement in Plasma
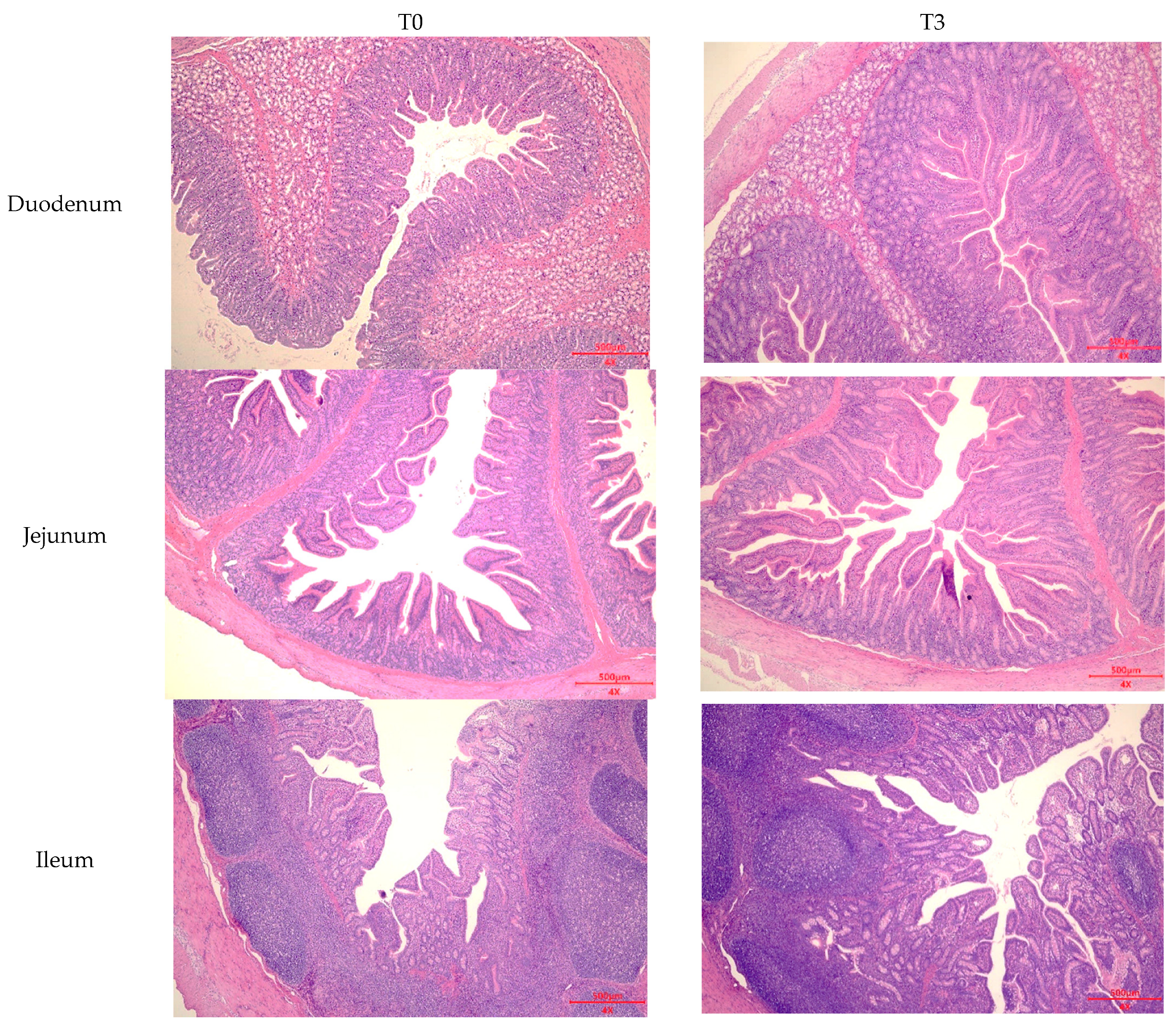
2.6. Intestinal Morphology
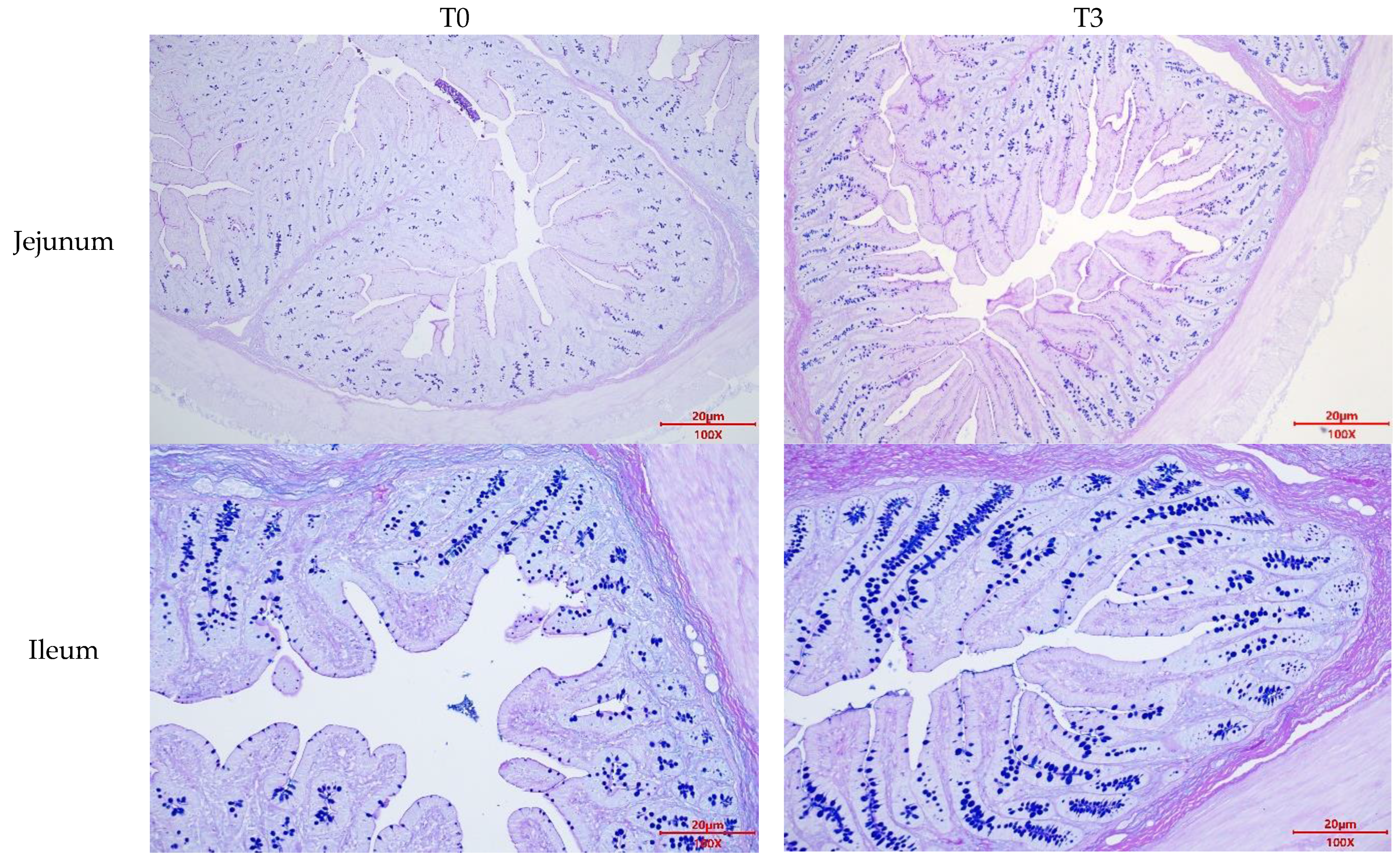
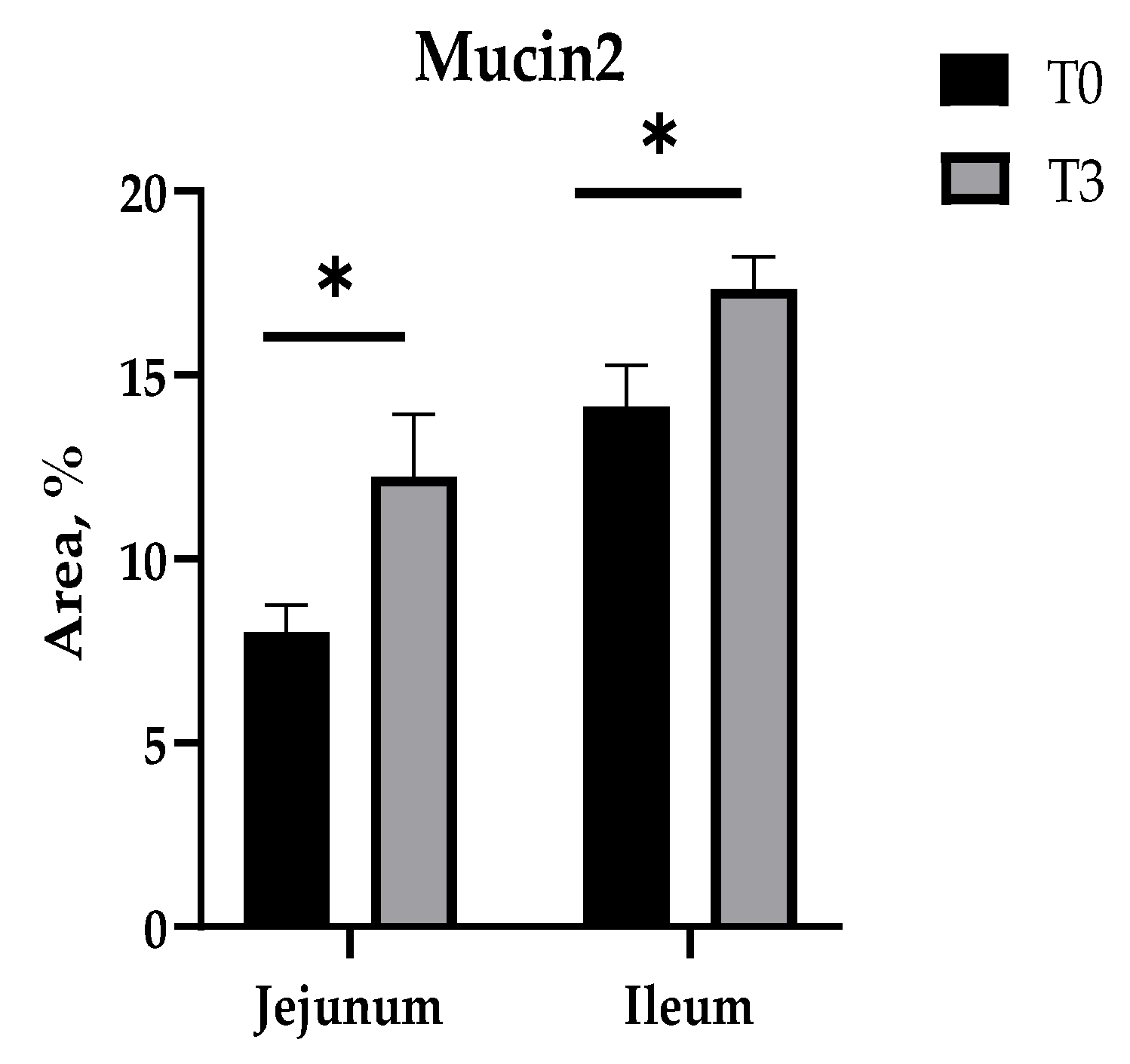
2.7. Mucin2 Area Determination
2.8. RNA Extraction and Quantitative Real-Time PCR (qPCR)
2.9. Analysis of Microbiota Diversity
2.10. Short-Chain Fatty Acids (SCFA) Concentration
2.11. Statistical Analysis
3. Results
3.1. Growth Performance and Diarrhea Incidence
3.2. Antioxidant Status
3.3. Immunoglobulin Concentration in Plasma
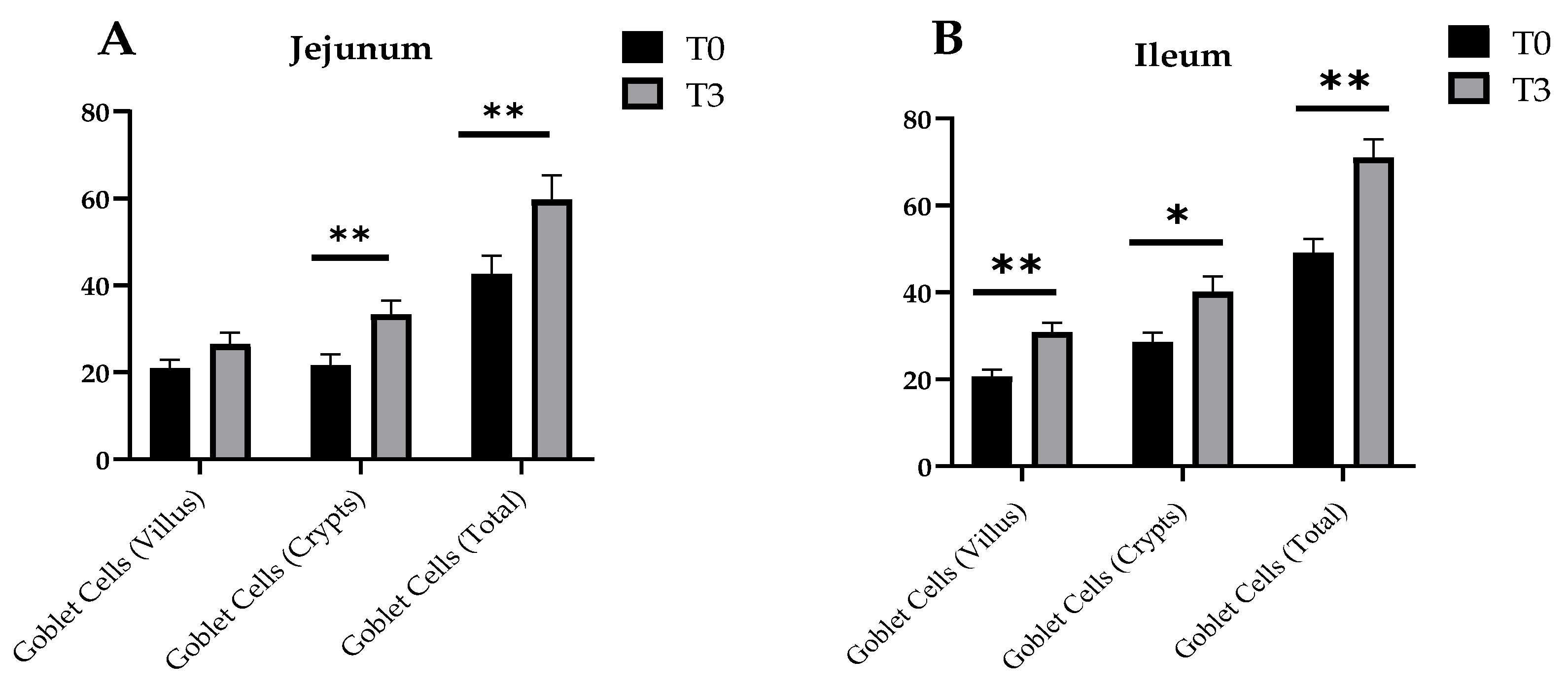
3.4. Intestinal Morphology and Goblet Cell Number
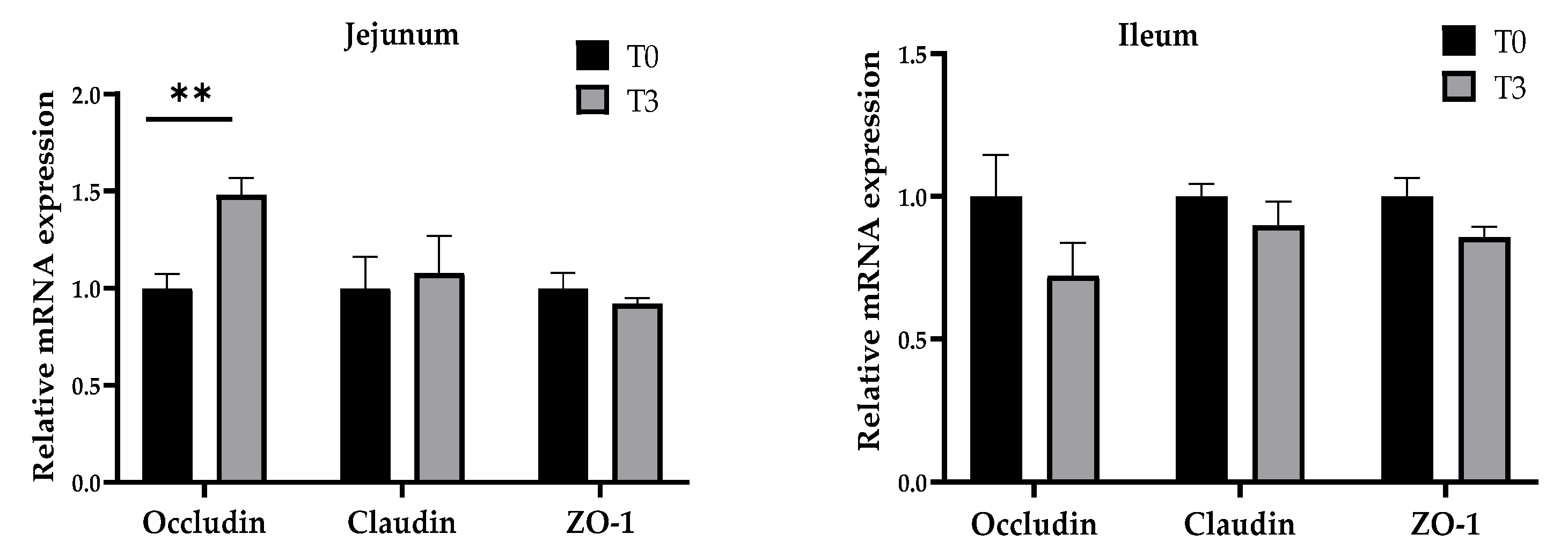
3.5. Relative Gene Expression
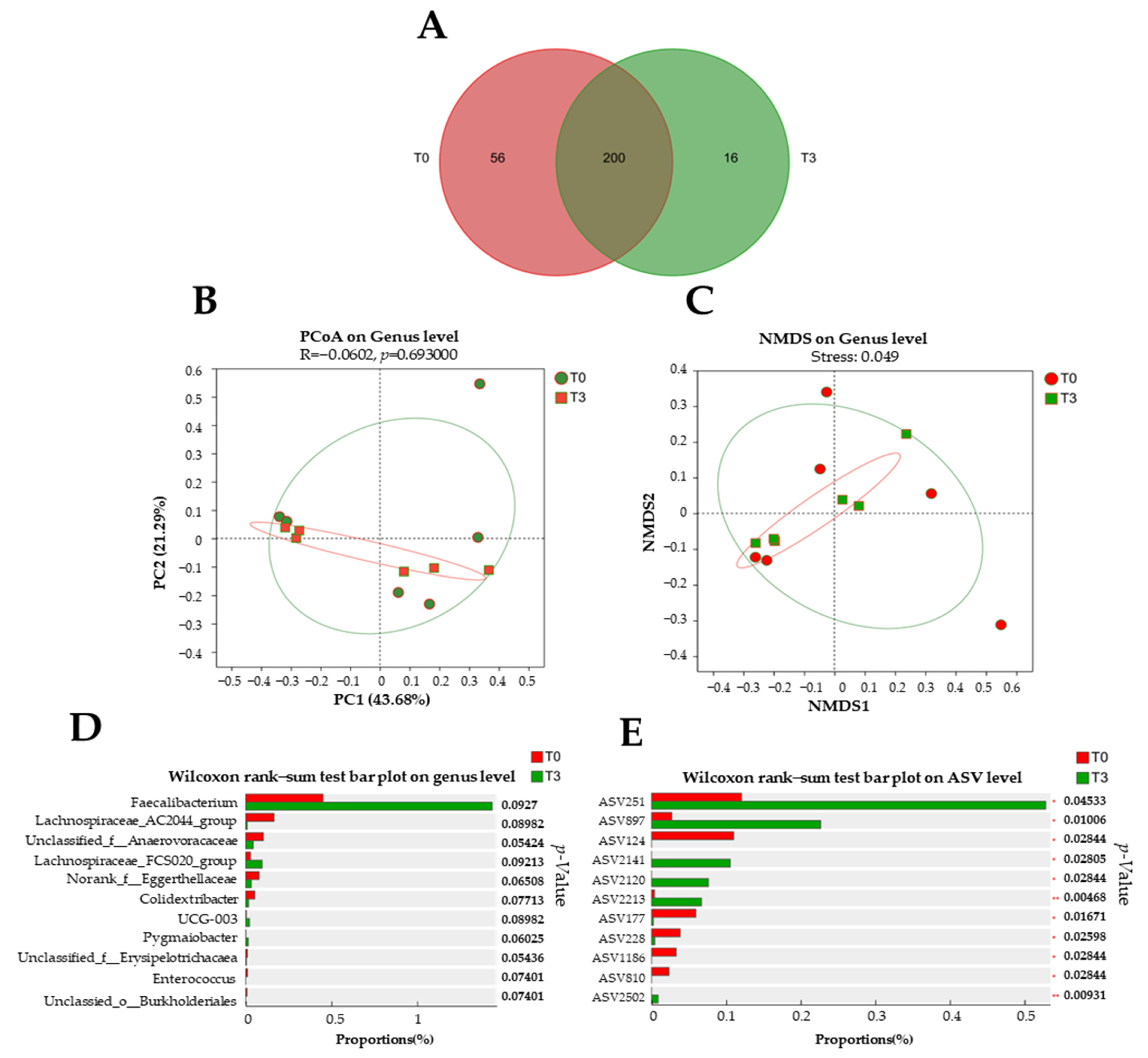
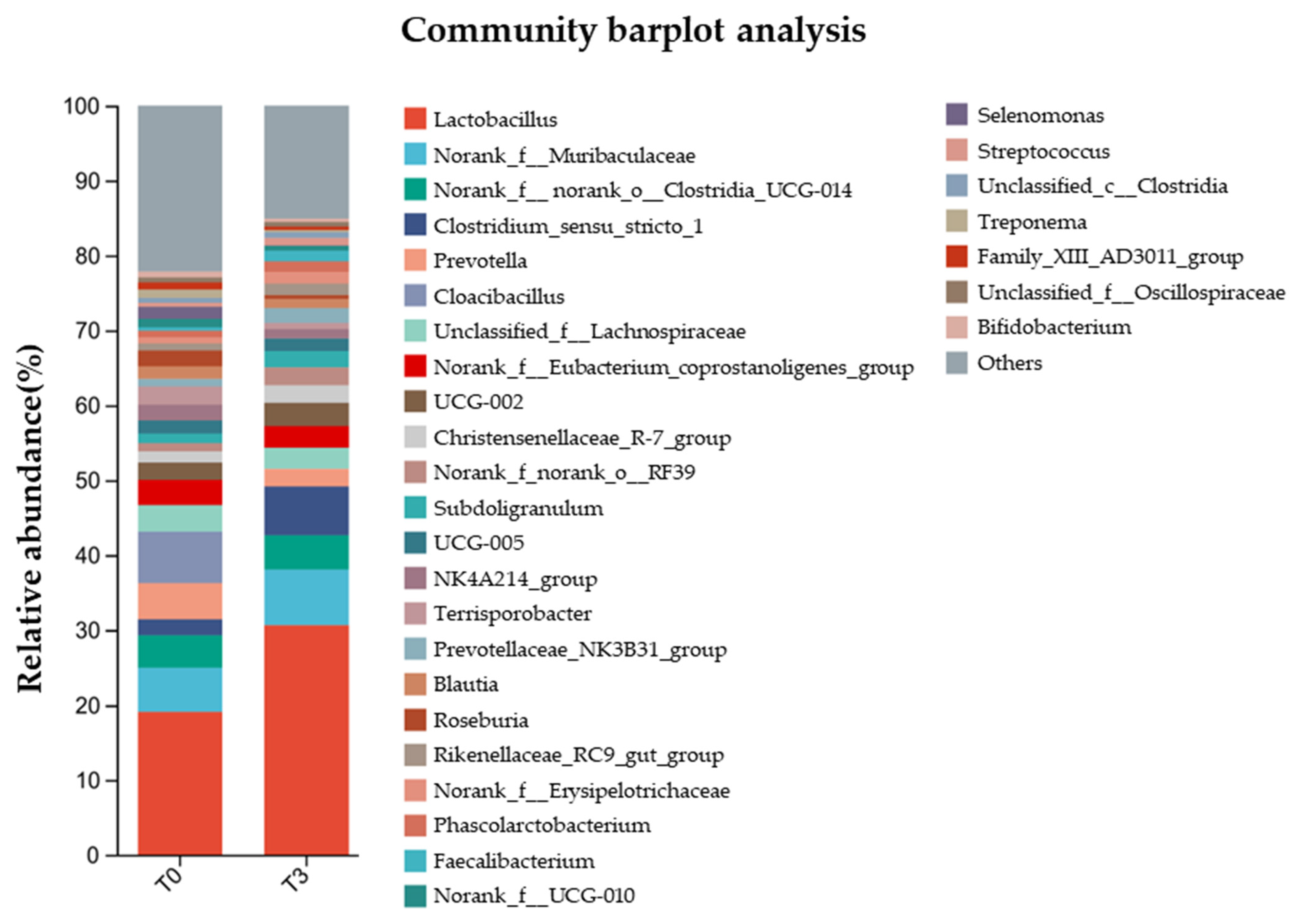
3.6. Microbiota Diversity
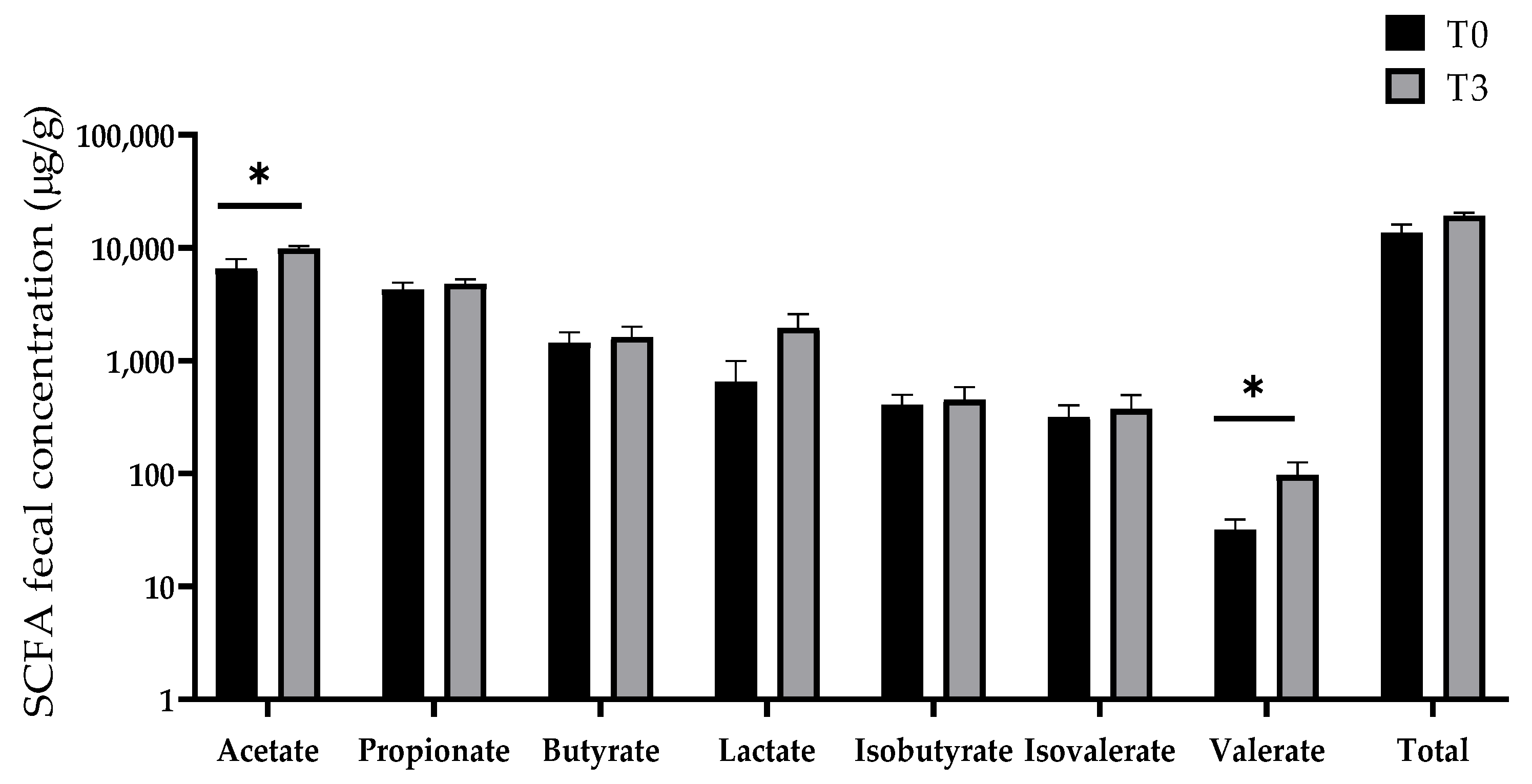
3.7. SCFA Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

References
- Xu, Q.; Jian, H.; Zhao, W.; Li, J.; Zou, X.; Dong, X. Early Weaning Stress Induces Intestinal Microbiota Disturbance, Mucosal Barrier Dysfunction and Inflammation Response Activation in Pigeon Squabs. Front. Microbiol. 2022, 13, 877866. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Morris, A.; Kayal, A.; Milošević, I.; Van, T.; Sharma Bajagai, Y.; Stanley, D. Pioneering Gut Health Improvements in Piglets with Phytogenic Feed Additives. Appl. Microbiol. Biotechnol. 2024, 108, 142. [Google Scholar] [CrossRef]
- Moeser, A.J.; Pohl, C.S.; Rajput, M. Weaning Stress and Gastrointestinal Barrier Development: Implications for Lifelong Gut Health in Pigs. Anim. Nutr. 2017, 3, 313–321. [Google Scholar] [CrossRef]
- Rhouma, M.; Fairbrother, J.M.; Beaudry, F.; Letellier, A. Post Weaning Diarrhea in Pigs: Risk Factors and Non-Colistin-Based Control Strategies. Acta Vet. Scand. 2017, 59, 31. [Google Scholar] [CrossRef] [PubMed]
- Vinelli, V.; Biscotti, P.; Martini, D.; Del Bo’, C.; Marino, M.; Meroño, T.; Nikoloudaki, O.; Calabrese, F.M.; Turroni, S.; Taverniti, V.; et al. Effects of Dietary Fibers on Short-Chain Fatty Acids and Gut Microbiota Composition in Healthy Adults: A Systematic Review. Nutrients 2022, 14, 2559. [Google Scholar] [CrossRef]
- Usuda, H.; Okamoto, T.; Wada, K. Leaky Gut: Effect of Dietary Fiber and Fats on Microbiome and Intestinal Barrier. Int. J. Mol. Sci. 2021, 22, 7613. [Google Scholar] [CrossRef]
- Yin, Y.; Wang, F.; Yang, M.; Yin, Y.; Chen, J.; Yang, Z. Lycium barbarum Polysaccharides as Antibiotic Substitutes Improve Growth Performance, Serum Immunity, Antioxidant Status, and Intestinal Health for Weaned Piglets. Front. Microbiol. 2022, 12, 819993. [Google Scholar] [CrossRef] [PubMed]
- Galgano, S.; Conway, L.; Fellows, A.; Houdijk, J. Impact of Precursor-Derived Peracetic Acid on Post-Weaning Diarrhea, Intestinal Microbiota, and Predicted Microbial Functional Genes in Weaned Pigs. Front. Microbiol. 2024, 15, 1356538. [Google Scholar] [CrossRef]
- Duarte, M.E.; Kim, S.W. Phytobiotics from Oregano Extracts Enhance the Intestinal Health and Growth Performance of Pigs. Antioxidants 2022, 11, 2066. [Google Scholar] [CrossRef]
- Zhu, Q.; Azad, M.d.A.K.; Li, R.; Li, C.; Liu, Y.; Yin, Y.; Kong, X. Dietary Probiotic and Synbiotic Supplementation Starting from Maternal Gestation Improves Muscular Lipid Metabolism in Offspring Piglets by Reshaping Colonic Microbiota and Metabolites. mSystems 2024, 9, e0004824. [Google Scholar] [CrossRef]
- Omolaso, B.O.; Oluwole, F.S.; Odukanmi, O.A.; Adesanwo, J.K.; Ishola, A.A.; Adewole, K.E. Evaluation of the Gastrointestinal Anti-Motility Effect of Anacardium occidentale Stem Bark Extract: A Mechanistic Study of Antidiarrheal Activity. J. Pharm. Anal. 2021, 11, 776–782. [Google Scholar] [CrossRef] [PubMed]
- Oko, A.O.; Okose, V.C.; Ekuma, E.T. Anacardium occidentale (Linn) Stem Bark Extracts: Effects on Poultry Colibacillosis Disease. Nig. J. Biotechnol. 2022, 39, 74–86. [Google Scholar] [CrossRef]
- Hashim, N.S.; Tan, M.L.; Ooi, K.L.; Sulaiman, S.F. The Effect of Flavonols in Anacardium occidentale L. Leaf Extracts on Skin Pathogenic Microorganisms. Nat. Prod. Res. 2023, 37, 2009–2012. [Google Scholar] [CrossRef] [PubMed]
- Martínez, Y.; Martínez, O.; Liu, G.; Ren, W.; Rodríguez, R.; Fonseca, Y.; Olmo, C.; Isert, M.; Aroche, R.; Valdivié, M.; et al. Effect of Dietary Supplementation with Anacardium occidentale on Growth Performance and Immune and Visceral Organ Weights in Replacement Laying Pullets. J. Food Agric. Environ. 2013, 11, 1352–1357. [Google Scholar]
- Aroche, R.; Martínez, Y.; Ayala, L.; Rodríguez, R.; Rodríguez, Y. Growth Performance and Diarrhea Incidence in Post-Weaning Pigs, Supplemented with Plant Mixed Leaves Powder, with Nutraceutical Properties. Rev. Cien. Agri. 2017, 14, 19–26. [Google Scholar] [CrossRef]
- Aroche, R.; Martínez, Y.; Ruan, Z.; Guan, G.; Waititu, S.; Nyachoti, C.M.; Más, D.; Lan, S. Dietary Inclusion of a Mixed Powder of Medicinal Plant Leaves Enhances the Feed Efficiency and Immune Function in Broiler Chickens. J. Chem. 2018, 2018, 4073068. [Google Scholar] [CrossRef]
- Más, D.; Martínez, Y.; Rodríguez, R.; Salazar, I.; Aroche, R.; López, B.; Marcella, D. Efecto de la Suplementación Dietética Con Polvos de Hojas de Guayaba (Psidium guajava) y Marañón (Anacardium occidentale) En El Comportamiento Productivo y la Incidencia de Diarrea En Cerdos Antes y Después del Destete. Rev. Comput. Prod. Porc. 2016, 23, 106–113. [Google Scholar]
- National Research Council. Nutrient Requirements of Swine, 11th ed.; National Academic Press: Washington, DC, USA, 2012. [Google Scholar]
- Jiang, X.R.; Awati, A.; Agazzi, A.; Vitari, F.; Ferrari, A.; Bento, H.; Crestani, M.; Domeneghini, C.; Bontempo, V. Effects of a Blend of Essential Oils and an Enzyme Combination on Nutrient Digestibility, Ileum Histology and Expression of Inflammatory Mediators in Weaned Piglets. Animal 2015, 9, 417–426. [Google Scholar] [CrossRef]
- Jiang, X.R.; Agazzi, A.; Awati, A.; Vitari, F.; Bento, H.; Ferrari, A.; Alborali, G.L.; Crestani, M.; Domeneghini, C.; Bontempo, V. Influence of a Blend of Essential Oils and an Enzyme Combination on Growth Performance, Microbial Counts, Ileum Microscopic Anatomy and the Expression of Inflammatory Mediators in Weaned Piglets Following an Escherichia coli Infection. Anim. Feed Sci. Technol. 2015, 209, 219–229. [Google Scholar] [CrossRef]
- Aroche, R.; Xian-Ren, J.; Bertot, R.; Li, X.; Avellaneda Barbosa, M.; Martínez, Y. In Vitro Antimicrobial and Antioxidant Activity of Leaves and Aqueous Extract of Four Medicinal Plants with Phytobiotic Potential in Animal Production. Cuban J. Agric. Sci. 2023, 57, e27. [Google Scholar]
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The Biological Stress of Early Weaned Piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Sunderam, V.; Thiyagarajan, D.; Lawrence, A.V.; Mohammed, S.S.S.; Selvaraj, A. In-Vitro Antimicrobial and Anticancer Properties of Green Synthesized Gold Nanoparticles Using Anacardium occidentale Leaves Extract. Saudi J. Biol. Sci. 2019, 26, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Duangjan, C.; Rangsinth, P.; Gu, X.; Wink, M.; Tencomnao, T. Lifespan Extending and Oxidative Stress Resistance Properties of a Leaf Extracts from Anacardium occidentale L. in Caenorhabditis elegans. Oxid. Med. Cell. Longev. 2019, 2019, e9012396. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, R.; Fusco, R.; Peritore, A.F.; Cordaro, M.; D’Amico, R.; Genovese, T.; Gugliandolo, E.; Crupi, R.; Smeriglio, A.; Mandalari, G.; et al. The Antioxidant and Anti-Inflammatory Properties of Anacardium occidentale L. Cashew Nuts in a Mouse Model of Colitis. Nutrients 2020, 12, 834. [Google Scholar] [CrossRef]
- Chotphruethipong, L.; Benjakul, S.; Kijroongrojana, K. Optimization of Extraction of Antioxidative Phenolic Compounds from Cashew (Anacardium occidentale L.) Leaves Using Response Surface Methodology. J. Food Biochem. 2017, 41, e12379. [Google Scholar] [CrossRef]
- Ukwenya, V.O.; Alese, M.O.; Ogunlade, B.; Folorunso, I.M.; Omotuyi, O.I. Anacardium occidentale Leaves Extract and Riboceine Mitigate Hyperglycemia through Anti-Oxidative Effects and Modulation of Some Selected Genes Associated with Diabetes. J. Diabetes Metab. Disord. 2023, 22, 455–468. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Peng, Q.; Wu, Y.; Peng, C.; Wang, S.; Zou, L.; Qi, M.; Peng, C.; Liu, H.; Li, R.; et al. The Effect of an Essential Oil Blend on Growth Performance, Intestinal Health, and Microbiota in Early-Weaned Piglets. Nutrients 2023, 15, 450. [Google Scholar] [CrossRef]
- Ma, L.; Zhou, B.; Liu, H.; Chen, S.; Zhang, J.; Wang, T.; Wang, C. Dietary Rutin Improves the Antidiarrheal Capacity of Weaned Piglets by Improving Intestinal Barrier Function, Antioxidant Capacity, and Cecal Microbiota Composition. J. Sci. Food Agric. 2024, 104, 6262–6275. [Google Scholar] [CrossRef]
- May, S.; Knoell, A.; Vega, C.G.; Sardi, M.I.; Khafipour, E. 192 Effects of Inclusion of a Probiotic and/or a Postbiotic in Diets Containing a Phytogenic Feed Additive on Growth Performance and Fecal Microbiome Composition and Function of Nursery Pigs. J. Anim. Sci. 2024, 102, 136–137. [Google Scholar] [CrossRef]
- Liu, S.; Yunxia, X.; Cao, S.; Wen, X.; Xiao, H.; Li, Y.; Chi, L.; He, D.; Jiang, Z.; Wang, L. Dietary Stevia Residue Extract Supplementation Improves Antioxidant Capacity and Intestinal Microbial Composition of Weaned Piglets. Antioxidants 2022, 11, 2016. [Google Scholar] [CrossRef]
- Deng, Z.; Wang, J.; Wang, J.; Yan, Y.; Huang, Y.; Chen, C.; Sun, L.; Liu, M. Tannic Acid Extracted from Gallnut Improves Intestinal Health with Regulation of Redox Homeostasis and Gut Microbiota of Weaned Piglets. Anim. Res. One Health 2024, 2, 16–27. [Google Scholar] [CrossRef]
- Zhu, J.; Lian, J.; Deng, H.; Luo, J.; Chen, T.; Sun, J.; Zhang, Y.-L.; Yang, Y.; Liu, P.; Xi, Q. Effects of Spinach Extract and Licorice Extract on Growth Performance, Antioxidant Capacity, and Gut Microbiota in Weaned Piglets. Animals 2024, 14, 321. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Chen, Z.; Huang, Z.; Wan, A.; Zhou, M.; Gao, J. Effects of Dietary Traditional Chinese Medicine Residues on Growth Performance, Intestinal Health and Gut Microbiota Compositions in Weaned Piglets. Front. Cell. Infect. Microbiol. 2023, 13, 1283789. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Wu, H.; Wang, X.; He, J.; He, S.; Yin, Y. Resveratrol Attenuates Oxidative Stress-Induced Intestinal Barrier Injury through PI3K/Akt-Mediated Nrf2 Signaling Pathway. Oxid. Med. Cell. Longev. 2019, 2019, 7591840. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Ma, J.; Kang, M.; Tang, W.; Xia, S.; Yin, J.; Yin, Y. Flavonoids, Gut Microbiota, and Host Lipid Metabolism. Eng. Life Sci. 2023, 24, e2300065. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Q.; Zhuo, Y.; Fang, Z.; Che, L.; Xu, S.; Feng, B.; Lin, Y.; Jiang, X.; Zhao, X.; et al. Effects of Multi-Strain Probiotics and Perilla frutescens Seed Extract Supplementation Alone or Combined on Growth Performance, Antioxidant Indices, and Intestinal Health of Weaned Piglets. Animals 2022, 12, 2246. [Google Scholar] [CrossRef]
- Xiao, Y.; Huang, R.; Wang, N.; Yuankun, D.; Yin, Y.; Qi, M.; Jing, W. Ellagic Acid Alleviates Oxidative Stress by Mediating Nrf2 Signaling Pathways and Protects against Paraquat-Induced Intestinal Injury in Piglets. Antioxidants 2022, 11, 252. [Google Scholar] [CrossRef]
- Pierzynowska, K.; Woliński, J.; Weström, B.; Pierzynowski, S.G. Maternal Immunoglobulins in Infants—Are They More than Just a Form of Passive Immunity? Front. Immunol. 2020, 11, 855. [Google Scholar] [CrossRef]
- Xiao, C.; Kai, L.; Teng, C.; Wei, Z.; Li, J.; Zhang, S.; Liu, L.; Lv, H.; Zhong, R. Dietary Qi-Weng-Huangbo Powder Enhances Growth Performance, Diarrhoea and Immune Function of Weaned Piglets by Modulating Gut Health and Microbial Profiles. Front. Immunol. 2023, 14, 1342852. [Google Scholar] [CrossRef]
- Abbas, A.K.; Lichtman, A.H.; Pillai, S. Cellular and Molecular Immunology, 10e, South Asia Edition—E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2021; ISBN 978-81-312-6458-4. [Google Scholar]
- Zeng, Y.; Dai, X.; Chen, Q.; Liu, Y.; Bumbie, G.Z.; Sun, W.; Tang, Z. Effect of Dietary Pomelo Peel Powder on Growth Performance, Diarrhea, Immune Function, Antioxidant Function, Ileum Morphology, and Colonic Microflora of Weaned Piglets. Animals 2022, 12, 3216. [Google Scholar] [CrossRef]
- Fu, L.; Sun, M.; Dong, W.; Zhang, G.; Han, D.; Zang, J.; Liu, H. Effects of Compound of Hawthorn (Crataegus pinnatifida) and Chinese Yam (Dioscorea opposita Thunb.) Extracts on Growth Performance, Intestinal Health, and Immune Function in Weaned Pigs. Anim. Sci. J. 2022, 93, e13790. [Google Scholar] [CrossRef]
- Mohammadi Gheisar, M.; Kim, I.H. Phytobiotics in Poultry and Swine Nutrition—A Review. Ital. J. Anim. Sci. 2018, 17, 92–99. [Google Scholar] [CrossRef]
- Johansson, M.E.V.; Hansson, G.C. Immunological Aspects of Intestinal Mucus and Mucins. Nat. Rev. Immunol. 2016, 16, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Trachsel, J.; Briggs, C.; Gabler, N.K.; Allen, H.K.; Loving, C.L. Dietary Resistant Potato Starch Alters Intestinal Microbial Communities and Their Metabolites, and Markers of Immune Regulation and Barrier Function in Swine. Front. Immunol. 2019, 10, 1381. [Google Scholar] [CrossRef]
- Suzuki, T. Regulation of the Intestinal Barrier by Nutrients: The Role of Tight Junctions. Anim. Sci. J. 2020, 91, e13357. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Rubin, B.K.; Voynow, J.A. Mucins, Mucus, and Goblet Cells. CHEST 2018, 154, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhang, X.; Liu, X.; Li, Y.; Han, D.; Pi, Y.; Whitmore, M.; Xingmiao, L.; Zhang, G.; Zheng, J.; et al. Strain Specificity of Lactobacilli with Promoted Colonization by Galactooligosaccharides Administration in Protecting Intestinal Barriers during Salmonella Infection. J. Adv. Res. 2023, 56, 1–14. [Google Scholar] [CrossRef]
- Sartor, R.B.; Mazmanian, S.K. Intestinal Microbes in Inflammatory Bowel Diseases. Am. J. Gastroenterol. Suppl. 2012, 1, 15–21. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, Stability and Resilience of the Human Gut Microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef]
- Heo, J.M.; Opapeju, F.O.; Pluske, J.R.; Kim, J.C.; Hampson, D.J.; Nyachoti, C.M. Gastrointestinal Health and Function in Weaned Pigs: A Review of Feeding Strategies to Control Post-Weaning Diarrhoea without Using in-Feed Antimicrobial Compounds. J. Anim. Physiol. Anim. Nutr. 2013, 97, 207–237. [Google Scholar] [CrossRef]
- Priyadarshini, M.; Kotlo, K.U.; Dudeja, P.K.; Layden, B.T. Role of Short Chain Fatty Acid Receptors in Intestinal Physiology and Pathophysiology. Compr. Physiol. 2018, 8, 1091–1115. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | Pre-Starter (d 0–14) | Starter (d 14–28) |
|---|---|---|
| Corn | 48.04 | 61.32 |
| Soybean meal | 13.50 | 18.00 |
| Extruded soybean | 12.00 | 5.00 |
| Fish meal | 5.60 | 3.00 |
| Whey | 15.00 | 5.00 |
| Soybean oil | 1.30 | 1.30 |
| Monocalcium phosphate | 0.35 | 0.60 |
| Limestone (CaCO3) | 0.77 | 0.95 |
| Salt | 0.40 | 0.40 |
| L-lysine HCL | 1.10 | 1.03 |
| DL-methionine | 0.08 | 0.07 |
| Threonine | 0.26 | 0.24 |
| Tryptophane | 0.02 | 0.01 |
| Wheat bran | 0.11 | 1.81 |
| Choline chloride (60%) | 0.05 | 0.05 |
| Phytase | 0.02 | 0.02 |
| Acidifier | 0.20 | 0.20 |
| Premix 1 | 1.00 | 1.00 |
| Zinc oxide | 0.20 | - |
| Total | 100.00 | 100.00 |
| Analyzed nutrient levels, % | ||
| Crude protein | 20.26 | 18.53 |
| Calcium | 1.12 | 0.82 |
| Phosphorus | 0.75 | 0.58 |
| Calculated nutrient content | ||
| ME 2, kcal/kg | 3350 | 3310 |
| SID lysine 3, % | 1.30 | 1.15 |
| SID methionine, % | 0.38 | 0.34 |
| SID threonine, % | 0.76 | 0.68 |
| SID tryptophan, % | 0.21 | 0.19 |
| SID valine, % | 0.82 | 0.73 |
| SID isoleucine, % | 0.69 | 0.59 |
| Gene | Accession Number | Primer Sequence (5′-3′) | Size (bp) |
|---|---|---|---|
| GAPDH | NM_001206359.1 | F: GCTTGTCATCAATGGAAAGG | 86 |
| R: CATACGTAGCACCAGCATCA | |||
| Occludin | NM_001163647.2 | F: TCAGGTGCACCCTCCAGATT | 112 |
| R: TGGACTTTCAAGAGGCCTGG | |||
| Claudin-1 | NM_001244539.1 | F: CCTCAATACAGGAGGGAAGC | 76 |
| R: CTCTCCCCACATTCGAGATGATT | |||
| ZO-1 | CV870309 | F: CGATCACTCCAGCATACAAT | 111 |
| R: CACTTGGCAGAAGATTGTGA |
| Experimental Treatments | ||||||||
|---|---|---|---|---|---|---|---|---|
| Items | T0 | T1 | T2 | T3 | SEM | p-Value | ||
| ANOVA | Linear | Quadratic | ||||||
| Body Weight (kg) | ||||||||
| Day 0 | 8.03 | 8.03 | 8.03 | 8.03 | 0.31 | 1.000 | 0.953 | 0.988 |
| Day 14 | 10.72 | 10.68 | 10.50 | 10.59 | 0.32 | 0.952 | 0.666 | 0.835 |
| Day 28 | 14.19 | 14.32 | 14.16 | 15.46 | 0.52 | 0.458 | 0.230 | 0.384 |
| Average Daily Gain (g) | ||||||||
| Days 0–14 | 192 | 189 | 177 | 183 | 9.59 | 0.951 | 0.655 | 0.838 |
| Days 14–28 | 248 | 260 | 261 | 348 | 24.54 | 0.478 | 0.195 | 0.461 |
| Days 0–28 | 220 | 225 | 219 | 265 | 11.41 | 0.441 | 0.222 | 0.372 |
| Average Daily Feed Intake (g) | ||||||||
| Days 0–14 | 267 | 299 | 261 | 251 | 10.38 | 0.484 | 0.417 | 0.354 |
| Days 14–28 | 449 | 495 | 474 | 598 | 28.39 | 0.147 | 0.054 | 0.404 |
| Days 0–28 | 358 | 397 | 368 | 425 | 13.99 | 0.233 | 0.132 | 0.720 |
| Gain–Feed Intake Ratio | ||||||||
| Days 0–14 | 0.72 | 0.63 | 0.65 | 0.73 | 0.02 | 0.096 | 0.722 | 0.017 |
| Days 14–28 | 0.46 | 0.51 | 0.55 | 0.58 | 0.03 | 0.717 | 0.266 | 0.869 |
| Days 0–28 | 0.60 | 0.57 | 0.60 | 0.63 | 0.02 | 0.793 | 0.511 | 0.497 |
| Experimental Treatments | |||||
|---|---|---|---|---|---|
| Items | T0 | T1 | T2 | T3 | p-Value |
| Days 0–14 | 6.35 | 5.16 | 5.15 | 9.92 | 0.106 |
| Days 14–28 | 18.47 a | 8.33 b | 9.35 b | 5.60 b | <0.001 |
| Days 0–28 | 12.63 a | 7.00 b | 7.50 b | 8.16 b | 0.007 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aroche, R.; Gao, G.; Li, Y.; Zhang, Y.; Rodríguez, R.; Martínez, Y.; Li, X. Effect of Anacardium occidentale Leaf Powder on Growth Performance, Diarrhea Incidence, Blood Biochemistry, and Intestinal Traits in Weaned Piglets. Animals 2024, 14, 3382. https://doi.org/10.3390/ani14233382
Aroche R, Gao G, Li Y, Zhang Y, Rodríguez R, Martínez Y, Li X. Effect of Anacardium occidentale Leaf Powder on Growth Performance, Diarrhea Incidence, Blood Biochemistry, and Intestinal Traits in Weaned Piglets. Animals. 2024; 14(23):3382. https://doi.org/10.3390/ani14233382
Chicago/Turabian StyleAroche, Roisbel, Ge Gao, Yanpin Li, Yonggang Zhang, Román Rodríguez, Yordan Martínez, and Xilong Li. 2024. "Effect of Anacardium occidentale Leaf Powder on Growth Performance, Diarrhea Incidence, Blood Biochemistry, and Intestinal Traits in Weaned Piglets" Animals 14, no. 23: 3382. https://doi.org/10.3390/ani14233382
APA StyleAroche, R., Gao, G., Li, Y., Zhang, Y., Rodríguez, R., Martínez, Y., & Li, X. (2024). Effect of Anacardium occidentale Leaf Powder on Growth Performance, Diarrhea Incidence, Blood Biochemistry, and Intestinal Traits in Weaned Piglets. Animals, 14(23), 3382. https://doi.org/10.3390/ani14233382