
Microbial Contamination Survey of Environmental Fresh and Saltwater Resources of Upolu Island, Samoa

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Sampling
2.2. Coliform Bacteria Detection
2.3. Sample Filtration for DNA Sequencing
2.4. Water Parameter Measurements
2.5. DNA Extraction, 16S rRNA Sequencing and Data Analysis
3. Results
3.1. Freshwater and Seawater Physicochemical Parameters
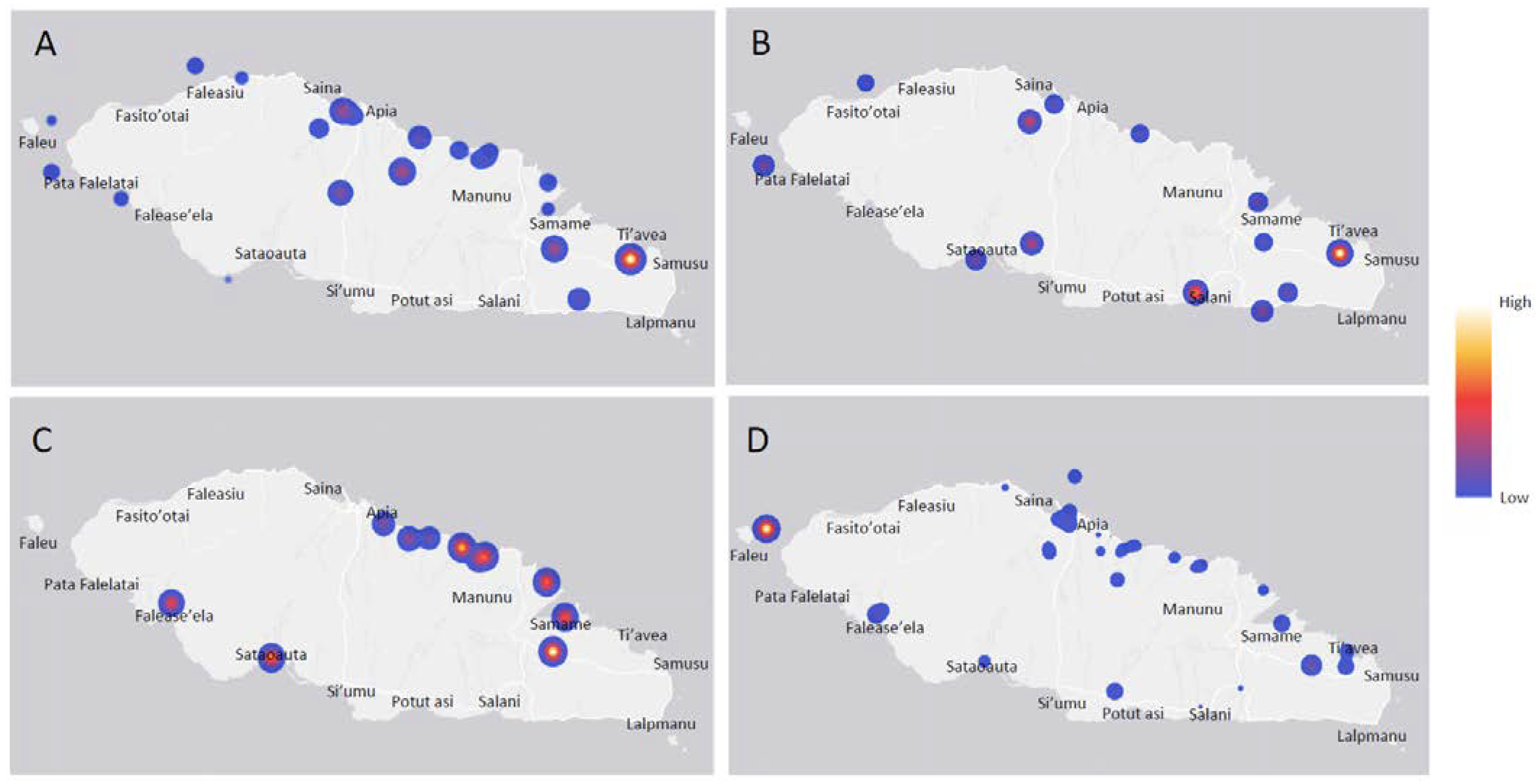
3.2. Contaminant Estimate with Fecal Indicator Bacteria (FIB)
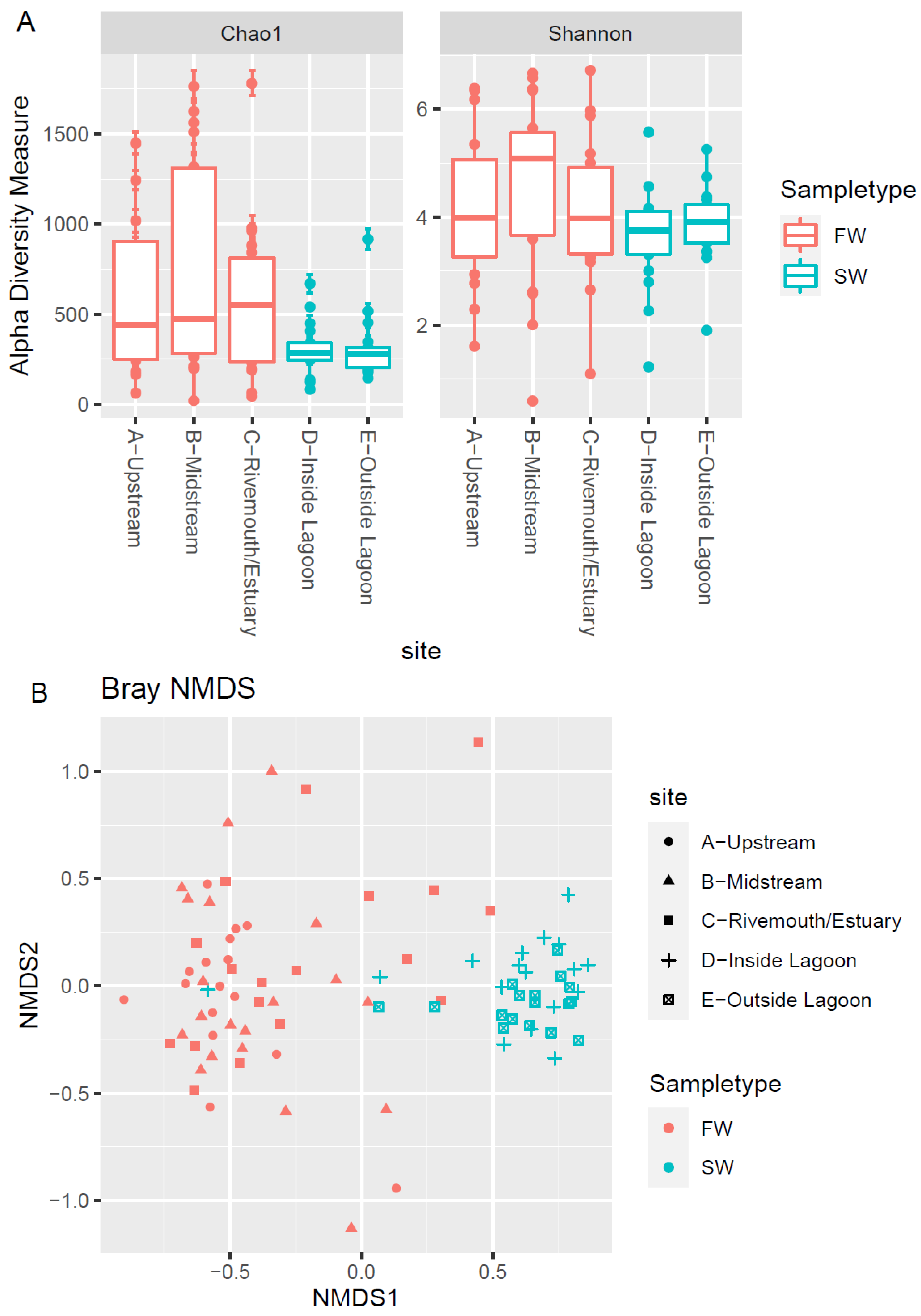
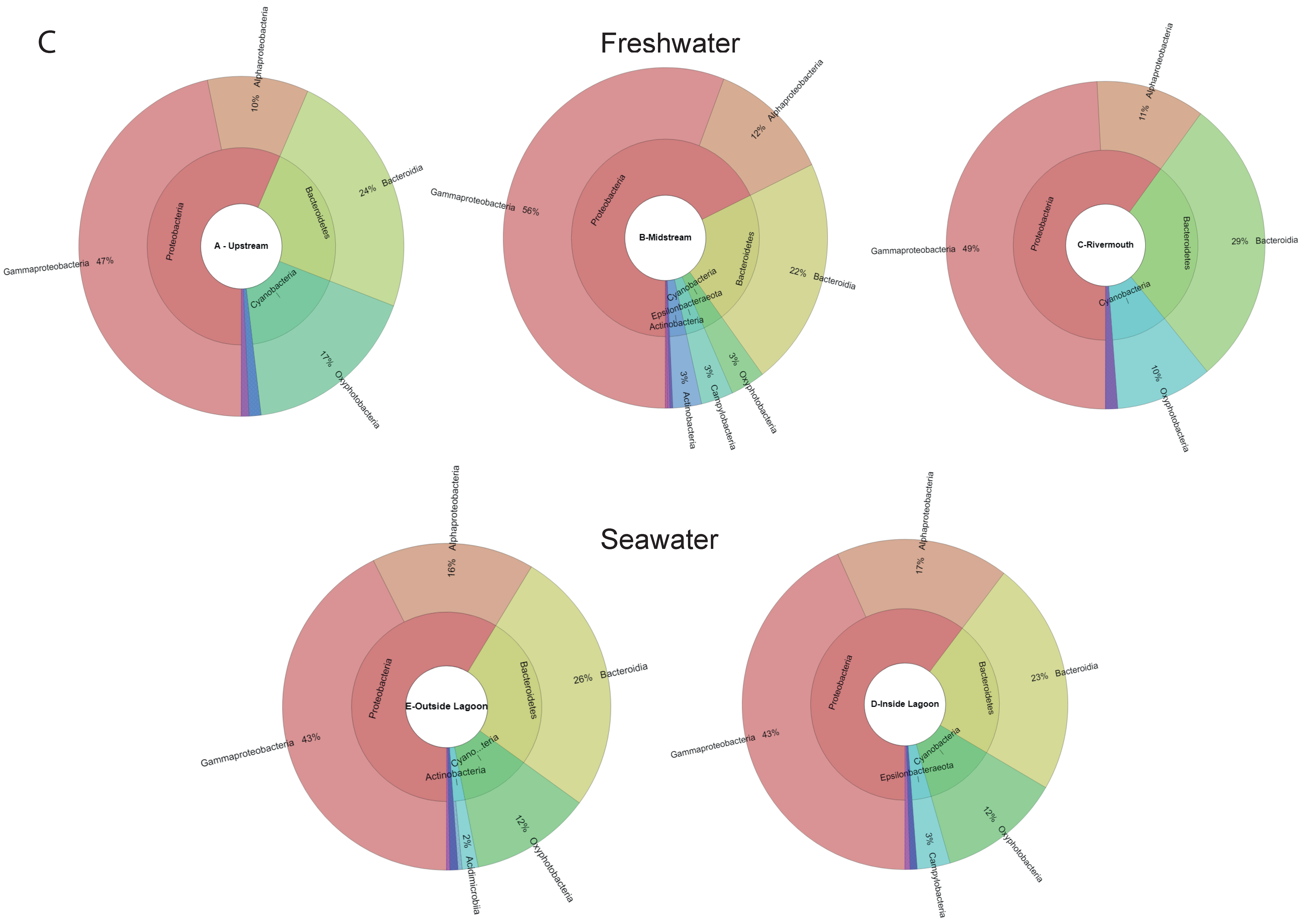
3.3. Microbial Community Biodiversity
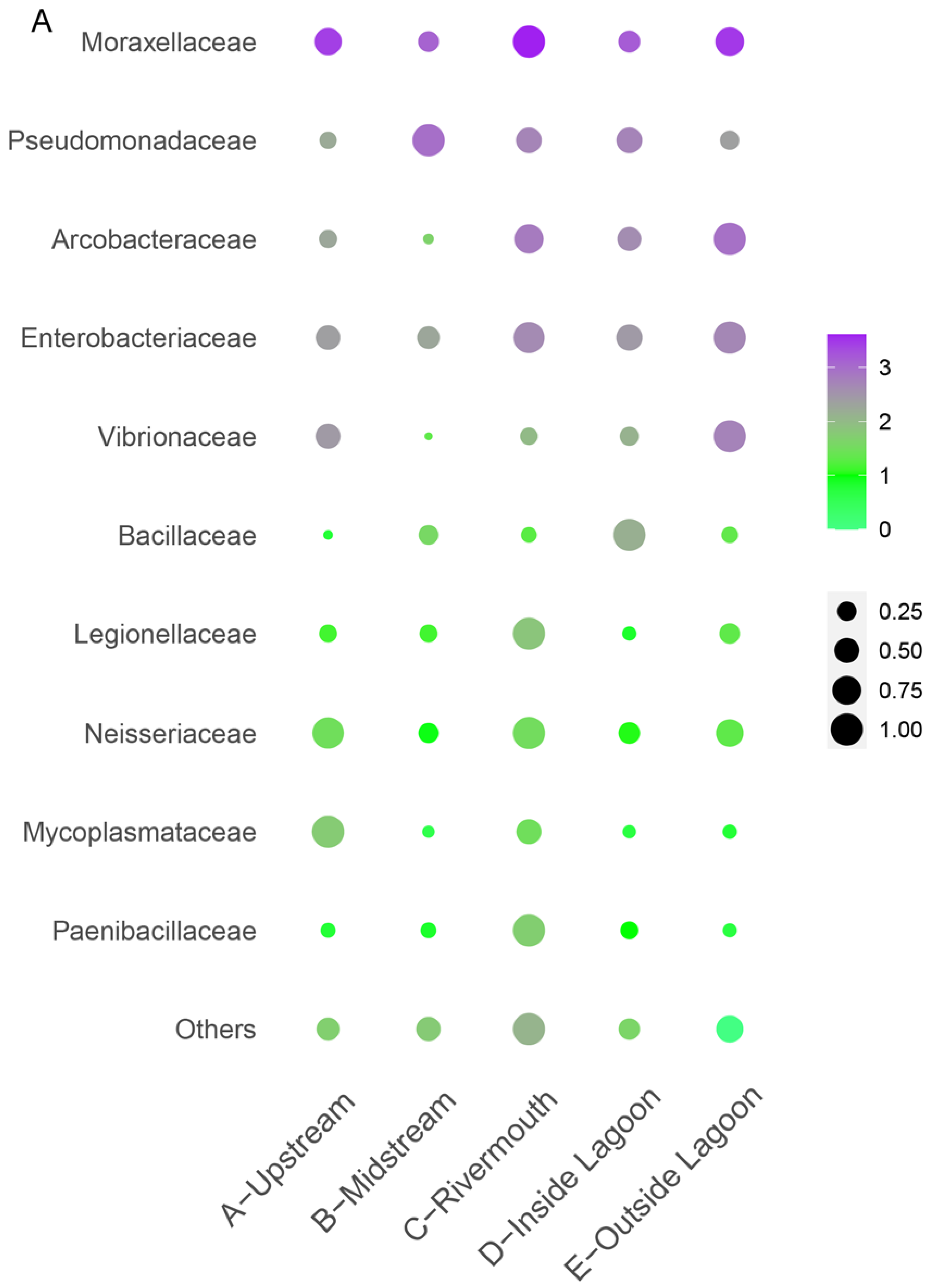
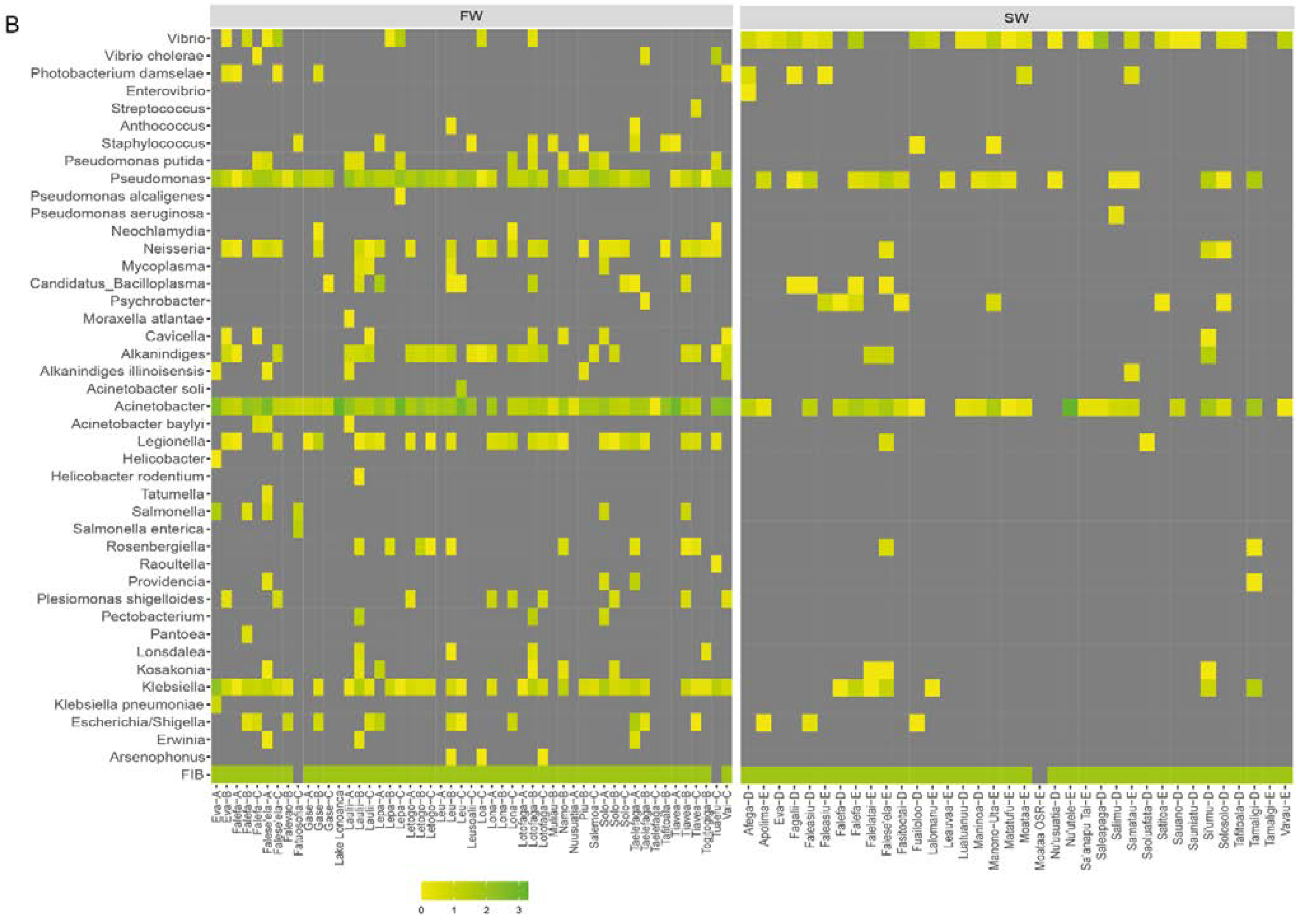
3.4. Distribution of Potential Pathogenic Bacteria
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nada Osseiran, W.; Yemi Lufadeju, U. 1 in 3 People Globally Do not Have Access to Safe Drinking Water—UNICEF. WHO. 2019. Available online: https://www.who.int/news/item/18-06-2019-1-in-3-people-globally-do-not-have-access-to-safe-drinking-water-unicef-who (accessed on 10 May 2021).
- Wear, S.L.; Thurber, R.V. Sewage pollution: Mitigation is key for coral reef stewardship. Ann. N. Y. Acad. Sci. 2015, 1355, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.A.; Post, G.B.; Buckley, B.T.; Lippincott, R.L.; Robson, M.G. Future challenges to protecting public health from drinking-water contaminants. Annu. Rev. Public Health 2012, 33, 209–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hancock, G.J.; Webster, I.T.; Stieglitz, T.C. Horizontal mixing of Great Barrier Reef waters: Offshore diffusivity determined from radium isotope distribution. J. Geophys. Res. Ocean. 2006, 111, 1–14. [Google Scholar] [CrossRef]
- Leonard, A.F.C.; Singer, A.; Ukoumunne, O.C.; Gaze, W.H.; Garside, R. Is it safe to go back into the water? A systematic review and meta-analysis of the risk of acquiring infections from recreational exposure to seawater. Int. J. Epidemiol. 2018, 47, 572–586. [Google Scholar] [CrossRef]
- Rabieh, S.; Bayaraa, O.; Romeo, E.; Amosa, P.; Calnek, K.; Idaghdour, Y.; Ochsenkühn, M.A.; Amin, S.A.; Goldstein, G.; Bromage, T.G. MH-ICP-MS Analysis of the Freshwater and Saltwater Environmental Resources of Upolu Island, Samoa. Molecules 2020, 25, 4871. [Google Scholar] [CrossRef]
- Liu, J.; Chen, X.; Shu, H.; Lin, X.; Zhou, Q.; Bramryd, T.; Shu, W.; Huang, L. Microbial community structure and function in sediments from e-waste contaminated rivers at Guiyu area of China. Environ. Pollut. 2018, 235, 171–179. [Google Scholar] [CrossRef]
- Varol, M.; Şen, B. Assessment of nutrient and heavy metal contamination in surface water and sediments of the upper Tigris River, Turkey. Catena 2012, 92, 1–10. [Google Scholar] [CrossRef]
- Whitehead, P.G.; Wilby, R.L.; Battarbee, R.W.; Kernan, M.; Wade, A.J. A review of the potential impacts of climate change on surface water quality. Hydrol. Sci. J. 2009, 54, 101–123. [Google Scholar] [CrossRef]
- Leong-nowell, T.A.; Leavai, F.; Ching, L.A.; Fiu, L.; Nisbet, M.; Jones, D.; Blackmore, T.; Ioane-cleverley, T. Aspiration pneumonia and challenges following the Samoa Tsunami in 2009. N. Z. Med. J. 2012, 125, 70–78. [Google Scholar]
- Sikorski, M.J.; Desai, S.N.; Tupua, S.; Thomsen, R.E.; Han, J.; Rambocus, S.; Nimarota-Brown, S.; Punimata, L.; Tusitala, S.; Sialeipata, M.; et al. Tenacious endemic typhoid fever in Samoa. Clin. Infect. Dis. 2020, 71, S120–S126. [Google Scholar] [CrossRef] [PubMed]
- Paul, J.H.; Rose, J.B.; Brown, J.; Shinn, E.A.; Miller, S.; Farrah, S.R. Viral tracer studies indicate contamination of marine waters by sewage disposal practices in Key Largo, Florida. Appl. Environ. Microbiol. 1995, 61, 2230–2234. [Google Scholar] [CrossRef] [Green Version]
- Richards, S.; Paterson, E.; Withers, P.J.A.; Stutter, M. Septic tank discharges as multi-pollutant hotspots in catchments. Sci. Total Environ. 2016, 542, 854–863. [Google Scholar] [CrossRef]
- Griffin, D.W.; Gibson, C.J.; Lipp, E.K.; Riley, K.; Paul, J.H.; Rose, J.B. Erratum: Detection of viral pathogens by reverse transcriptase PCR and of microbial indicators by standard methods in the canals of the Florida keys. Appl. Environ. Microbiol. 2000, 66, 876. [Google Scholar] [CrossRef] [Green Version]
- Fujita, M.; Suzuki, J.; Sato, D.; Kuwahara, Y.; Yokoki, H.; Kayanne, H. Anthropogenic impacts on water quality of the lagoonal coast of Fongafale Islet, Funafuti Atoll, Tuvalu. Sustain. Sci. 2013, 8, 381–390. [Google Scholar] [CrossRef] [Green Version]
- Amosa, P.; Imo, T.; Latu, F.; Vaurasi, V. Microbiological Assessment of River Water Quality in Samoa. Int. J. Sci. Res. 2016, 5, 225–234. [Google Scholar]
- Wade, T.J.; Pai, N.; Eisenberg, J.N.S.; Colford, J.M. Do U.S. Environmental Protection Agency water quality guidelines for recreational waters prevent gastrointestinal illness? A systematic review and meta-analysis. Environ. Health Perspect. 2003, 111, 1102–1109. [Google Scholar] [CrossRef] [PubMed]
- Harwood, V.J.; Levine, A.D.; Scott, T.M.; Chivukula, V.; Lukasik, J.; Farrah, S.R.; Rose, J.B. Validity of the indicator organism paradigm for pathogen reduction in reclaimed water and public health protection. Appl. Environ. Microbiol. 2005, 71, 3163–3170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, B.J.; Wong, J.; Heiner, C.; Oh, S.; Theriot, C.M.; Gulati, A.S.; McGill, S.K.; Dougherty, M.K. High-throughput amplicon sequencing of the full-length 16S rRNA gene with single-nucleotide resolution. bioRxiv 2018, 47, e103. [Google Scholar] [CrossRef] [Green Version]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [Green Version]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Dixon, P. Computer program review VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive metagenomic visualization in a Web browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef] [Green Version]
- WHO. Guidelines for Drinking-Water Quality; WHO: Geneva, Switzerland, 2011; Volume 1. [Google Scholar]
- Hazen, T.C. Fecal coliforms as indicators in tropical waters: A review. Toxic. Assess. 1988, 3, 461–477. [Google Scholar] [CrossRef]
- Anderson, D.A.; Armstrong, R.A.; Weil, E. Hyperspectral sensing of disease stress in the Caribbean reef-building coral, Orbicella faveolata—Perspectives for the field of coral disease monitoring. PLoS ONE 2013, 8, e81478. [Google Scholar] [CrossRef]
- Horne, A.J.; Goldman, C.R. Limnology; McGraw-Hill: New York, NY, USA, 1994. [Google Scholar]
- Lee, S.-Y.; Eom, Y.-B. Analysis of Microbial Composition Associated with Freshwater and Seawater. Biomed. Sci. Lett. 2016, 22, 150–159. [Google Scholar] [CrossRef] [Green Version]
- Hahn, M.W. The microbial diversity of inland waters. Curr. Opin. Biotechnol. 2006, 17, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Lemke, M.J.; Lienau, E.K.; Rothe, J.; Pagioro, T.A.; Rosenfeld, J.; Desalle, R. Description of freshwater bacterial assemblages from the Upper Paraná River floodpulse system, Brazil. Microb. Ecol. 2009, 57, 94–103. [Google Scholar]
- Humbert, J.F.; Dorigo, U.; Cecchi, P.; Le Berre, B.; Debroas, D.; Bouvy, M. Comparison of the structure and composition of bacterial communities from temperate and tropical freshwater ecosystems. Environ. Microbiol. 2009, 11, 2339–2350. [Google Scholar] [CrossRef] [PubMed]
- Hancock, J.T. Redox-Mediated Signal Transduction: Methods and Protocols; Humana Press: Totowa, NJ, USA, 2008; Volume 476, ISBN 9781588298423. [Google Scholar]
- Rochelle-Newall, E.; Nguyen, T.M.H.; Le, T.P.Q.; Sengtaheuanghoung, O.; Ribolzi, O. A short review of fecal indicator bacteria in tropical aquatic ecosystems: Knowledge gaps and future directions. Front. Microbiol. 2015, 6, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Winfield, M.D.; Groisman, E.A. Role of nonhost environments in the lifestyles of Salmonella and Escherichia coli. Appl. Environ. Microbiol. 2003, 69, 3687–3694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faiilagi, S.A. Assessing the impacts of land use patterns on river water quality at catchment level: A Case Study of Fuluasou River Catchment in Samoa. Master’s Thesis, Massey University, Palmerston North, New Zealand, 2015. [Google Scholar]
- Fabricius, K.E. Effects of terrestrial runoff on the ecology of corals and coral reefs: Review and synthesis. Mar. Pollut. Bull. 2005, 50, 125–146. [Google Scholar] [CrossRef] [PubMed]
- Keswani, A.; Oliver, D.M.; Gutierrez, T.; Quilliam, R.S. Microbial hitchhikers on marine plastic debris: Human exposure risks at bathing waters and beach environments. Mar. Environ. Res. 2016, 118, 10–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, T.P.; Anderson, K.D.; Connolly, S.R.; Heron, S.F.; Kerry, J.T.; Lough, J.M.; Baird, A.H.; Baum, J.K.; Berumen, M.L.; Bridge, T.C.; et al. Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science 2018, 359, 80–83. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.C.; Chan, W.W.; Metelitsa, A.I.; Fiorillo, L.; Lin, A.N. Pseudomonas skin infection: Clinical features, epidemiology, and management. Am. J. Clin. Dermatol. 2011, 12, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Chambers, H.F. The changing epidemiology of staphylococcus aureus? Emerg. Infect. Dis. 2001, 7, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Lowy, F.D. Staphylococcus Aureus Infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef]
- Taiaroa, G.; Matalavea, B.; Tafuna, M.; Lacey, J.A.; Price, D.J.; Isaia, L.; Leaupepe, H.; Viali, S.; Lee, D.; Gorrie, C.L.; et al. The Lancet Regional Health—Western Pacific Scabies and impetigo in Samoa: A school-based clinical and molecular epidemiological study. Lancet Reg. Health West. Pacific 2021, 6, 100081. [Google Scholar] [CrossRef]
- Motlagh, A.M.; Yang, Z. Detection and occurrence of indicator organisms and pathogens. Water Environ. Res. 2019, 91, 1402–1408. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.F.; Ng, C.; Nshimyimana, J.P.; Loh, L.L.; Gin, K.Y.H.; Thompson, J.R. Next-generation sequencing (NGS) for assessment of microbial water quality: Current progress, challenges, and future opportunities. Front. Microbiol. 2015, 6, 1027. [Google Scholar] [CrossRef] [PubMed]
- Rupic, M.; Wetzell, L.; Marra, J.J.; Salwani, S. 2014–2016 El Niño Assessment Report: An Overview of the Impacts of the 2014–16 El Niño on the U.S.-Affiliated Pacific Islands (USAPI); National Oceanic and Atmospheric Administration: Washington, DC, USA, 2018; p. 48. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Freshwater | DO # in mg/L | pH | ORP $ in mV | NO3+ in mg/L | Cl− in mg/L |
| Minimum | 2.1 ± 0.2 | 6.6 ± 0.1 | 76.5 ± 5 | 0.01 ± 0.05 | 2.18 |
| Median | 8.2 ± 0.2 | 7.5 ± 0.1 | 149.9 ± 5 | 0.01 ± 0.05 | 8 |
| Maximum | 10.8 ± 0.2 | 8.3 ± 0.1 | 191.1 ± 5 | 20.8 ± 0.05 | 8500 |
| Seawater | DO # in mg/L | pH | ORP $ | SPC & | PSU * |
| Minimum | 5.7 ± 0.2 | 7.8 ± 0.1 | 88.2 ± 5 | 51.1 ± 0.5 | 33.5 ± 0.5 |
| Median | 6.5 ± 0.2 | 7.9 ± 0.1 | 131.8 ± 5 | 53.3 ± 0.5 | 35.4 ± 0.5 |
| Maximum | 8.1 ± 0.2 | 8.0 ± 0.1 | 176.3 ± 5 | 59.7 ± 0.5 | 45.1 ± 0.5 |
| No. of Samples | Coliform | FIB > 1 Cell/100 mL | |
|---|---|---|---|
| Freshwater | 54 | 53 | 51 |
| Seawater | 34 | 33 | 33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ochsenkühn, M.A.; Fei, C.; Bayaara, O.; Romeo, E.; Amosa, P.; Idaghdour, Y.; Goldstein, G.; Bromage, T.G.; Amin, S.A. Microbial Contamination Survey of Environmental Fresh and Saltwater Resources of Upolu Island, Samoa. Environments 2021, 8, 112. https://doi.org/10.3390/environments8110112
Ochsenkühn MA, Fei C, Bayaara O, Romeo E, Amosa P, Idaghdour Y, Goldstein G, Bromage TG, Amin SA. Microbial Contamination Survey of Environmental Fresh and Saltwater Resources of Upolu Island, Samoa. Environments. 2021; 8(11):112. https://doi.org/10.3390/environments8110112
Chicago/Turabian StyleOchsenkühn, Michael A., Cong Fei, Odmaa Bayaara, Emarosa Romeo, Patila Amosa, Youssef Idaghdour, Gary Goldstein, Timothy G. Bromage, and Shady A. Amin. 2021. "Microbial Contamination Survey of Environmental Fresh and Saltwater Resources of Upolu Island, Samoa" Environments 8, no. 11: 112. https://doi.org/10.3390/environments8110112
APA StyleOchsenkühn, M. A., Fei, C., Bayaara, O., Romeo, E., Amosa, P., Idaghdour, Y., Goldstein, G., Bromage, T. G., & Amin, S. A. (2021). Microbial Contamination Survey of Environmental Fresh and Saltwater Resources of Upolu Island, Samoa. Environments, 8(11), 112. https://doi.org/10.3390/environments8110112